

Intestinal IgA-Coated Bacteria in Healthy- and Altered-Microbiomes (Dysbiosis) and Predictive Value in Successful Fecal Microbiota Transplantation

 ,
,
Abstract
1. Introduction and Methods
2. Origin and Production of Intestinal IgA
3. The Immune Microbiome
4. Factors Important in IgA Coating of Intestinal Bacteria
5. Targets of IgA Coating on Bacteria
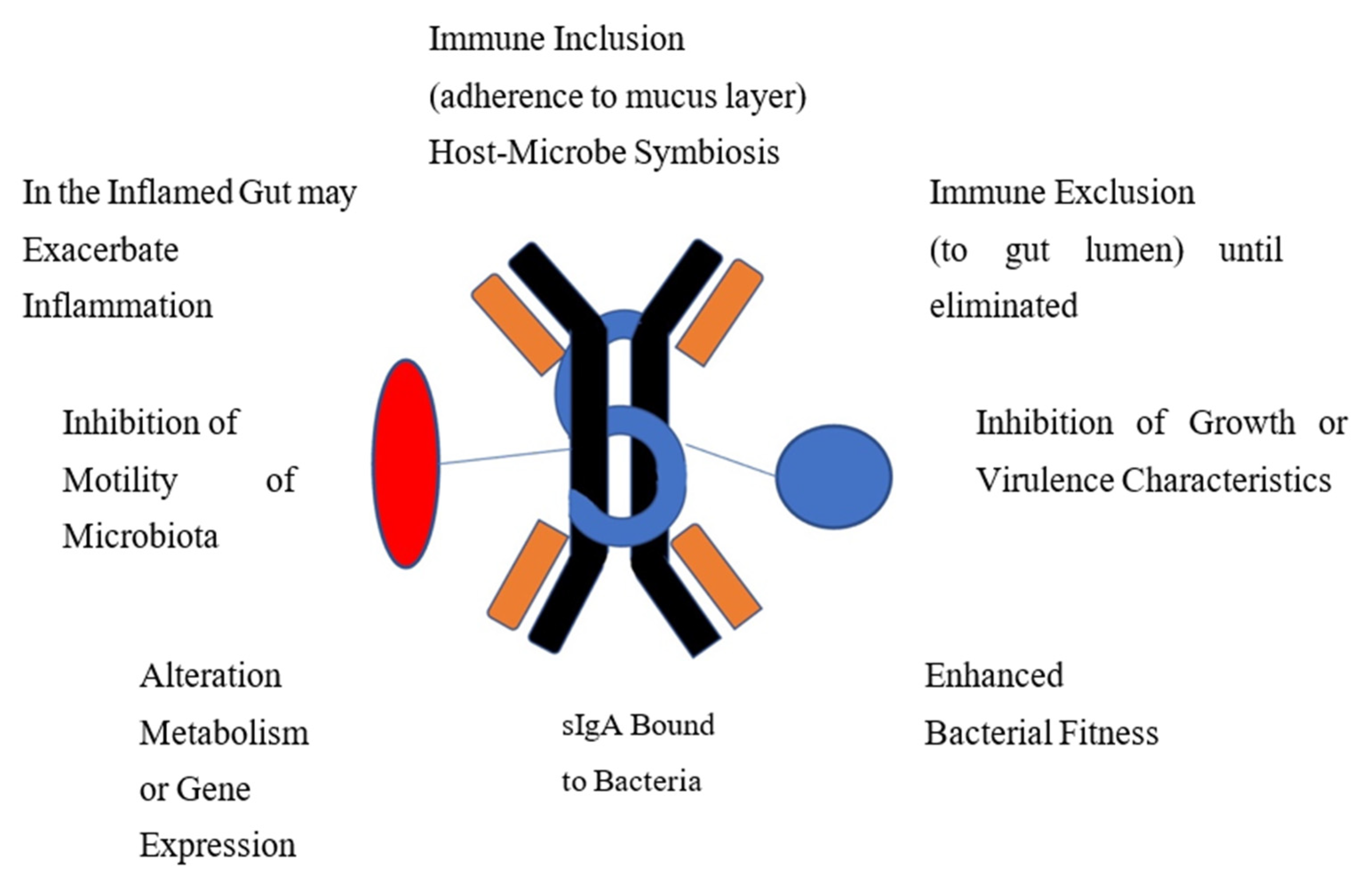
6. Function of IgA Coated Bacteria
7. Immunoglobulin (Ig)-Coating of Bacteria in the Gut in Conditions with Reduced Microbiota Diversity (Dysbiosis), Conditions Potentially Modified by Microbial Restoration with Fecal Microbiota Transplantation
7.1. Inflammatory Bowel Disease (IBD)
7.2. Enteric Infection
7.3. Celiac Disease in Children
7.4. Childhood Allergies and Asthma
7.5. Undernutrition
7.6. Obesity and Type 2 Diabetes
7.7. Clostridioides Difficile Infection (CDI)
7.8. Irritable Bowel Syndrome (IBS)
7.9. Multiple Sclerosis (MS)
7.10. Breast Cancer
7.11. Summary of Targets and Effects of the Immune-biome in Disorders of Dysbiosis
8. IgA-Biome before and after FMT in Disorders Associated with Dysbiosis
8.1. Inflammatory Bowel Disease
8.2. Recurrent Clostridioides Difficile Infection (CDI)
8.3. Systemic Sclerosis
8.4. Aging of Germinal Centers and Reduction of Intestinal IgA Produced

{kind=link}
{kind=link}
| Condition | Findings | Comment |
|---|---|---|
| IBD | The percent of IgA/IgG-coated bacteria was elevated in IBD. FMT shifted the IgA-enriched bacteria to that of the donors [61]. Route of FMT (oral or colonoscopy) did not influence the pattern of IgA/IgG-coated bacteria. Diversity of the IgA-biome at 4 weeks after FMT was more diverse and the diversity correlated with clinical response in UC [62]. Of the donor taxa only Osoribacter splanchnicus was significantly correlated with clinical improvement in one study [62]. The IgA-biome is effective in preventing colitis in mice exposed to dextran-sodium-sulfate (DDS), an IBD model [32,63]. | FMT shifted the IgA enrichment of bacteria to that of the donor in patients with UC. Phylogenetically diverse IgA-coated bacteria were transferred from donors to recipients. Diversity of the IgA-biome four weeks after FMT correlated with clinical response. |
| Recurrent CDI | Proinflammatory Proteobacteria strains were the most highly bound to IgA. Post-FMT, the IgA-biome of patients mirrored the donors [55]. | The dysbiosis in these patients relates to multiple antibiotics received. Replacing the microbiota with healthy taxa coated by IgA is curative in most cases. |
| Systemic Sclerosis | Abundance, richness and diversity of IgA-coated and IgM-coated bacteria fluctuated dynamically after transplantation in the FMT group, not in the placebo, and GI symptoms were improved [64]. | More studies are needed with systemic sclerosis to determine the value of FMT in disease management. |
| Aging | Germinal centers (GC), important in the production of IgA, lose functional capacity with aging. Advanced age mice, treated with FMT and cholera toxin vaccine experienced an increase in functional GCs [65]. | Impaired immune (IgA) reactivity of gut associated lymphoid tissues (GALT) in aging mice was shown to be reversible by FMT from younger aged donors. |
9. Summary and Comments
9.1. Five Scenarios May Best Explain the Biology of the IgA-Biome in Different Clinical Settings
9.2. New Studies Suggested Evaluating IgA-Biome in Fecal Microbiota Transplantation
9.3. Review Limitations
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fagarasan, S.; Kawamoto, S.; Kanagawa, O.; Suzuki, K. Adaptive immune regulation in the gut: T cell-dependent and T cell-independent IgA synthesis. Annu. Rev. Immunol. 2010, 28, 243–273. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Fagarasan, S. Diverse regulatory pathways for IgA synthesis in the gut. Mucosal Immunol. 2009, 2, 468–471. [Google Scholar] [CrossRef] [PubMed]
- Gutzeit, C.; Magri, G.; Cerutti, A. Intestinal IgA production and its role in host-microbe interaction. Immunol. Rev. 2014, 260, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Steinman, R.M. Decisions about dendritic cells: Past, present, and future. Annu. Rev. Immunol. 2012, 30, 1–22. [Google Scholar] [CrossRef]
- Esterhazy, D.; Loschko, J.; London, M.; Jove, V.; Oliveira, T.Y.; Mucida, D. Classical dendritic cells are required for dietary antigen-mediated induction of peripheral T(reg) cells and tolerance. Nat. Immunol. 2016, 17, 545–555. [Google Scholar] [CrossRef]
- Bunker, J.J.; Flynn, T.M.; Koval, J.C.; Shaw, D.G.; Meisel, M.; McDonald, B.D.; Ishizuka, I.E.; Dent, A.L.; Wilson, P.C.; Jabri, B.; et al. Innate and Adaptive Humoral Responses Coat Distinct Commensal Bacteria with Immunoglobulin A. Immunity 2015, 43, 541–553. [Google Scholar] [CrossRef]
- Zeng, M.Y.; Cisalpino, D.; Varadarajan, S.; Hellman, J.; Warren, H.S.; Cascalho, M.; Inohara, N.; Nunez, G. Gut Microbiota-Induced Immunoglobulin G Controls Systemic Infection by Symbiotic Bacteria and Pathogens. Immunity 2016, 44, 647–658. [Google Scholar] [CrossRef]
- Koch, M.A.; Reiner, G.L.; Lugo, K.A.; Kreuk, L.S.; Stanbery, A.G.; Ansaldo, E.; Seher, T.D.; Ludington, W.B.; Barton, G.M. Maternal IgG and IgA Antibodies Dampen Mucosal T Helper Cell Responses in Early Life. Cell 2016, 165, 827–841. [Google Scholar] [CrossRef]
- Fadlallah, J.; El Kafsi, H.; Sterlin, D.; Juste, C.; Parizot, C.; Dorgham, K.; Autaa, G.; Gouas, D.; Almeida, M.; Lepage, P.; et al. Microbial ecology perturbation in human IgA deficiency. Sci. Transl. Med. 2018, 10, eaan1217. [Google Scholar] [CrossRef]
- Catanzaro, J.R.; Strauss, J.D.; Bielecka, A.; Porto, A.F.; Lobo, F.M.; Urban, A.; Schofield, W.B.; Palm, N.W. IgA-deficient humans exhibit gut microbiota dysbiosis despite secretion of compensatory IgM. Sci. Rep. 2019, 9, 13574. [Google Scholar] [CrossRef]
- Mohammed, N.; Tang, L.; Jahangiri, A.; de Villiers, W.; Eckhardt, E. Elevated IgG levels against specific bacterial antigens in obese patients with diabetes and in mice with diet-induced obesity and glucose intolerance. Metabolism 2012, 61, 1211–1214. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Harmsen, H.J.; Pouwels, S.D.; Funke, A.; Bos, N.A.; Dijkstra, G. Crohn’s disease patients have more IgG-binding fecal bacteria than controls. Clin. Vaccine Immunol. 2012, 19, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Benckert, J.; Schmolka, N.; Kreschel, C.; Zoller, M.J.; Sturm, A.; Wiedenmann, B.; Wardemann, H. The majority of intestinal IgA+ and IgG+ plasmablasts in the human gut are antigen-specific. J. Clin. Investig. 2011, 121, 1946–1955. [Google Scholar] [CrossRef]
- Cerutti, A. The regulation of IgA class switching. Nat. Rev. Immunol. 2008, 8, 421–434. [Google Scholar] [CrossRef]
- Woof, J.M.; Kerr, M.A. The function of immunoglobulin A in immunity. J. Pathol. 2006, 208, 270–282. [Google Scholar] [CrossRef]
- D’Auria, G.; Peris-Bondia, F.; Dzunkova, M.; Mira, A.; Collado, M.C.; Latorre, A.; Moya, A. Active and secreted IgA-coated bacterial fractions from the human gut reveal an under-represented microbiota core. Sci. Rep. 2013, 3, 3515. [Google Scholar] [CrossRef]
- Kastl, A.J., Jr.; Terry, N.A.; Wu, G.D.; Albenberg, L.G. The Structure and Function of the Human Small Intestinal Microbiota: Current Understanding and Future Directions. Cell Mol. Gastroenterol. Hepatol. 2020, 9, 33–45. [Google Scholar] [CrossRef]
- O’Hara, A.M.; Shanahan, F. The gut flora as a forgotten organ. EMBO Rep. 2006, 7, 688–693. [Google Scholar] [CrossRef]
- Peach, S.; Lock, M.R.; Katz, D.; Todd, I.P.; Tabaqchali, S. Mucosal-associated bacterial flora of the intestine in patients with Crohn’s disease and in a control group. Gut 1978, 19, 1034–1042. [Google Scholar] [CrossRef]
- Sekirov, I.; Russell, S.L.; Antunes, L.C.; Finlay, B.B. Gut microbiota in health and disease. Physiol. Rev. 2010, 90, 859–904. [Google Scholar] [CrossRef]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Sterlin, D.; Fadlallah, J.; Slack, E.; Gorochov, G. The antibody/microbiota interface in health and disease. Mucosal Immunol. 2020, 13, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.A.; Pearson, C.; Ilott, N.E.; Huus, K.E.; Hegazy, A.N.; