
Molecular Identification and Subtype Analysis of Blastocystis sp. Isolates from Wild Mussels (Mytilus edulis) in Northern France
 , , , , ,
, , , , ,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Mussels Sampling Areas
2.2. Processing of Mussels
2.3. DNA Extraction and PCR Identification of Blastocystis sp.
2.4. Blastocystis sp. Isolates from Seawater Samples
2.5. Phylogenetic Analysis and Subtyping of Blastocystis sp. Isolates
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nemati, S.; Reza Zali, M.; Johnson, P.; Mirjalali, H.; Karanis, P. Molecular prevalence and subtype distribution of Blastocystis sp. in Asia and Australia. J. Water Health 2021, 19, 687. [Google Scholar] [CrossRef] [PubMed]
- Rauff-Adetotun, A.A.; Meor Termizi, F.H.; Shaari, N.; Lee, I.L. The coexistence of Blastocystis spp. in humans, animals and environmental sources from 2010–2021 in Asia. Biology 2021, 10, 990. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, P.; Munoz, M.; Ramirez, J.D. An update on the distribution of Blastocystis subtypes in the Americas. Heliyon 2023, 8, e12592. [Google Scholar] [CrossRef] [PubMed]
- Ning, C.-Q.; Hu, Z.-H.; Chen, J.-H.; Tian, L.-G. Epidemiology of Blastocystis infection from 1990 to 2019 in China. Infect. Dis. Poverty 2020, 9, 168. [Google Scholar] [CrossRef]
- Hublin, J.S.Y.; Maloney, J.G.; Santin, M. Blastocystis in domesticated and wild mammals and birds. Res. Vet. Sci. 2021, 135, 260–282. [Google Scholar] [CrossRef] [PubMed]
- Sanggari, A.; Komala, T.; Rauff-Adetotun, A.A.; Awosolu, O.B.; Attah, O.A.; Farah Haziqah, M.T. Blastocystis in captivated and free-ranging wild animals worldwide: A review. Trop. Biomed. 2022, 39, 338–372. [Google Scholar] [PubMed]
- Gantois, N.; Lamot, A.; Seesao, Y.; Creusy, C.; Li, L.L.; Monchy, S.; Benamrouz-Vanneste, S.; Karpouzopoulos, J.; Bourgain, J.L.; Rault, C.; et al. First report on the prevalence and subtype distribution of Blastocystis sp. in edible marine fish and marine mammals: A large scale-study conducted in Atlantic Northeast and on the coasts of Northern France. Microorganisms 2020, 8, 460. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Zhang, Y.; Qiao, H.; Li, S.; Wang, H.; Zhang, N.; Zhang, X. Cockroach as a vector of Blastocystis sp. is risk for Golden Monkeys in zoo. Korean J. Parasitol. 2020, 58, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Siwila, J.; Mwaba, F.; Chidumayo, N.; Mubanga, C. Food and waterborne protozoan parasites: The African perspective. Food Waterborne Parasitol. 2020, 20, e00088. [Google Scholar] [CrossRef] [PubMed]
- Khaled, S.; Gantois, N.; Tidjani Ly, A.; Senghor, S.; Even, G.; Dautel, E.; Dejager, R.; Sawant, M.; Baydoun, M.; Benamrouz-Vanneste, S.; et al. Prevalence and subtype distribution of Blastocystis sp. in Senegalese school children. Microorganisms 2020, 8, 1408. [Google Scholar] [CrossRef]
- Guilavogui, T.; Gantois, N.; Even, G.; Desramaut, J.; Dautel, E.; Denoyelle, C.; Cissé, F.I.; Touré, S.C.; Kourouma, B.L.; Sawant, M.; et al. Detection, molecular identification and transmission of the intestinal protozoa Blastocystis sp. in Guinea from a large-scale epidemiological study conducted in the Conakry area. Microorganisms 2022, 10, 446. [Google Scholar] [CrossRef] [PubMed]
- Naguib, D.; Gantois, N.; Desramaut, J.; Arafat, N.; Mandour, M.; Abdelmaogood, A.K.K.; Mosa, A.F.; Denoyelle, C.; Even, G.; Certad, G.; et al. Molecular epidemiology and genetic diversity of the enteric protozoan parasite Blastocystis sp. in the Northern Egypt population. Pathogens 2023, 12, 1359. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.S.W. New insights on classification, identification, and clinical relevance of Blastocystis spp. Clin. Microbiol. Rev. 2008, 21, 639–665. [Google Scholar] [CrossRef] [PubMed]
