
The Vertical Metabolic Activity and Community Structure of Prokaryotes along Different Water Depths in the Kermadec and Diamantina Trenches

Abstract
1. Introduction
2. Materials and Methods
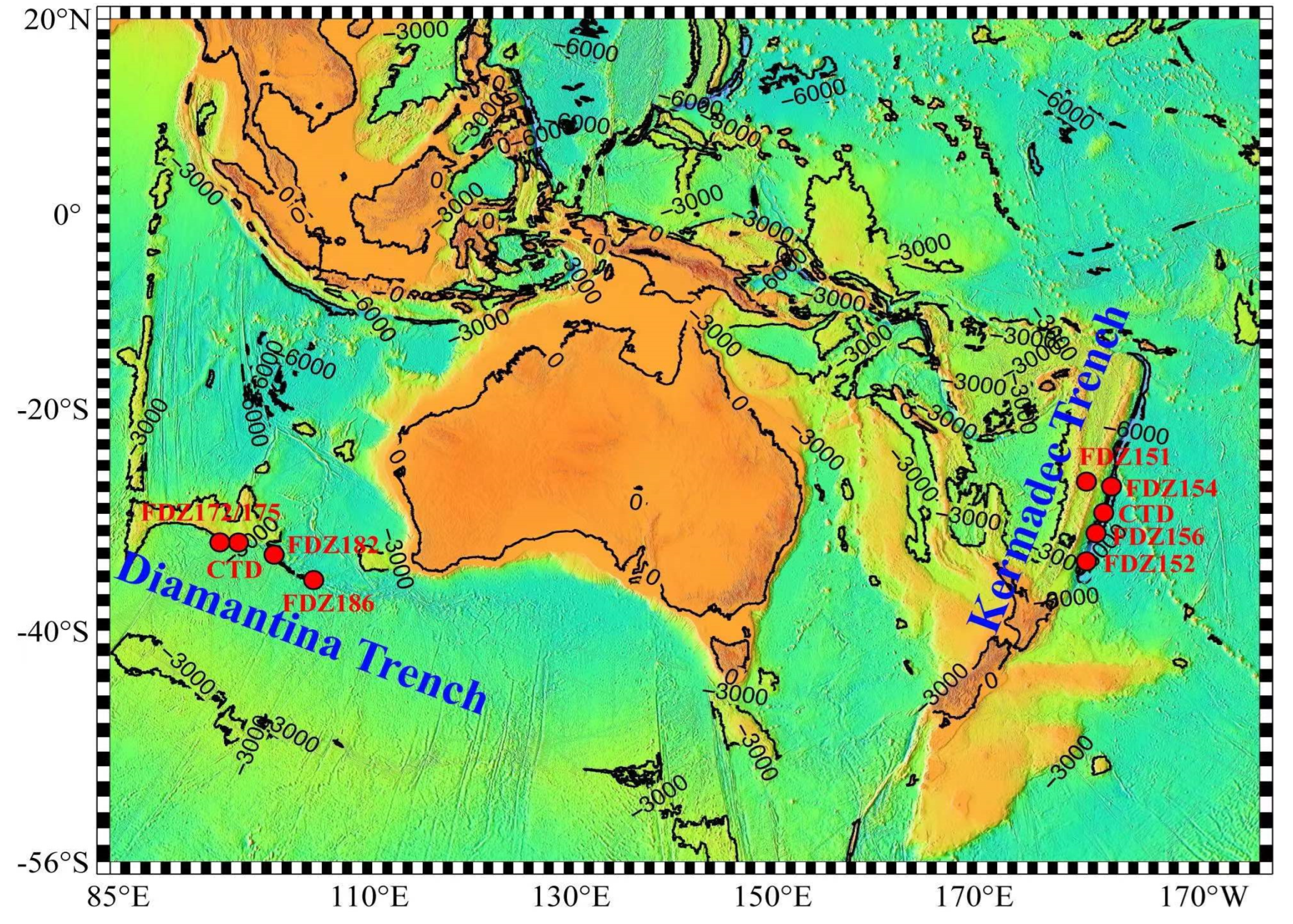
2.1. Sample Collection and Environmental Factor Measurement
2.2. Microbial Metabolic Activity Analyses
2.3. DNA Extraction, PCR Amplification, and Sequencing
2.4. Quantitative PCR
2.5. Bioinformatics Analysis
2.6. Statistical Analysis and Accession Number
3. Results
3.1. Hydrographic Conditions of Sampling Stations
3.2. Microbial Community Composition and Diversity
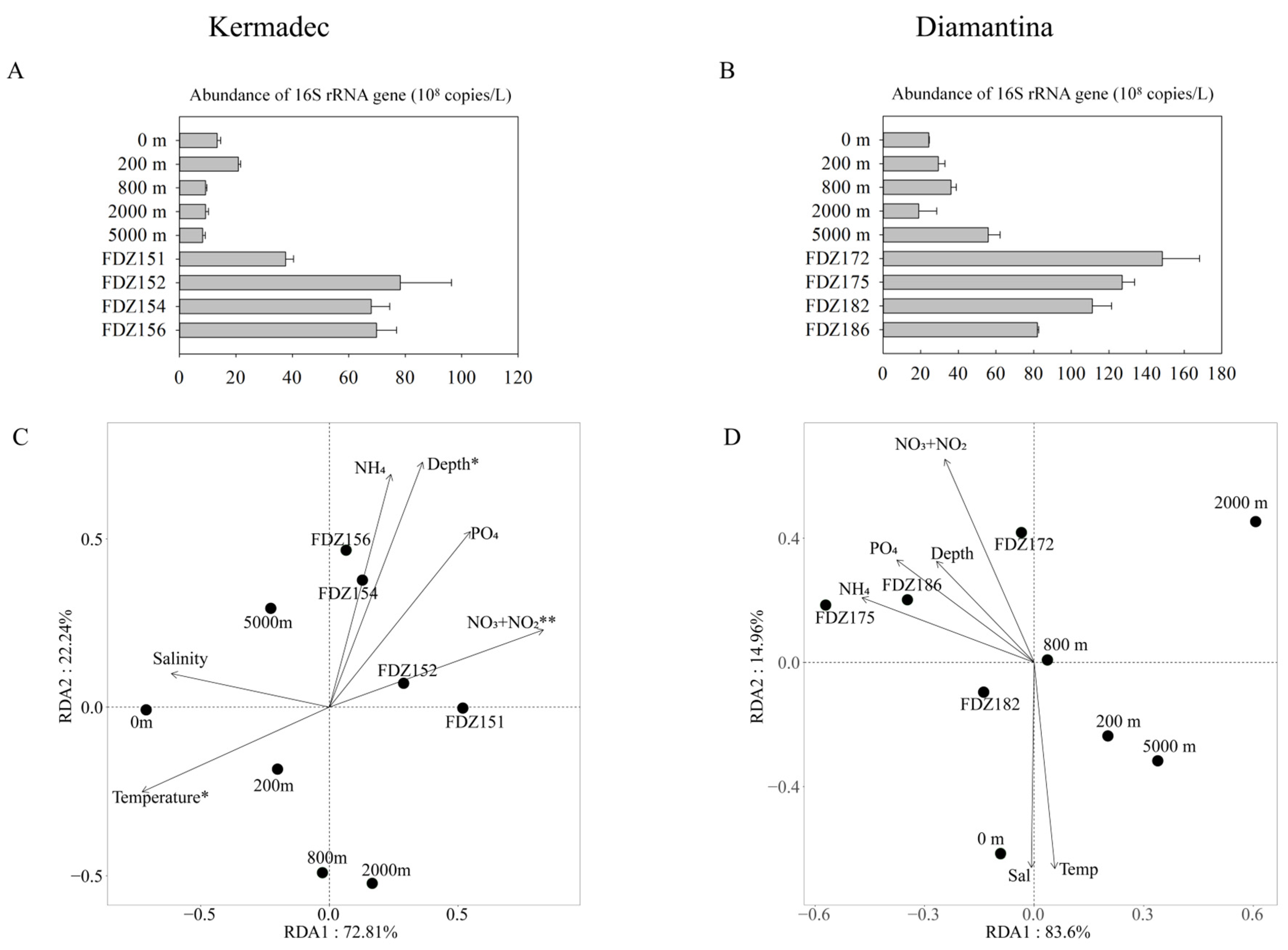
3.3. Gene Abundance and Environmental Impacts
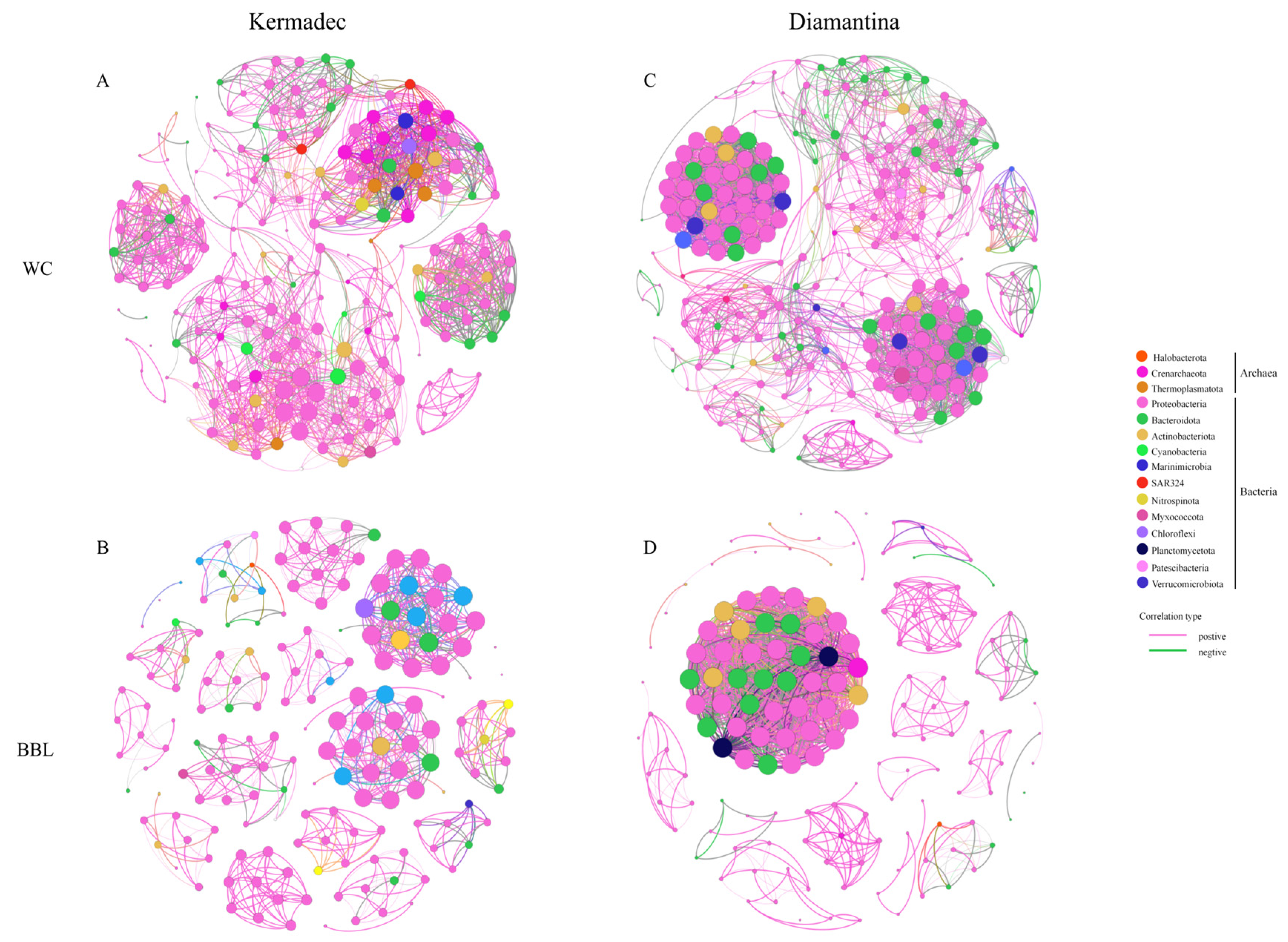
3.4. Co-Occurrence Network and Potential Metabolic Functions of Prokaryotes
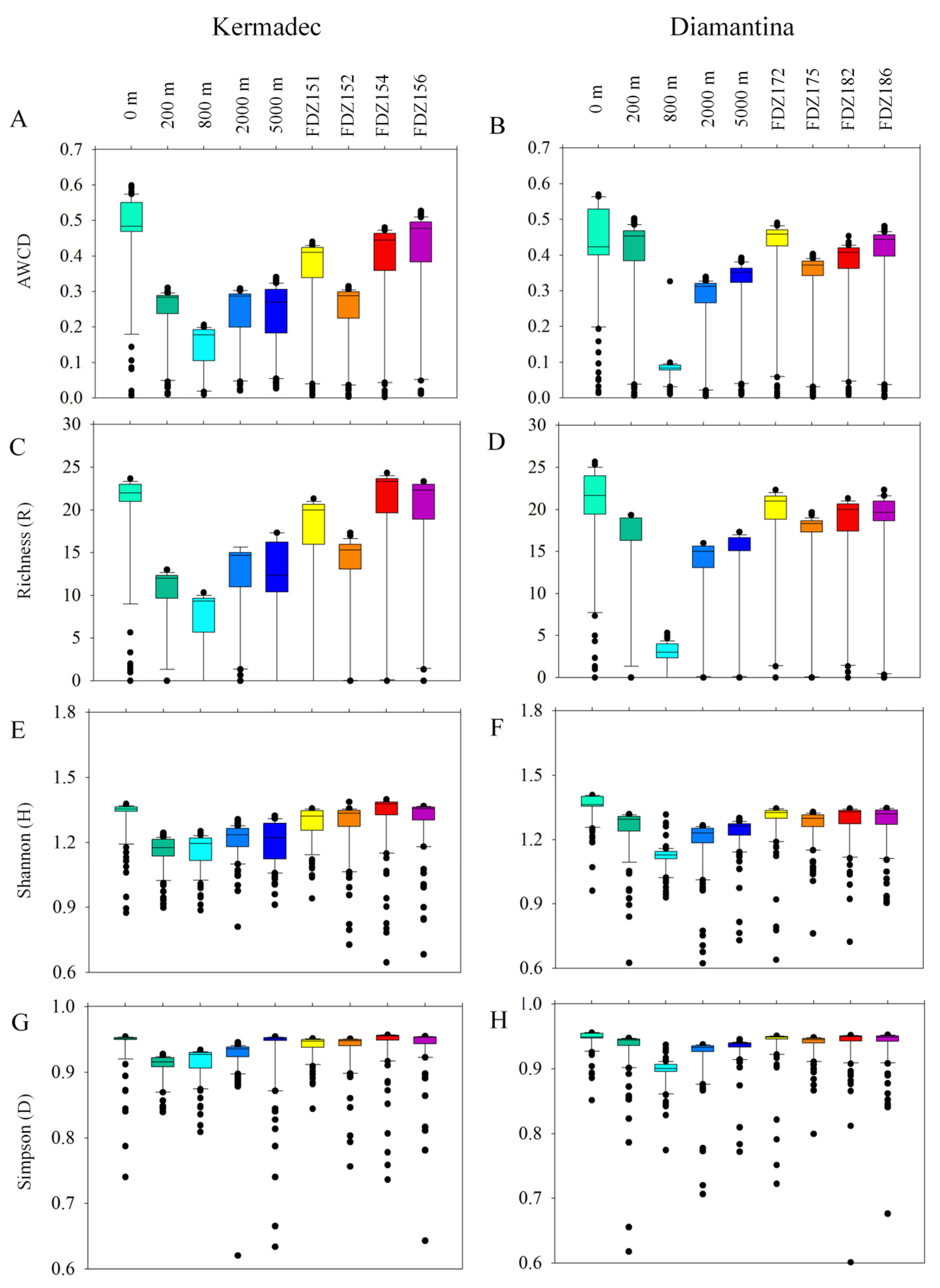
3.5. Prokaryotic Metabolic Activity
4. Discussion
4.1. Geographical Distribution and Environmental Effects
4.2. Carbon Source Utilization and Potential Metabolic Activity
4.3. Microbial Interaction and Ecological Significance
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Correction Statement
References
- Azam, F.; Malfatti, F. Microbial structuring of marine ecosystems. Nat. Rev. Microbiol. 2007, 5, 782–791. [Google Scholar] [CrossRef]
- Dang, H.; Lovell, C.R. Microbial surface colonization and biofilm development in marine environments. Microbiol. Mol. Biol. Rev. 2016, 80, 91–138. [Google Scholar] [CrossRef]
- Steinberg, D.K.; Van Mooy, B.A.S.; Buesseler, K.O.; Boyd, P.W.; Kobari, T.; Karl, D.M. Bacterial vs. zooplankton control of sinking particle flux in the ocean’s twilight zone. Limnol. Oceanogr. 2008, 53, 1327–1338. [Google Scholar] [CrossRef]
- Grabowski, E.; Letelier, R.M.; Laws, E.A.; Karl, D.M. Coupling carbon and energy fluxes in the North Pacific Subtropical Gyre. Nat. Commun. 2019, 10, 1895. [Google Scholar] [CrossRef] [PubMed]
- Herndl, G.J.; Reinthaler, T. Microbial control of the dark end of the biological pump. Nat. Geosci. 2013, 6, 718–724. [Google Scholar] [CrossRef]
- Mestre, M.; Ruiz-González, C.; Logares, R.; Duarte, C.M.; Gasol, J.M.; Sala, M.M. Sinking particles promote vertical connectivity in the ocean microbiome. Proc. Natl. Acad. Sci. USA 2018, 115, E6799–E6807. [Google Scholar] [CrossRef]
- Jamieson, A.J.; Fujii, T.; Mayor, D.J.; Solan, M.; Priede, I.G. Hadal trenches: The ecology of the deepest places on Earth. Trends Ecol. Evol. 2010, 25, 190–197. [Google Scholar] [CrossRef]
- Walsh, E.A.; Kirkpatrick, J.B.; Rutherford, S.D.; Smith, D.C.; Sogin, M.; D’hondt, S. Bacterial diversity and community composition from seasurface to subseafloor. ISME J. 2016, 10, 979–989. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Wei, X.; Song, W.; Wang, L.; Cao, J.; Wu, J.; Thomas, T.; Jin, T.; Wang, Z.; Wei, W.; et al. Novel Chloroflexi genomes from the deepest ocean reveal metabolic strategies for the adaptation to deep-sea habitats. Microbiome 2022, 10, 75. [Google Scholar] [CrossRef]
- Nagata, T.; Tamburini, C.; Arístegui, J.; Baltar, F.; Bochdansky, A.B.; Fonda-Umani, S.; Fukuda, H.; Gogou, A.; Hansell, D.A.; Hansman, R.L.; et al. Emerging concepts on microbial processes in the bathypelagic ocean—Ecology, biogeochemistry, and genomics. Deep. Sea Res. Part II 2010, 57, 1519–1536. [Google Scholar] [CrossRef]
- Hansell, D.A.; Ducklow, H.W. Bacterioplankton distribution and production in the bathypelagic ocean: Directly coupled to particulate organic carbon export? Limnol. Oceanogr. 2003, 48, 150–156. [Google Scholar] [CrossRef]
- Salazar, G.; Cornejo-Castillo, F.M.; Benítez-Barrios, V.; Fraile-Nuez, E.; Álvarez-Salgado, X.A.; Duarte, C.M.; Gasol, J.M.; Acinas, S.G. Global diversity and biogeography of deep-sea pelagic prokaryotes. ISME J. 2015, 10, 596–608. [Google Scholar] [CrossRef]
- Turley, C. Bacteria in the cold deep-sea benthic boundary layer and sediment-water interface of the NE Atlantic. FEMS Microbiol. Ecol. 2000, 33, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Tarn, J.; Peoples, L.M.; Hardy, K.; Cameron, J.; Bartlett, D.H. Identification of free-living and particle-associated microbial communities present in Hadal regions of the Mariana Trench. Front. Microbiol. 2016, 7, 665. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, F.; Liu, H.; Jing, H. Metabolic activity and community structure of prokaryotes associated with particles in the twilight zone of the South China Sea. Front. Microbiol. 2022, 13, 1056860. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhang, Y.; Jing, H.; Liu, H. Spatial variation and metabolic diversity of microbial communities in the surface sediments of the Mariana Trench. Front. Microbiol. 2022, 13, 1051999. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.; Grossart, H.; Schweitzer, B.; Ploug, H. Microbial ecology of organic aggregates in aquatic ecosystems. Aquat. Microb. Ecol. 2002, 28, 175–211. [Google Scholar] [CrossRef]
- Leduc, D.; Rowden, A.A. Nematode communities in sediments of the Kermadec Trench, Southwest Pacific Ocean. Deep. Sea Res. Part I 2018, 134, 23–31. [Google Scholar] [CrossRef]
- Angel, M.V. Ocean trench conservation. Environmentalist 1982, 2, 1–17. [Google Scholar] [CrossRef]
- Feigl, V.; Ujaczki, É.; Vaszita, E.; Molnár, M. Influence of red mud on soil microbial communities: Application and comprehensive evaluation of the Biolog EcoPlate approach as a tool in soil microbiological studies. Sci. Total Environ. 2017, 595, 903–911. [Google Scholar] [CrossRef]
- Farkas, É.; Feigl, V.; Gruiz, K.; Vaszita, E.; Fekete-Kertész, I.; Tolner, M.; Kerekes, I.; Pusztai, É.; Kari, A.; Uzinger, N.; et al. Long-term effects of grain husk and paper fibre sludge biochar on acidic and calcareous sandy soils—A scale-up field experiment applying a complex monitoring toolkit. Sci. Total Environ. 2020, 731, 138988. [Google Scholar] [CrossRef]
- Tam, L.; Derry, A.; Kevan, P.; Trevors, J. Functional diversity and community structure of microorganisms in rhizosphere and non-rhizosphere Canadian arctic soils. Biodivers. Conserv. 2001, 10, 1933–1947. [Google Scholar] [CrossRef]
- Moore, J.C. “Diversity, Taxonomic Versus Functional” in Encyclopedia of Biodiversity, 2nd ed.; Levin, S., Ed.; Elsevier Academic Press: Amsterdam, The Netherlands, 2013; Volume 2, pp. 648–656. [Google Scholar]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M. Development of a prokaryotic universal primer for simultaneous analysis of bacteria and Archaea using next-generation sequencing. PLoS ONE 2014, 9, e105592. [Google Scholar] [CrossRef] [PubMed]
- Takai, K.; Horikoshi, K. Rapid detection and quantification of members of the archaeal community by quantitative PCR using fluorogenic probes. Appl. Environ. Microbiol. 2000, 66, 5066–5072. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Gonzalez Peña, A.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Revelle, W. Package ‘Psych’. The Comprehensive R Archive Network. 30 August 2015. Available online: https://cran.r-project.org/web/packages/psych/psych.pdf (accessed on 1 December 2023).
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An open source software for exploring and manipulating networks. Proc. Int. AAAI Conf. Web Soc. Media 2009, 3, 361–362. [Google Scholar] [CrossRef]
- Aßhauer, K.P.; Wemheuer, B.; Daniel, R.; Meinicke, P. Tax4Fun: Predicting functional profiles from metagenomic 16S rRNA data. Bioinformatics 2015, 31, 2882–2884. [Google Scholar] [CrossRef]
- Hammer, Y.; Harper, D.A.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol Electron 2001, 4, 1. [Google Scholar] [CrossRef]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [PubMed]
- Glud, R.N.; Wenzhöfer, F.; Middelboe, M.; Oguri, K.; Turnewitsch, R.; Canfield, D.E.; Kitazato, H. High rates of microbial carbon turnover in sediments in the deepest oceanic trench on Earth. Nat. Geosci. 2013, 6, 284–288. [Google Scholar] [CrossRef]
- Acinas, S.G.; Antón, J.; Rodríguez-Valera, F. Diversity of free-living and attached bacteria in offshore western Mediterranean waters as depicted by analysis of genes encoding 16S rRNA. Appl. Environ. Microbiol. 1999, 65, 514–522. [Google Scholar] [CrossRef] [PubMed]
- Pinhassi, J.; Berman, T. Differential growth response of colony-forming alpha and gamma-proteobacteria in dilution culture and nutrient addition experiments from Lake Kinneret (Israel), the eastern Mediterranean Sea, and the Gulf of Eilat. Appl. Environ. Microbiol. 2003, 69, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Peoples, L.M.; Donaldson, S.; Osuntokun, O.; Xia, Q.; Nelson, A.; Blanton, J.; Allen, E.E.; Church, M.J.; Bartlett, D.H. Vertically distinct microbial communities in the Mariana and Kermadec trenches. PLoS ONE 2018, 13, e0195102. [Google Scholar] [CrossRef]
- Gaba, S.; Kumari, A.; Medema, M.; Kaushik, R. Pan-genome analysis and ancestral state reconstruction of class halobacteria: Probability of a new super-order. Sci. Rep. 2020, 10, 21205. [Google Scholar] [CrossRef]
- Oren, A.; Oremland, R.S. Diversity of anaerobic halophilic microorganisms. In Instruments, Methods, and Missions for Astrobiology III; SPIE: San Diego, CA, USA, 2000; Volume 4137, pp. 96–105. [Google Scholar] [CrossRef]
- Zhang, A.; Carroll, A.L.; Atsumi, S. Carbon recycling by cyanobacteria: Improving CO2 fixation through chemical pro-duction. FEMS Microbiol. Lett. 2017, 364, fnx165. [Google Scholar] [CrossRef]
- Kong, L.-F.; He, Y.-B.; Xie, Z.-X.; Luo, X.; Zhang, H.; Yi, S.-H.; Lin, Z.-L.; Zhang, S.-F.; Yan, K.-Q.; Xu, H.-K.; et al. Illuminating key microbial players and metabolic processes involved in the remineralization of particulate organic carbon in the ocean’s twilight zone by metaproteomics. Appl. Environ. Microbiol. 2021, 87, AEM0098621. [Google Scholar] [CrossRef]
- Dai, H.; Zhang, Y.; Fang, W.; Liu, J.; Hong, J.; Zou, C.; Zhang, J. Microbial community structural response to variations in physicochemical features of different aquifers. Front. Microbiol. 2023, 14, 1025964. [Google Scholar] [CrossRef]
- Melita, M.; Amalfitano, S.; Preziosi, E.; Ghergo, S.; Frollini, E.; Parrone, D.; Zoppini, A. Physiological profiling and functional diversity of groundwater microbial communities in a municipal solid waste landfill area. Water 2019, 11, 2624. [Google Scholar] [CrossRef]
- Buchan, A.; LeCleir, G.R.; Gulvik, C.A.; Gonzalez, J.M. Master recyclers: Features and functions of bacteria associated with phytoplankton blooms. Nat. Rev. Microbiol. 2014, 12, 686–698. [Google Scholar] [CrossRef]
- Mishamandani, S.; Gutierrez, T.; Berry, D.; Aitken, M.D. Response of the bacterial community associated with a cosmopolitan marine diatom to crude oil shows a preference for the biodegradation of aromatic hydrocarbons. Environ. Microbiol. 2016, 18, 1817–1833. [Google Scholar] [CrossRef]
- Pacheco, A.R.; Moel, M.; Segrè, D. Costless metabolic secretions as drivers of interspecies interactions in microbial ecosystems. Nat. Commun. 2019, 10, 103. [Google Scholar] [CrossRef]
- Machado, D.; Maistrenko, O.M.; Andrejev, S.; Kim, Y.; Bork, P.; Patil, K.R.; Patil, K.R. Polarization of microbial communities between competitive and cooperative metabolism. Nat. Ecol. Evol. 2021, 5, 195–203. [Google Scholar] [CrossRef]
