
Impacts of Groundwater Pumping on Subterranean Microbial Communities in a Deep Aquifer Associated with an Accretionary Prism


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Sample Collection
2.2. Measurement of Physical and Chemical Parameters
2.3. Analysis of Stable Hydrogen and Oxygen Isotopic Ratios
2.4. Total Cell Count by the Total Direct Count Method
2.5. DNA Extraction
2.6. Next-Generation Sequencer (NGS) Analysis of 16S rRNA Genes
2.7. Clone Library Analysis of Archaeal 16S rRNA Genes
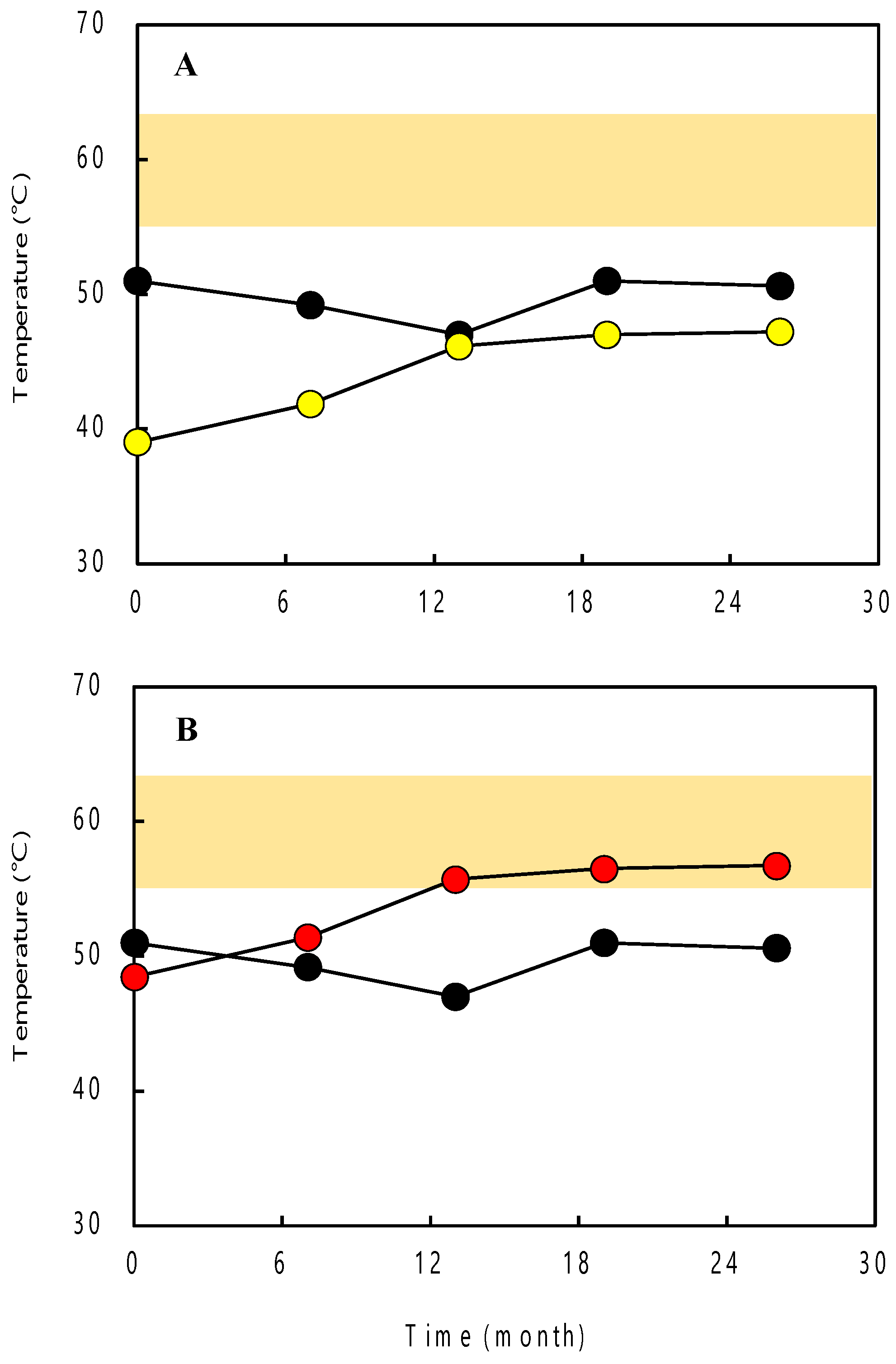
2.8. Estimation of Growth Temperatures
2.9. Measurement of Fermentation and CH4 Production Potential
3. Results
3.1. Physicochemical Signatures of Groundwater and Natural Gas
3.2. Stable Isotopic Signature of Groundwater
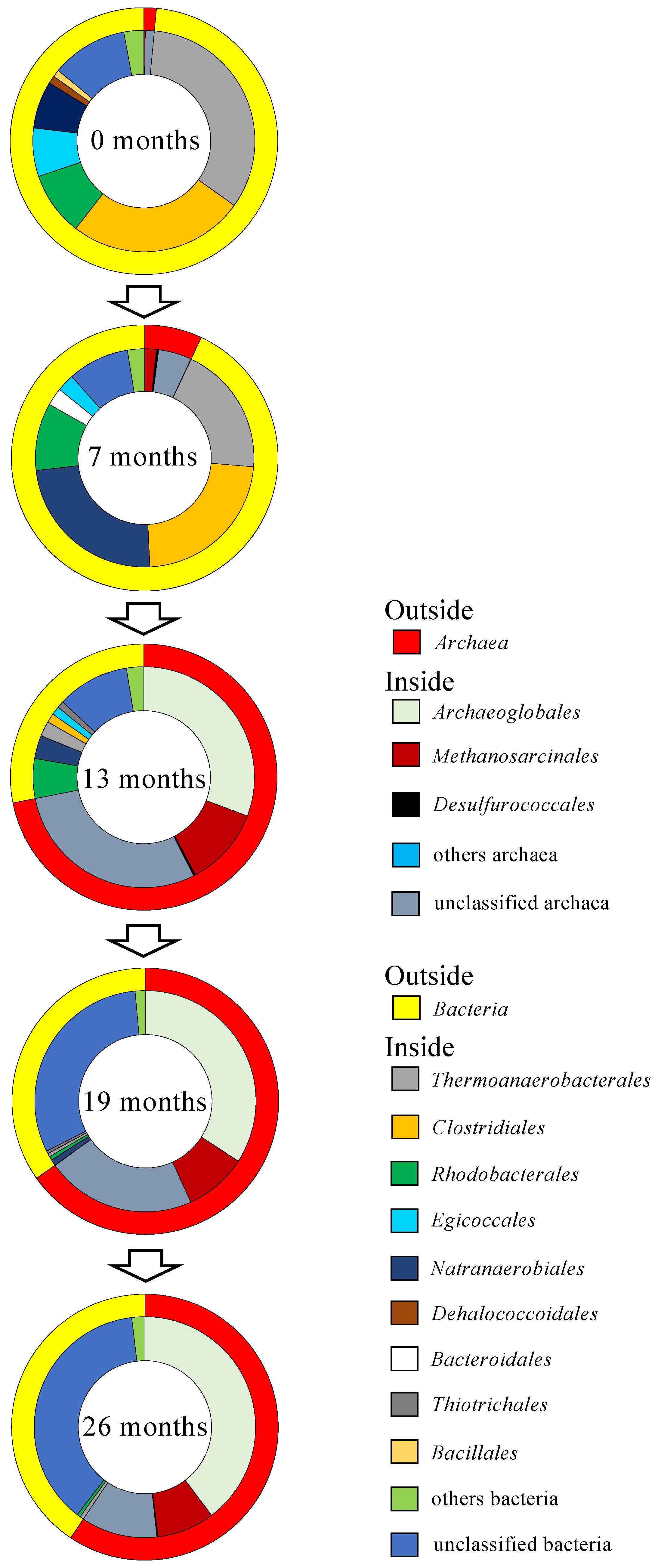
3.3. Abundance of Microbial Cells and Diversity of Prokaryotic Communities by NGS Analysis
3.4. Archaeal 16S rRNA Genes by Clone Library Analysis and Growth Temperature Estimation
3.5. Potential for H2-Producing Fermentative Bacteria and Methanogens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taira, A.; Byrne, T.; Ashi, J. Photographic Atlas of an Accretionary Prism: Geologic Structures of the Shimanto Belt, Japan; University of Tokyo Press: Tokyo, Japan, 1992; 130p. [Google Scholar]
- Kano, K.; Nakaji, M.; Takeuchi, S. Asymmetrical mélange fabrics as possible indicators of the convergent direction of plates: A case study from the Shimanto Belt of the Akaishi Mountains, central Japan. Tectonophysics 1991, 185, 375–399. [Google Scholar] [CrossRef]
- Reed, D.W.; Fujita, Y.; Delwiche, M.E.; Blackwelder, D.B.; Sheridan, P.P.; Uchida, T.; Colwell, F.S. Microbial communities from methane hydrate-bearing deep marine sediments in a Forearc Basin. Appl. Environ. Microbiol. 2002, 68, 3759–3770. [Google Scholar] [CrossRef] [PubMed]
- Hervé, F.; Calderón, M.; Fanning, C.M.; Pankhurst, R.J.; Godoy, E. Provenance variations in the Late Paleozoic accretionary complex of central Chile as indicated by detrital zircons. Gondwana Res. 2013, 23, 1122–1135. [Google Scholar] [CrossRef]
- Lee, H.; Fischer, T.P.; Maarten de Moor, J.; Sharp, Z.D.; Takahata, N.; Sano, Y. Nitrogen recycling at the Costa Rican subduction zone: The role of incoming plate structure. Sci. Rep. 2017, 7, 13933. [Google Scholar] [CrossRef] [PubMed]
- Maki, S.; Nagata, S.; Fukuta, O.; Furukawa, S. Geochemical study on organic matter from sedimentary rocks in the Miyazaki Group and the Shimanto Supergroup of Miyazaki Prefecture, Japan. Bull. Geol. Surv. Jpn. 1980, 31, 1–24. [Google Scholar]