- Andersen, L.O.; Stensvold, C.R. Blastocystis in health and disease: Are we moving from a clinical to a public health perspective? J. Clin. Microbiol. 2016, 54, 524–528. [Google Scholar] [CrossRef] [PubMed]
- Stensvold, C.R.; Clark, C.G. Current status of Blastocystis: A personal view. Parasitol. Int. 2016, 65, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Fréalle, E.; El Safadi, D.; Cian, A.; Aubry, E.; Certad, G.; Osman, M.; Wacrenier, A.; Dutoit, E.; Creusy, C.; Dubos, F.; et al. Acute Blastocystis-associated appendicular peritonitis in a child, Casablanca, Morocco. Emerg. Infect. Dis. 2015, 21, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Angelici, M.C.; Nardis, C.; Scarpelli, R.; Ade, P. Blastocystis hominis transmission by non-potable water: A case report in Italy. New Microbiol. 2018, 41, 173–177. [Google Scholar] [PubMed]
- Aykur, M.; Camyar, A.; Türk, B.G.; Sin, A.Z.; Dagci, H. Evaluation of association with subtypes and alleles of Blastocystis with chronic spontaneous urticarial. Acta Trop. 2022, 231, 106455. [Google Scholar] [CrossRef] [PubMed]
- Ajjampur, S.S.; Tan, K.S.W. Pathogenic mechanisms in Blastocystis spp.—Interpreting results from in vitro and in vivo studies. Parasitol. Int. 2016, 65, 772–779. [Google Scholar] [CrossRef]
- Deng, L.; Wojciech, L.; Gascoigne, N.R.J.; Peng, G.; Tan, K.S.W. New insights into the interactions between Blastocystis, the gut microbiota, and host immunity. PLoS Pathog. 2021, 17, e1009253. [Google Scholar] [CrossRef]
- Deng, L.; Wojciech, L.; Png, C.W.; Koh, E.Y.; Aung, T.T.; Kioh, D.Y.Q.; Chan, E.C.Y.; Malleret, B.; Zhang, Y.; Peng, G.; et al. Experimental colonization with Blastocystis ST4 is associated with protective immune responses and modulation of gut microbiome in a DSS-induced colitis mouse model. Cell. Mol. Life Sci. 2022, 79, 245. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Lee, J.W.J.; Tan, K.S.W. Infection with pathogenic Blastocystis ST7 is associated with decreased bacterial diversity and altered gut microbiome profiles in diarrheal patients. Parasit. Vectors 2022, 15, 312. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Wojciech, L.; Png, C.W.; Kioh, Y.Q.D.; Ng, G.C.; Chan, E.C.Y.; Zhang, Y.; Gascoigne, N.R.J.; Tan, K.S.W. Colonization with ubiquitous protist Blastocystis ST1 ameliorates DSS-induced colitis and promotes beneficial microbiota and immune outcomes. NPJ Biofilms Microbiomes 2023, 9, 22. [Google Scholar] [CrossRef] [PubMed]
- Maloney, J.G.; Molokin, A.; Segui, R.; Maravilla, P.; Martinez-Hernandez, F.; Villalobos, G.; Tsaousis, A.D.; Gentekaki, E.; Munoz-Antoli, C.; Klisiowicz, D.R.; et al. Identification and molecular characterization of four new Blastocystis subtypes designated ST35-ST38. Microorganisms 2023, 11, 46. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Yao, Y.; Xiao, H.; Xie, M.; Xiong, Y.; Yang, S.; Ni, Q.; Zhang, M.; Xu, H. Extensive prevalence and significant genetic differentiation of Blastocystis in high- and low-altitude populations of wild rhesus macaques in China. Parasit. Vectors 2023, 16, 107. [Google Scholar] [CrossRef] [PubMed]
- Stensvold, C.R.; Berg, R.P.K.D.; Maloney, J.G.; Molokin, A.; Santin, M. Molecular characterization of Blastocystis and Entamoeba of muskoxen and sheep in Greenland. Int. J. Parasitol. 2023, 53, 673–685. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Castro, C.; Maloney, J.G.; Agudelo-López, S.P.; Toro-Londoño, M.A.; Botero-Garcés, J.H.; Orozco, M.C.; Quintero-Quinchia, Y.C.; Correa-Cote, J.C.; Múnera-Duque, A.; Ricaurte-Ciro, J.C.; et al. Identification and validation of novel Blastocystis subtype ST41 in a Colombian patient undergoing colorectal cancer screening. J. Eukaryot. Microbiol. 2023, 17, e12978. [Google Scholar] [CrossRef] [PubMed]