- Yang, C.; Lv, D.; Jiang, S.; Lin, H.; Sun, J.; Li, K.; Sun, J. Soil salinity regulation of soil microbial carbon metabolic function in the Yellow River Delta, China. Sci. Total Environ. 2021, 790, 148258. [Google Scholar] [CrossRef] [PubMed]
- Picazo, A.; Villaescusa, J.A.; Rochera, C.; Miralles-Lorenzo, J.; Quesada, A.; Camacho, A. Functional metabolic diversity of bacterioplankton in Maritime Antarctic Lakes. Microorganisms 2021, 9, 2077. [Google Scholar] [CrossRef]
- Urbani, R.; Magaletti, E.; Sist, P.; Cicero, A.M. Extracellular carbohydrates released by the marine diatoms Cylindrotheca closterium, Thalassiosira pseudonana and Skeletonema costatum: Effect of P-depletion and growth status. Sci. Total Environ. 2005, 353, 300–306. [Google Scholar] [CrossRef]
- Emborg, J.; Dalgaard, P. Formation of histamine and biogenic amines in cold-smoked tuna: An investigation of psychrotolerant bacteria from samples implicated in cases of histamine fish poisoning. J. Food Prot. 2006, 69, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Naila, A.; Flint, S.; Fletcher, G.; Bremer, P.; Meerdink, G. Control of biogenic amines in food—Existing and emerging approaches. J. Food Sci. 2010, 75, R139–R150. [Google Scholar] [CrossRef]
- López-González, J.; Suárez-Estrella, F.; Vargas-García, M.; López, M.; Jurado, M.; Moreno, J. Dynamics of bacterial microbiota during lignocellulosic waste composting: Studies upon its structure, functionality and biodiversity. Bioresour. Technol. 2015, 175, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Calcagno, V.; Jarne, P.; Loreau, M.; Mouquet, N.; David, P. Diversity spurs diversification in ecological communities. Nat. Commun. 2017, 8, 15810. [Google Scholar] [CrossRef] [PubMed]
- Hunt, D.E.; Ward, C.S. A network-based approach to disturbance transmission through microbial interactions. Front. Microbiol. 2015, 6, 1182. [Google Scholar] [CrossRef]
- Faust, K.; Raes, J. Microbial interactions: From networks to models. Nat. Rev. Microbiol. 2012, 10, 538–550. [Google Scholar] [CrossRef]
- Hoefs, M.; Schouten, S.; De Leeuw, J.W.; King, L.L.; Wakeham, S.G.; Damste, J. Ether lipids of planktonic archaea in the marine water column. Appl. Environ. Microbiol. 1997, 63, 3090–3095. [Google Scholar] [CrossRef]
- Parada, A.E.; Weber, P.K.; Mayali, X.; Fuhrman, J.A.; Pett-Ridge, J.; Dekas, A.E. Metatranscriptomic analysis of marine Thaumarchaea suggests intra-phylum variability in in situ heterotrophic carbon and nitrogen metabolim. In Proceedings of the AGU Fall Meeting Abstracts, San Francisco, CA, USA, 12–16 December 2016; p. B13J-03. [Google Scholar]
- Zhou, Z.; Tran, P.Q.; Kieft, K.; Anantharaman, K. Genome diversification in globally distributed novel marine Proteobacteria is linked to environmental adaptation. ISME J. 2020, 14, 2060–2077. [Google Scholar] [CrossRef]
- Gruber, D.F.; Simjouw, J.-P.; Seitzinger, S.P.; Taghon, G.L. Dynamics and characterization of refractory dissolved organic matter produced by a pure bacterial culture in an experimental predator-prey system. Appl. Environ. Microbiol. 2006, 72, 4184–4191. [Google Scholar] [CrossRef] [PubMed]
- Hoppe, H.-G.; Ullrich, S. Profiles of ectoenzymes in the Indian Ocean: Phenomena of phosphatase activity in the mesopelagic zone. Aquat. Microb. Ecol. 1999, 19, 139–148. [Google Scholar] [CrossRef]
- Tamburini, C.; Garcin, J.; Ragot, M.; Bianchi, A. Biopolymer hydrolysis and bacterial production under ambient hydrostatic pressure through a 2000 m water column in the NW Mediterranean. Deep. Sea Res. Part II 2002, 49, 2109–2123. [Google Scholar] [CrossRef]
- Huang, J.; Gao, K.; Yang, L.; Lu, Y. Successional action of Bacteroidota and Firmicutes in decomposing straw polymers in a paddy soil. Environ. Microbiome 2023, 18, 76. [Google Scholar] [CrossRef]
- van Bergeijk, D.A.; Terlouw, B.R.; Medema, M.H.; van Wezel, G.P. Ecology and genomics of Actinobacteria: New concepts for natural product discovery. Nat. Rev. Microbiol. 2020, 18, 546–558. [Google Scholar] [CrossRef]
- Teske, A.; Durbin, A.; Ziervogel, K.; Cox, C.; Arnosti, C. Microbial community composition and function in permanently cold seawater and sediments from an Arctic fjord of Svalbard. Appl. Environ. Microbiol. 2011, 77, 2008–2018. [Google Scholar] [CrossRef] [PubMed]
- Belyaev, G.M. Deep-Sea Ocean Trenches and Their Fauna; Nauka Publishing House: Moscow, Russia, 1989. [Google Scholar]
- Jamieson, A.J. The Hadal Zone: Life in the Deepest Oceans; Cambridge University Press: London, UK, 2015. [Google Scholar]
- Pandey, S.; Bhagawati, C.; Dandapat, S.; Chakraborty, A. Surface chlorophyll anomalies associated with Indian Ocean Dipole and El Niño Southern Oscillation in North Indian Ocean: A case study of 2006–2007 event. Environ. Monit. Assess. 2020, 191 (Suppl. S3), 807. [Google Scholar] [CrossRef] [PubMed]
- Galand, P.E.; Potvin, M.; Casamayor, E.O.; Lovejoy, C. Hydrography shapes bacterial biogeography of the deep Arctic Ocean. ISME J. 2009, 4, 564–576. [Google Scholar] [CrossRef] [PubMed]
- Buesseler, K.O.; Lamborg, C.H.; Boyd, P.W.; Lam, P.J.; Trull, T.W.; Bidigare, R.R.; Bishop, J.K.B.; Casciotti, K.L.; Dehairs, F.; Elskens, M.; et al. Revisiting Carbon Flux Through the Ocean’s Twilight Zone. Science 2007, 316, 567–570. [Google Scholar] [CrossRef]
- Mayor, D.J.; Cook, K.; Thornton, B.; Walsham, P.; Witte, U.F.M.; Zuur, A.F.; Anderson, T.R. Absorption efficiencies and basal turnover of C, N and fatty acids in a marine Calanoid copepod. Funct. Ecol. 2011, 25, 509–518. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Regions | Stns. | Longitude | Latitude | Depth (m) | Original Reads | Quality Reads | ASVs |
|---|---|---|---|---|---|---|---|
| Kermadec Trench | CTD-0 m | −176.4300 | −29.9167 | 0 | 76,240 | 23,118 | 207 |
| CTD-200 m | −176.4300 | −29.9167 | 200 | 63,214 | 28,839 | 318 | |
| CTD-800 m | −176.4300 | −29.9167 | 800 | 66,628 | 23,559 | 316 | |
| CTD-2000 m | −176.4300 | −29.9167 | 2000 | 81,419 | 25,578 | 371 | |
| CTD-5000 m | −176.4300 | −29.9167 | 5000 | 61,903 | 33,156 | 176 | |
| FDZ151 | −178.1515 | −27.0460 | 2311 | 67,341 | 39,850 | 194 | |
| FDZ152 | −178.1519 | −34.2276 | 6969 | 62,866 | 34,872 | 158 | |
| FDZ154 | −175.6629 | −27.4976 | 9639 | 63,346 | 36,404 | 193 | |
| FDZ156 | −177.1942 | −31.7540 | 9165 | 62,593 | 33,740 | 162 | |
| Diamantina Trench | CTD-0 m | 97.7227 | −32.5819 | 0 | 75,374 | 33,225 | 278 |
| CTD-200 m | 97.7227 | −32.5819 | 200 | 78,964 | 34,370 | 195 | |
| CTD-800 m | 97.7227 | −32.5819 | 800 | 79,762 | 39,750 | 166 | |
| CTD-2000 m | 97.7227 | −32.5819 | 2000 | 80,092 | 28,715 | 125 | |
| CTD-5000 m | 97.7227 | −32.5819 | 5000 | 74,029 | 32,712 | 173 | |
| FDZ172 | 95.8837 | −32.5689 | 5164 | 68,712 | 40,635 | 114 | |
| FDZ175 | 95.8820 | −32.5679 | 3060 | 71,551 | 46,621 | 114 | |
| FDZ182 | 101.2575 | −33.6463 | 6900 | 66,139 | 37,606 | 163 | |
| FDZ186 | 105.1887 | −35.8289 | 6594 | 76,933 | 49,103 | 115 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Jing, H. The Vertical Metabolic Activity and Community Structure of Prokaryotes along Different Water Depths in the Kermadec and Diamantina Trenches. Microorganisms 2024, 12, 708. https://doi.org/10.3390/microorganisms12040708
Liu H, Jing H. The Vertical Metabolic Activity and Community Structure of Prokaryotes along Different Water Depths in the Kermadec and Diamantina Trenches. Microorganisms. 2024; 12(4):708. https://doi.org/10.3390/microorganisms12040708
Chicago/Turabian StyleLiu, Hao, and Hongmei Jing. 2024. "The Vertical Metabolic Activity and Community Structure of Prokaryotes along Different Water Depths in the Kermadec and Diamantina Trenches" Microorganisms 12, no. 4: 708. https://doi.org/10.3390/microorganisms12040708
APA StyleLiu, H., & Jing, H. (2024). The Vertical Metabolic Activity and Community Structure of Prokaryotes along Different Water Depths in the Kermadec and Diamantina Trenches. Microorganisms, 12(4), 708. https://doi.org/10.3390/microorganisms12040708