- Sakata, S.; Maekawa, T.; Igari, S.; Sano, Y. Geochemistry and origin of natural gases dissolved in brines from gas fields in southwest Japan. Geofluids 2012, 12, 327–335. [Google Scholar] [CrossRef]
- Baito, K.; Imai, S.; Matsushita, M.; Otani, M.; Sato, Y.; Kimura, H. Biogas production using anaerobic groundwater containing a subterranean microbial community associated with the accretionary prism. Microb. Biotechnol. 2015, 8, 837–845. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H.; Nashimoto, H.; Shimizu, M.; Hattori, S.; Yamada, K.; Koba, K.; Yoshida, N.; Kato, K. Microbial methane production in deep aquifer associated with the accretionary prism in Southwest Japan. ISME J. 2010, 4, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, M.; Ishikawa, S.; Nagai, K.; Hirata, Y.; Ozawa, K.; Mitsunobu, S.; Kimura, H. Regional variation of CH4 and N2 production processes in the deep aquifers of an accretionary prism. Microbes Environ. 2016, 31, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Imachi, H. Topic of influence, methane and microbes. Microbes Environ. 2007, 32, 297–299. [Google Scholar] [CrossRef]
- Matsushita, M.; Magara, K.; Sato, Y.; Shinzato, N.; Kimura, H. Geochemical and microbiological evidence for microbial methane production in deep aquifers of the cretaceous accretionary prism. Microbes Environ. 2018, 33, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, H. Cultivation renaissance in the post-metagenomics era: Combining the new and old. Microbes Environ. 2019, 34, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, M.; Ishikawa, S.; Magara, K.; Sato, Y.; Kimura, H. The potential for CH4 production by syntrophic microbial communities in diverse deep aquifers associated with an accretionary prism and its overlying sedimentary layers. Microbes Environ. 2020, 35, ME19103. [Google Scholar] [CrossRef] [PubMed]
- Dawson, K.S.; Osburn, M.R.; Sessions, A.L.; Orphan, V.J. Metabolic associations with archaea drive shifts in hydrogen isotope fractionation in sulfate-reducing bacterial lipids in cocultures and methane seeps. Geobiology 2015, 13, 462–477. [Google Scholar] [CrossRef] [PubMed]
- Noble, R.T.; Fuhrman, J.A. Use of SYBR Green I for rapid epifluorescence counts of marine viruses and bacteria. Aquat. Microb. Ecol. 1998, 14, 113–118. [Google Scholar] [CrossRef]