- Santin, M.; Figueiredo, A.; Molokin, A.; George, N.S.; Köster, P.C.; Dashti, A.; Gonzalez-Barrio, D.; Carmena, D.; Maloney, J.G. Division of Blastocystis ST10 into three new subtypes: ST42-ST44. J. Eukaryot. Microbiol. 2024, 71, e12998. [Google Scholar] [CrossRef] [PubMed]
- Koelher, A.V.; Herath, H.M.P.D.; Hall, R.S.; Wilcox, S. Marked genetic diversity within Blastocystis in Australian wildlife revealed using a next generation sequencing-phylogenetic approach. Int. J. Parasitol. Parasites Wildl. 2024, 23, 23–100902. [Google Scholar] [CrossRef]
- Stensvold, C.R.; Clark, C.G. Pre-empting Pandora’s box: Blastocystis subtypes revisited. Trends Parasitol. 2020, 36, 229–232. [Google Scholar] [CrossRef]
- Alfellani, M.A.; Stensvold, C.R.; Vidal-Lapiedra, A.; Onuoha, E.S.; Fagbenro-Beyioku, A.F.; Clark, C.G. Variable geographic distribution of Blastocystis subtypes and its potential implications. Acta Trop. 2013, 126, 11–18. [Google Scholar] [CrossRef]
- Ramírez, J.D.; Sánchez, A.; Hernández, C.; Florez, C.; Bernal, M.C.; Giraldo, J.C.; Reyes, P.; Lopez, M.C.; Garcia, L.; Cooper, P.J.; et al. Geographic distribution of human Blastocystis subtypes in South America. Infect. Genet. Evol. 2016, 41, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Stensvold, C.R.; Alfellani, M.A.; Norskov-Lauritsen, S.; Prip, K.; Victory, E.L.; Maddox, C.; Nielsen, H.V.; Clark, C.G. Subtype distribution of Blastocystis isolates from synanthropic and zoo animals and identification of a new subtype. Int. J. Parasitol. 2009, 39, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Köster, P.C.; Martinez-Nevado, E.; Gonzalez, A.; Abello-Poveda, M.T.; Fernandez-Bellon, H.; de la Riva-Fraga, M.; Marquet, B.; Guéry, J.-P.; Knauf-Witzens, T.; Weigold, A.; et al. Intestinal protists in captive non-human primates and their handlers in six European zoological gardens. Molecular evidence of zoonotic transmission. Front. Vet. Sci. 2021, 8, 819887. [Google Scholar] [CrossRef]
- Wang, W.; Owen, H.; Traub, R.J.; Cuttell, L.; Inpankaew, T.; Bielefeldt-Ohmann, H. Molecular epidemiology of Blastocystis in pigs and their in-contact humans in Southeast Queensland, Australia, and Cambodia. Vet. Parasitol. 2014, 203, 264–269. [Google Scholar] [CrossRef]
- Greige, S.; El Safadi, D.; Bécu, N.; Gantois, N.; Pereira, B.; Chabé, M.; Benamrouz-Vanneste, S.; Certad, G.; El Hage, R.; Chemaly, M.; et al. Prevalence and subtype distribution of Blastocystis sp. isolates from poultry in Lebanon and evidence of zoonotic potential. Parasit. Vectors 2018, 11, 389. [Google Scholar] [CrossRef] [PubMed]
- Giangaspero, A.; Marangi, M.; Stefania, M.; Annoscia, G.; Putignani, L.; Capelli, G.; Bonassisa, L. Efficiency of the Q3 lab-on-chip real time-PCR platform for detecting protozoan pathogens in bivalve mollusks. J. Food Sci. Technol. 2019, 56, 5000–5008. [Google Scholar] [CrossRef] [PubMed]
- Slodkowicz-Kowalska, A.; Majewska, A.C.; Rzymski, P.; Skrzypczak, L. Human waterborne protozoan parasites in freshwater bivalves (Anadonta anatina and Unio tumidus) as potential indicators of fecal pollution in urban reservoir. Limnologica 2015, 51, 32–36. [Google Scholar] [CrossRef]
- Suarez, P.; Vallejos-Almirall, A.; Fernandez, I.; Gonzalez-Chavarria, I.; Alonso, J.L.; Vidal, G. Identification of Cryptosporidium parvum and Blastocystis hominis subtype ST3 in Cholga mussel and treated sewage: Preliminary evidence of fecal contamination in harvesting area. Food Waterborne Parasitol. 2024, 34, e00214. [Google Scholar] [CrossRef]
- Poirier, P.; Wawrzyniak, I.; Albert, A.; El Alaoui, H.; Delbac, F.; Livrelli, V. Development and evaluation of a real-time PCR assay for detection and quantification of Blastocystis parasites in human stool samples: Prospective study of patients with hematological malignancies. J. Clin. Microbiol. 2011, 49, 975–983. [Google Scholar] [CrossRef]
- Genitsaris, S.; Monchy, S.; Viscogliosi, E.; Sime-Ngando, T.; Ferreira, S.; Christaki, U. Seasonal variations of marine protist community structure based on taxon-specific traits using the eastern English Channel as a model coastal system. FEMS Microbiol. Ecol. 2015, 91, fiv034. [Google Scholar] [CrossRef] [PubMed]
- Rachik, S.; Christaki, U.; Li, L.L.; Genitsaris, S.; Breton, E.; Monchy, S. Diversity and potential activity patterns of planktonic eukaryotic microbes in a mesoeutrophic coastal area (eastern English Channel). PLoS ONE 2018, 13, e0196987. [Google Scholar] [CrossRef] [PubMed]
- Guillou, L.; Bachar, D.; Audic, S.; Bass, D.; Berney, C.; Bittner, L.; Boutte, C.; Burgaud, G.; de Vargas, C.; Decelle, J.; et al. The Protist Ribosomal Reference database (PR2): A catalog of unicellular eukaryote Small Sub-Unit rRNA sequences with curated taxonomy. Nucleic Acids Res. 2013, 41, D597–D604. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Capella-Gutierrez, S.; Silla-Martinez, J.M.; Gabaldon, T. TrimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol Biol Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Barbera, P.; Kozlov, A.M.; Czech, L.; Morel, B.; Darriba, D.; Flouri, T.; Stamatakis, A. EPA-ng: Massively parallel evolutionary placement of genetic sequences. Syst. Biol. 2019, 68, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Czech, L.; Barbera, P.; Stamatakis, A. Genesis and Gappa: Processing, analyzing and visualizing phylogenetic (placement) data. Bioinformatics 2020, 36, 3263–3265. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Naguib, D.; Gantois, N.; Desramaut, J.; Arafat, N.; Even, G.; Certad, G.; Chabé, M.; Viscogliosi, E. Prevalence, subtype distribution and zoonotic significance of Blastocystis sp. isolates from poultry, cattle and pets in Northern Egypt. Microorganisms 2022, 10, 2259. [Google Scholar] [CrossRef] [PubMed]
- Santin, M.; Molokin, A.; Maloney, J.G. A longitudinal study of Blastocystis in dairy calves from birth through 24 months demonstrates dynamic shifts in infection rates and subtype prevalence and diversity by age. Parasit. Vectors 2023, 16, 177. [Google Scholar] [CrossRef] [PubMed]
- Audebert, C.; Gantois, N.; Ducrocq, S.; Darras, M.; Merlin, S.; Martel, S.; Viscogliosi, E.; Even, G.; Chabé, M. Animal, herd and feed characteristics associated with Blastocystis prevalence and molecular diversity in dairy cattle from the North of France. Parasitologia 2022, 2, 45–53. [Google Scholar] [CrossRef]
- Verin, F.; Devreker, D.; Lefebvre, A. Etude Sanitaire de la Zone 62.07 “Wimereux”—Département du Pas-de-Calais. ODE/RST/LER.BL/17.02. 2017. Available online: https://archimer.ifremer.fr/doc/00378/48874/ (accessed on 1 December 2023).
- Verin, F.; Caboche, J.; Lagache, C.; Lefebvre, A. Etude de la Qualité Microbiologique et Chimique de la Zone n° 62.10 « Baie de Canche: Hardelot/Le Touquet» Groupe 3. RST.LER.BL/09.08. 2009. Available online: https://archimer.ifremer.fr/doc/00684/79567/ (accessed on 1 December 2023).
- Hitier, B. Modélisation Mathématique du Transport, de la Dispersion et de la Dégradation des Rejets Bactériens sur le Littoral Nord-Pas de Calais-Picardie. 2010. Available online: https://archimer.ifremer.fr/doc/00082/19366/ (accessed on 1 December 2023).
- Ithoi, I.; Jali, A.; Mak, J.W.; Wan Sulaiman, W.Y.; Mahmud, R. Occurrence of Blastocystis in water of two rivers from recreational areas in Malaysia. J. Parasitol. Res. 2011, 2011, 123916. [Google Scholar] [CrossRef]

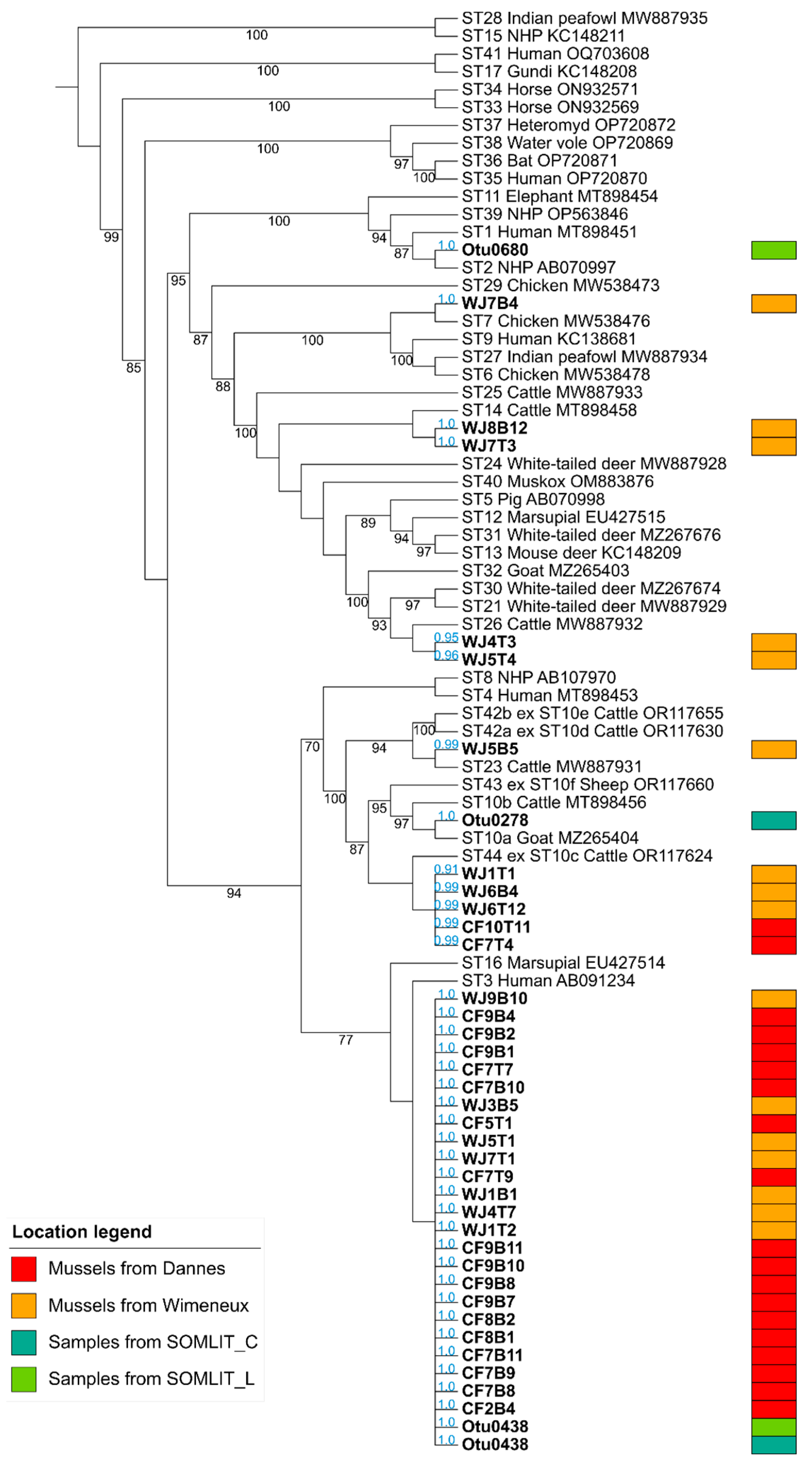

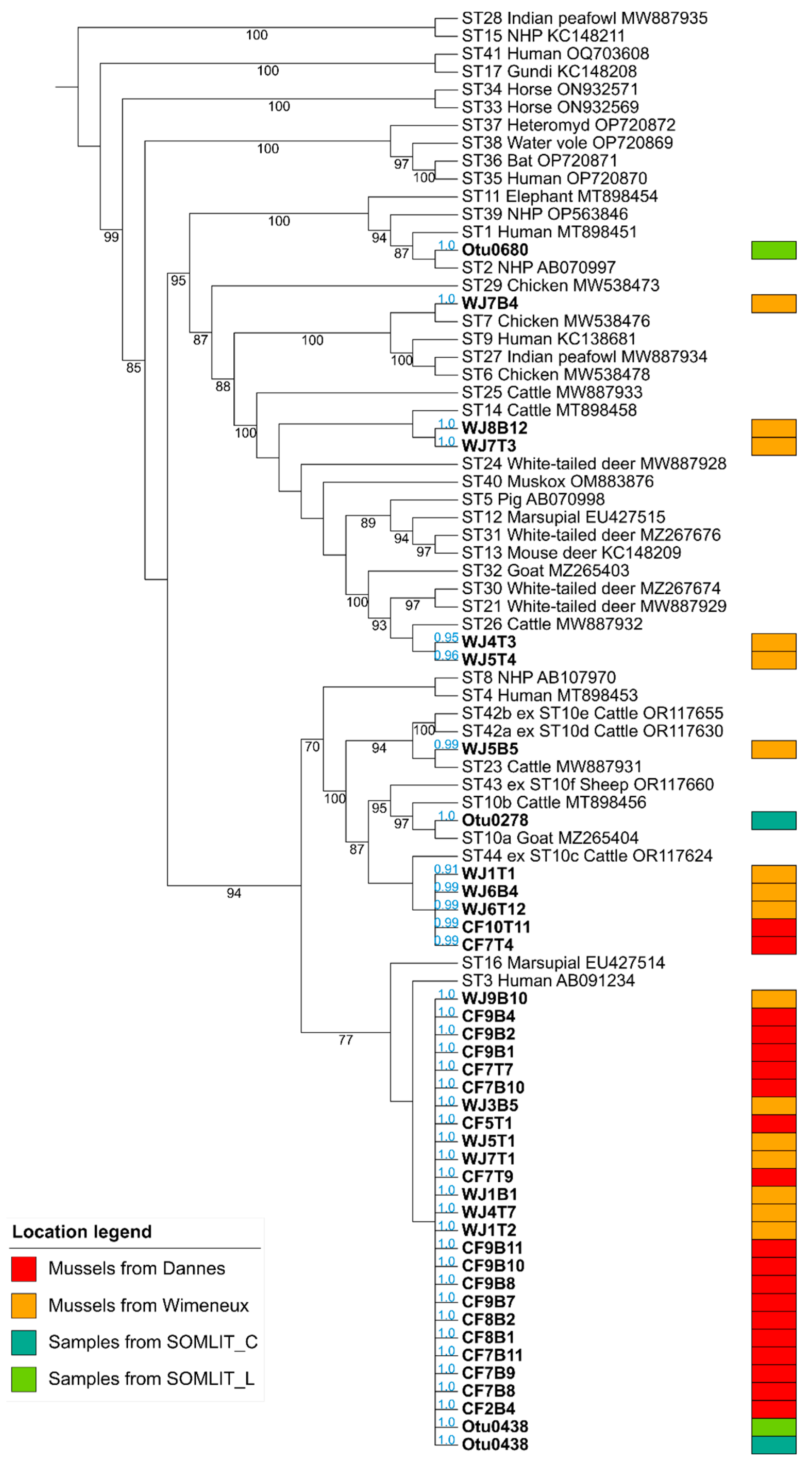
{kind=link}
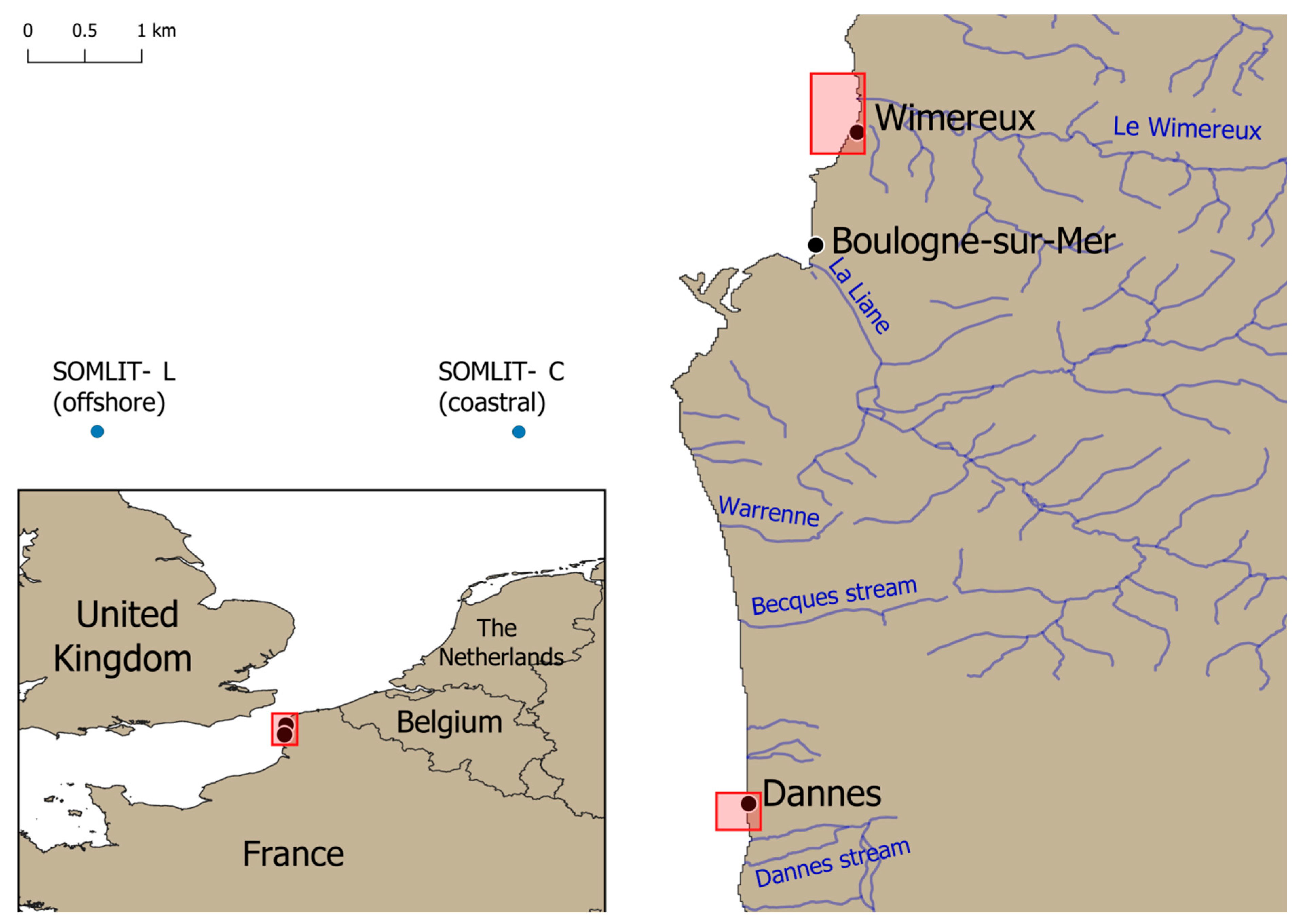
{kind=link}
| Collection Site | Gills | Gastrointestinal Tract | Both Organs | Infected Mussels (Gills and/or Gastrointestinal Tract) |
|---|---|---|---|---|
| Wimereux | 16/50 (32.0%) | 18/50 (36.0%) | 10/50 (20.0%) | 24/50 (48.0%) |
| Dannes | 29/50 (58.0%) | 19/50 (38.0%) | 10/50 (20%) | 38/50 (76.0%) |
| Total | 45/100 (45.0%) | 37/100 (37.0%) | 19/100 (19.0%) | 62/100 (62.0%) |
| Gills | Gastrointestinal Tract | ||||
|---|---|---|---|---|---|
| Sample | qPCR Assay | Blastocystis sp. ST | Sample | qPCR Assay | Blastocystis sp. ST |
| Wimereux | |||||
| WJ1B1 | + | ST3 | WJ1T1 | + | ST44 |
| WJ1B2 | - | WJ1T2 | + | ST3 | |
| WJ1B3 | + | MI a | WJ1T3 | - | |
| WJ2B2 | + | MI a | WJ2T2 | - | |
| WJ3B5 | + | ST3 | WJ3T5 | - | |
| WJ4B1 | + | MI a | WJ4T1 | + | MI a |
| WJ4B3 | - | WJ4T3 | + | ST26 | |
| WJ4B7 | + | MI a | WJ4T7 | + | ST3 |
| WJ5B1 | + | MI a | WJ5T1 | + | ST3 |
| WJ5B4 | - | WJ5T4 | + | ST26 | |
| WJ5B5 | + | ST23 | WJ5T5 | + | MI a |
| WJ6B4 | + | ST44 | WJ6T4 | - | |
| WJ6B12 | - | WJ6T12 | + | ST44 | |
| WJ7B1 | + | MI a | WJ7T1 | + | ST3 |
| WJ7B3 | + | MI a | WJ7T3 | + | ST14 |
| WJ7B4 | + | ST7 | WJ7T4 | + | MI a |
| WJ8B2 | + | MI a | WJ8T2 | - | |
| WJ8B12 | + | ST14 | WJ8T12 | - | |
| WJ9B10 | + | ST3 | WJ9T10 | + | MI a |
| WJ10B3 | - | WJ10T3 | + | MI a | |
| WJ10B4 | - | WJ10T4 | + | MI a | |
| WJ10B7 | - | WJ10T7 | + | MI a | |
| WJ10B8 | - | WJ10T8 | + | MI a | |
| WJ10B12 | + | MI a | WJ10T12 | + | MI a |
| Dannes | |||||
| CF1B2 | + | MI a | CF1T2 | + | MI a |
| CF1B5 | - | CF1T5 | + | MI a | |
| CF2B4 | + | ST3 | CF2T4 | - | |
| CF5B1 | + | MI a | CF5T1 | + | ST3 |
| CF7B4 | - | CF7T4 | + | ST44 | |
| CF7B5 | - | CF7T5 | + | MI a | |
| CF7B6 | + | MI a | CF7T6 | - | |
| CF7B7 | + | MI a | CF7T7 | + | ST3 |
| CF7B8 | + | ST3 | CF7T8 | - | |
| CF7B9 | + | ST3 | CF7T9 | + | ST3 |
| CF7B10 | + | ST3 | CF7T10 | - | |
| CF7B11 | + | ST3 | CF7T11 | - | |
| CF7B12 | + | MI a | CF7T12 | - | |
| CF8B1 | + | ST3 | CF8T1 | + | MI a |
| CF8B2 | + | ST3 | CF8T2 | + | MI a |
| CF8B3 | + | MI a | CF8T3 | + | MI a |
| CF8B4 | + | MI a | CF8T4 | + | MI a |
| CF8B5 | + | MI a | CF8T5 | - | |
| CF8B6 | + | MI a | CF8T6 | - | |
| CF8B7 | + | MI a | CF8T7 | + | MI a |
| CF8B8 | + | MI a | CF8T8 | - | |
| CF8B9 | + | MI a | CF8T9 | + | MI a |
| CF8B10 | + | MI a | CF8T10 | - | |
| CF8B11 | + | MI a | CF8T11 | - | |
| CF9B1 | + | ST3 | CF9T1 | - | |
| CF9B2 | + | ST3 | CF9T2 | - | |
| CF9B3 | + | MI a | CF9T3 | - | |
| CF9B4 | + | ST3 | CF9T4 | - | |
| CF9B7 | + | ST3 | CF9T7 | - | |
| CF9B8 | + | ST3 | CF9T8 | - | |
| CF9B10 | + | ST3 | CF9T10 | - | |
| CF9B11 | + | ST3 | CF9T11 | - | |
| CF10B1 | - | CF10T1 | + | MI a | |
| CF10B2 | - | CF10T2 | + | MI a | |
| CF10B7 | - | CF10T7 | + | MI a | |
| CF10B9 | - | CF10T9 | + | MI a | |
| CF10B10 | - | CF10T10 | + | MI a | |
| CF10B11 | - | CF10T11 | + | ST44 | |
| SOMLIT Station | OTU a | Number of Reads | Sampling Date | Season | Blastocystis sp. ST |
|---|---|---|---|---|---|
| C (coastal) | OTU0278 | 202 | 23/07/2012 | Summer | ST10a |
| 1 | 03/10/2012 | Autumn | |||
| OTU0438 | 14 | 21/06/2012 | Summer | ST3 | |
| 52 | 23/07/2012 | Summer | |||
| 2 | 13/11/2012 | Winter | |||
| L (offshore) | OTU0438 | 7 | 26/02/2013 | Winter | ST3 |
| 16 | 09/05/2012 | Spring | |||
| OTU0680 | 2 | 03/10/2012 | Autumn | ST2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryckman, M.; Gantois, N.; Dominguez, R.G.; Desramaut, J.; Li, L.-L.; Even, G.; Audebert, C.; Devos, D.P.; Chabé, M.; Certad, G.; et al. Molecular Identification and Subtype Analysis of Blastocystis sp. Isolates from Wild Mussels (Mytilus edulis) in Northern France. Microorganisms 2024, 12, 710. https://doi.org/10.3390/microorganisms12040710
Ryckman M, Gantois N, Dominguez RG, Desramaut J, Li L-L, Even G, Audebert C, Devos DP, Chabé M, Certad G, et al. Molecular Identification and Subtype Analysis of Blastocystis sp. Isolates from Wild Mussels (Mytilus edulis) in Northern France. Microorganisms. 2024; 12(4):710. https://doi.org/10.3390/microorganisms12040710
Chicago/Turabian StyleRyckman, Manon, Nausicaa Gantois, Ruben Garcia Dominguez, Jeremy Desramaut, Luen-Luen Li, Gaël Even, Christophe Audebert, Damien Paul Devos, Magali Chabé, Gabriela Certad, and et al. 2024. "Molecular Identification and Subtype Analysis of Blastocystis sp. Isolates from Wild Mussels (Mytilus edulis) in Northern France" Microorganisms 12, no. 4: 710. https://doi.org/10.3390/microorganisms12040710
APA StyleRyckman, M., Gantois, N., Dominguez, R. G., Desramaut, J., Li, L.-L., Even, G., Audebert, C., Devos, D. P., Chabé, M., Certad, G., Monchy, S., & Viscogliosi, E. (2024). Molecular Identification and Subtype Analysis of Blastocystis sp. Isolates from Wild Mussels (Mytilus edulis) in Northern France. Microorganisms, 12(4), 710. https://doi.org/10.3390/microorganisms12040710