- Yanagawa, K.; Tani, A.; Yamamoto, N.; Hachikubo, A.; Kano, A.; Matsumoto, R. Biogeochemical cycle of methanol in anoxic deep-sea sediments. Microbes Environ. 2016, 31, 190–193. [Google Scholar] [CrossRef] [PubMed]
- Vezzulli, L.; Pezzati, E.; Stauder, M.; Atagnaro, L.; Venier, P.; Pruzzo, C. Aquatic ecology of the oyster pathogens Vibrio splendidus and Vibrio aestuarianus. Environ. Microbiol. 2015, 17, 1065–1080. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H.; Ishibashi, J.; Masuda, H.; Kato, K.; Hanada, S. Selective phylogenetic analysis targeting 16S rRNA genes of hyperthermophilic archaea in the deep-subsurface hot biosphere. Appl. Environ. Microbiol. 2007, 73, 2110–2117. [Google Scholar] [CrossRef]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M. Development of a prokaryotic universal primer for simultaneous analysis of bacteria and archaea using next-generation sequencing. PLoS ONE 2014, 9, e105592. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H.; Mori, K.; Yamanaka, T.; Ishibashi, J. Growth temperatures of archaeal communities can be estimated from the guanine-plus-cytosine contents of 16S rRNA gene fragments. Environ. Microbiol. Rep. 2013, 5, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Huber, T.; Faulkner, G.; Hugenholtz, P. Bellerophon: A program to detect chimeric sequences in multiple sequence alignments. Bioinformatics 2004, 20, 2317–2319. [Google Scholar] [CrossRef] [PubMed]
- Good, I.J. The population frequencies of species and estimation of population parameters. Biometrika 1953, 40, 237–264. [Google Scholar] [CrossRef]
- Galtier, N.; Lobry, J.R. Relationships between genomic G+C content, RNA secondary structures, and optimal growth temperature in prokaryotes. J. Mol. Evol. 1997, 44, 632–636. [Google Scholar] [CrossRef] [PubMed]
- Khachane, A.N.; Timmis, K.N.; Dos Santos, V.A.P.M. Uracil content of 16S rRNA of thermophilic and psychrophilic prokaryotes correlates inversely with their optimal growth temperatures. Nucleic Acids Res. 2005, 33, 4016–4022. [Google Scholar] [CrossRef]
- Sato, Y.; Okano, K.; Kimura, H.; Honda, K. TEMPURA: Database of growth temperatures of usual and rare prokaryotes. Microbes Environ. 2020, 35, ME20074. [Google Scholar] [CrossRef]
- Tanaka, A.; Yamano, M.; Yano, Y.; Sasada, M. Geothermal gradient and heat flow data in and around Japan (I). Earth Planets Space 2004, 56, 1191–1194. [Google Scholar] [CrossRef]
- Craig, H. Isotope variations in meteoric waters. Science 1961, 133, 1702–1703. [Google Scholar] [CrossRef]
- Huber, H.; Stetter, K.O.; Order, I. Archaeoglobales ord. nov. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; The Archaea and the Deeply Branching and Phototrophic Bacteria; Garrity, G.M., Boone, D.R., Castenholz, R.W., Eds.; Springer: New York, NY, USA, 2001; Volume 1, p. 349. [Google Scholar]
- Brazelton, W.J.; Nelson, B.; Schrenk, M.O. Metagenomic evidence for H2 oxidation and H2 production by serpentinite-hosted subsurface microbial communities. Front. Microbiol. 2012, 2, 268. [Google Scholar] [CrossRef]
- Okonkwo, O.; Escudie, R.; Bernet, N.; Mangayil, R.; Lakaniemi, A.; Trably, E. Impacts of short-term temperature fluctuations on biohydrogen production and resilience of thermophilic microbial communities. Int. J. Hydrogen Energy 2019, 44, 8028–8037. [Google Scholar] [CrossRef]
- Mesbah, M.N.; Hedrick, B.D.; Peacock, D.A.; Rohde, M.; Wiegel, J. Natranaerobius thermophilus gen. nov., sp. nov., a halophilic, alkalithermophilic bacterium from soda lakes of the Wadi an Natrun, Egypt, and proposal of Natranaerobiaceae fam. nov. and Natranaerobiales ord. nov. Int. J. Syst. Evol. Microbiol. 2007, 57, 2507–2512. [Google Scholar] [CrossRef]
- Zavarzina, G.D.; Zhilina, N.T.; Kuznetsov, B.B.; Kolganova, V.T.; Osipov, A.G.; Kotelev, S.M.; Zavarzin, A.G. Natranaerobaculum magadiense gen. nov., sp. nov., an anaerobic, alkalithermophilic bacterium from soda lake sediment. Int. J. Syst. Evol. Microbiol. 2013, 63, 4456–4461. [Google Scholar] [CrossRef] [PubMed]
- Patel, G.B. Fermentation of lactose by Bacteroides polypragmatus. Can. J. Microbiol. 1983, 29, 120–128. [Google Scholar] [CrossRef]
- Orsi, E.; Beekwilder, J.; Eggink, G.; Kengen, W.M.G.; Weusthuis, A.R. The transition of Rhodobacter sphaeroides into a microbial cell factory. Biotechnol. Bioeng. 2020, 118, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Harirchi, S.; Sar, T.; Ramezani, M.; Aliyu, H.; Etemadifar, Z.; Nojoumi, S.A.; Yazdian, F.; Awasthi, M.K.; Taherzadeh, M.J. Bacillales: From taxonomy to biotechnological and industrial perspectives. Microorganisms 2022, 10, 2355. [Google Scholar] [CrossRef] [PubMed]
- Boone, D.R.; Whitman, B.W.; Koga, Y. Order III. Methanosarcinales ord. nov. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Boone, R.D., Castenholz, W.R., Garrity, M.G., Eds.; Springer: New York, NY, USA, 2001; Volume 1, p. 268. [Google Scholar]
- Balch, W.E.; Fox, G.E.; Magrum, L.J.; Woese, C.R.; Wolfe, R.S. Methanogens: Reevaluation of a unique biological group. Microbiol. Rev. 1979, 43, 260–296. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y. Methanococcales. In Handbook of Hydrocarbon and Lipid Microbiology; Springer: Berlin/Heidelberg, Germany, 2010; pp. 573–581. [Google Scholar]
- Huser, A.B.; Wuhrmann, K.; Zehnder, J.B.A. Methanothrix soehngenii gen. nov. sp. nov., a new acetotrophic non-hydrogen-oxidizing methane bacterium. Arch. Microbiol. 1982, 132, 1–9. [Google Scholar] [CrossRef]
- Kamagata, Y.; Kawasaki, H.; Oyaizu, H.; Nakamura, K.; Mikami, E.; Endo, G.; Koga, Y.; Yamasato, K. Characterization of three thermophilic strains of Methanothrix (“Methanosaeta”) thermophila sp. nov. and rejection of Methanothrix (“Methanosaeta”) thermoacetophila. Int. J. Syst. Bacteriol. 1992, 42, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Liu, X.; Dong, X. Methanosaeta harundinacea sp. nov., a novel acetate-scavenging methanogen isolated from a UASB reactor. Int. J. Syst. Evol. Microbiol. 2006, 56, 127–131. [Google Scholar] [CrossRef]
- Beeder, J.; Nilsen, R.K.; Rosnes, J.T.; Torsvik, T.; Lien, T. Archaeoglobus fulgidus isolated from hot North Sea oil field waters. Appl. Environ. Microbiol. 1994, 60, 1227–1231. [Google Scholar] [CrossRef] [PubMed]
- Hoefs, J. Stable Isotope Geochemistry; Springer: Berlin/Heidelberg, Germany, 2009; pp. 93–224. [Google Scholar]
- Mori, K.; Hanada, S.; Maruyama, A.; Marumo, K. Thermanaeromonas toyohensis gen. nov., sp. nov., a novel thermophilic anaerobe isolated from a subterranean vein in the Toyoha Mines. Int. J. Syst. Evol. Microbiol. 2002, 52, 1675–1680. [Google Scholar] [PubMed]
- Collins, M.D.; Laqson, P.A.; Willems, A.; Cordoba, J.J.; Fernandez-Garayzabal, J.; Garcia, P.; Cal, J.; Hippe, H.; Farrow, J.A.E. The phylogeny of the genus Clostridium: Proposal of five new genera and eleven new species combinations. Int. J. Syst. Evol. Microbiol. 1994, 44, 812–826. [Google Scholar] [CrossRef] [PubMed]
- Plugge, C.M.; Balk, M.; Zoetendal, E.G.; Stams, A.J.M. Gelria glutamica gen. nov., sp. nov., a thermophilic, obligately syntrophic, glutamate-degrading anaerobe. Int. J. Syst. Evol. Microbiol. 2002, 52, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, H.; Wangner, D.I.; Wiegel, J. Caldicoprobacter oshimai gen. nov., sp. nov., an anaerobic, xylanolytic, extremely thermophilic bacterium isolated from sheep faeces, and proposal of Caldicoprobacteraceae fam. nov. Int. J. Syst. Evol. Microbiol. 2010, 60, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Kiliops, V.; Kiliops, S. Long-term fate of organic matter in the geosphere. In An Introduction to Organic Geochemistry; Killops, S., Killops, V., Eds.; Blackwell Publishing: Malden, MA, USA, 2004; pp. 117–165. [Google Scholar]
- Hartzell, P.; Reed, D.W. The genus Archaeoglobus. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006. [Google Scholar]
- Stetter, K.O.; Lauerer, G.; Thomm, M.; Neuner, A. Isolation of extremely thermophilic sulfate reducers: Evidence for a novel branch of archaebacteria. Science 1987, 236, 822–824. [Google Scholar] [CrossRef] [PubMed]
- Burggraf, S.; Jannasch, W.H.; Nicolaus, B.; Stetter, O.K. Archaeoglobus profundus sp. nov., represents a new species within the sulfate-reducing archaebacteria. Syst. Appl. Microbiol. 1990, 13, 24–28. [Google Scholar] [CrossRef]
- Huber, H.; Jannasch, H.; Rachel, R.; Fuchs, T.; Stetter, K.O. Archaeoglobus veneficus sp. nov., a novel facultative chemolithoautotrophic hyperthermophilic sulfite reducer, isolated from Abyssal Black Smokers. Syst. Appl. Microbiol. 1997, 20, 374–380. [Google Scholar] [CrossRef]
- Schonheit, P.; Kristjansson, J.K.; Thauer, R.K. Kinetic mechanism for the ability of sulfate reducers to outcompete methanogens for acetate. Arch. Microbiol. 1982, 132, 285. [Google Scholar] [CrossRef]
- Ingvorsen, K.; Zehnder, A.J.B.; Jrgensen, B.B. Kinetic of sulfate and acetate uptake by Desulfobacter postgatei. Appl. Environ. Microbiol. 1984, 47, 403. [Google Scholar] [CrossRef] [PubMed]
- Widdel, F. New types of acetate-oxidizing, sulfate-reducing Desulfobacter species, D. hydrogenophilus sp. nov., D. latus sp. nov., and D. curvatus sp. nov. Arch. Microbiol. 1987, 148, 286–291. [Google Scholar]
- Hattori, S. Syntrophic acetate-oxidizing microbes in methanogenic environments. Microbes Environ. 2008, 23, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, J.; Zaramela, L.S.; Wang, L.; Mbadinga, S.M.; Hou, Z.; Wu, X.; Gu, J.; Zengler, K.; Mu, B. Genomic and transcriptomic evidence supports methane metabolism in Archaeoglobi. mSystems 2020, 5, e00651-19. [Google Scholar] [CrossRef]
- Groβkopf, R.; Janssen, P.H.; Liesack, W. Diversity and structure of the methanogenic community in anoxic rice paddy soil microcosms as examined by cultivation and direct 16S rRNA gene sequence retrieval. Appl. Environ. Microbiol. 1998, 64, 960–969. [Google Scholar] [CrossRef] [PubMed]
- Millero, F.J.; Feistel, R.; Wright, D.G.; McDougall, T.J. The composition of standard seawater and the definition of the reference-composition salinity scale. Deep Sea Res. Part 1 Oceanogr. 2008, 55, 50–72. [Google Scholar] [CrossRef]
- Kimura, H.; Sato, M.; Sugiyama, C.; Naganuma, T. Coupling of thraustochytrids and POM, and of bacterio- and phytoplankton in a semi-enclosed coastal area: Implication for different substrate preference by the planktonic decomposers. Aquat. Microb. Ecol. 2001, 25, 293–300. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Date | Groundwater | Natural Gas | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Temp. (°C) | pH | ORP 1 (mV) | EC 2 (mS m−1) | Cell Density (Cells mL−1) | N2 (Vol. %) | CH4 (Vol. %) | C2H6 (Vol. %) | ||
| 0 months after pumping initiation | 51.0 | 8.5 | −310 | 2780 | 3.0 × 103 | 1.40 | 98.6 | 0.030 | |
| 7 months after pumping initiation | 49.2 | 8.7 | −329 | 2730 | 2.1 × 10⁴ | 1.35 | 98.6 | 0.030 | |
| 13 months after pumping initiation | 47.0 | 8.6 | −252 | 2950 | 2.8 × 103 | 1.48 | 98.5 | 0.040 | |
| 19 months after pumping initiation | 51.0 | 8.5 | −363 | 2680 | 5.6 × 104 | 2.15 | 97.8 | 0.030 | |
| 26 months after pumping initiation | 50.6 | 8.6 | −460 | 2750 | 6.9 × 103 | 2.16 | 97.8 | 0.033 | |
| OTU 1 | Phylogenetic Order | No. of Clones | PG+C (%) | Tmin (°C) | Topt (°C) | Tmax (°C) |
|---|---|---|---|---|---|---|
| 0 months after pumping initiation | ||||||
| MNT1_A02 | Methanosarcinales | 48 | 56.1 | 18.3(±14.1) | 35.4(±14.8) | 44.8(±13.8) |
| MNT1_A03 | Thermofilales | 11 | 60.2 | 34.3(±14.6) | 52.7(±15.3) | 62.4(±14.3) |
| MNT1_A05 | Methanobacteriales | 4 | 54.7 | 12.8(±13.9) | 29.4(±14.6) | 38.8(±13.7) |
| MNT1_A06 | Methanobacteriales | 4 | 57.9 | 25.3(±14.3) | 43.0(±15.0) | 52.5(±14.1) |
| MNT1_A08 | Methanococcales | 3 | 56.5 | 19.8(±14.1) | 37.1(±14.8) | 46.5(±13.9) |
| MNT1_A09 | Archaeoglobales | 2 | 61.2 | 38.2(±14.7) | 57.0(±15.4) | 66.6(±14.5) |
| MNT1_A07 | Thermofilales | 1 | 60.6 | 35.8(±14.6) | 54.4(±15.3) | 64.1(±14.4) |
| 57.0 2 | 21.6(±14.2) 3 | 39.0(±14.9) 3 | 48.5(±13.9) 3 | |||
| 7 months after pumping initiation | ||||||
| MNT1_A02 | Methanosarcniales | 52 | 56.1 | 18.3(±14.1) | 35.4(±14.8) | 44.8(±13.8) |
| MNT1_A01 | Archaeoglobales | 24 | 59.9 | 33.1(±14.5) | 51.5(±15.3) | 61.1(±14.3) |
| MNT1_A03 | Thermofilales | 9 | 60.2 | 34.3(±14.6) | 52.7(±15.3) | 62.4(±14.3) |
| MNT1_A04 | Archaeoglobales | 3 | 59.2 | 30.4(±14.4) | 48.5(±15.2) | 58.1(±14.2) |
| MNT1_A10 | Methanococcales | 2 | 58.1 | 26.1(±14.3) | 43.8(±15.0) | 53.4(±14.1) |
| 57.7 2 | 24.4(±14.3) 3 | 42.0(±15.0) 3 | 51.5(±14.0) 3 | |||
| 13 months after pumping initiation | ||||||
| MNT1_A01 | Archaeoglobales | 52 | 59.9 | 33.1(±14.5) | 51.5(±15.3) | 61.1(±14.3) |
| MNT1_A02 | Methanosarcinales | 29 | 56.1 | 18.3(±14.1) | 35.4(±14.8) | 44.8(±13.8) |
| MNT1_A04 | Archaeoglobales | 9 | 59.2 | 30.4(±14.4) | 48.5(±15.2) | 58.1(±14.2) |
| MNT1_A03 | Thermofilales | 2 | 60.2 | 34.3(±14.6) | 52.7(±15.3) | 62.4(±14.3) |
| 58.6 2 | 28.2(±14.4) 3 | 46.2(±15.1) 3 | 55.7(±14.1) 3 | |||
| 19 months after pumping initiation | ||||||
| MNT1_A01 | Archaeoglobales | 53 | 59.9 | 33.1(±14.5) | 51.5(±15.3) | 61.1(±14.3) |
| MNT1_A02 | Methanosarcinales | 17 | 56.1 | 18.3(±14.1) | 35.4(±14.8) | 44.8(±13.8) |
| MNT1_A04 | Archaeoglobales | 14 | 59.2 | 30.4(±14.4) | 48.5(±15.2) | 58.1(±14.2) |
| MNT1_A03 | Thermofilales | 1 | 60.2 | 34.3(±14.6) | 52.7(±15.3) | 62.4(±14.3) |
| 59.0 2 | 29.7(±14.4) 3 | 47.8(±15.1) 3 | 57.4(±14.2) 3 | |||
| 26 months after pumping initiation | ||||||
| MNT1_A01 | Archaeoglobales | 36 | 59.9 | 33.1(±14.5) | 51.5(±15.3) | 61.1(±14.3) |
| MNT1_A04 | Archaeoglobales | 35 | 59.2 | 30.4(±14.4) | 48.5(±15.2) | 58.1(±14.2) |
| MNT1_A02 | Methanosarcinales | 16 | 56.1 | 18.3(±14.1) | 35.4(±14.8) | 44.8(±13.8) |
| MNT1_A07 | Thermofilales | 2 | 60.6 | 35.8(±14.6) | 54.4(±15.3) | 64.1(±14.4) |
| 59.0 2 | 29.7(±14.4) 3 | 47.8(±15.1) 3 | 57.4(±14.2) 3 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iso, S.; Sato, Y.; Kimura, H. Impacts of Groundwater Pumping on Subterranean Microbial Communities in a Deep Aquifer Associated with an Accretionary Prism. Microorganisms 2024, 12, 679. https://doi.org/10.3390/microorganisms12040679
Iso S, Sato Y, Kimura H. Impacts of Groundwater Pumping on Subterranean Microbial Communities in a Deep Aquifer Associated with an Accretionary Prism. Microorganisms. 2024; 12(4):679. https://doi.org/10.3390/microorganisms12040679
Chicago/Turabian StyleIso, Shinsei, Yu Sato, and Hiroyuki Kimura. 2024. "Impacts of Groundwater Pumping on Subterranean Microbial Communities in a Deep Aquifer Associated with an Accretionary Prism" Microorganisms 12, no. 4: 679. https://doi.org/10.3390/microorganisms12040679
APA StyleIso, S., Sato, Y., & Kimura, H. (2024). Impacts of Groundwater Pumping on Subterranean Microbial Communities in a Deep Aquifer Associated with an Accretionary Prism. Microorganisms, 12(4), 679. https://doi.org/10.3390/microorganisms12040679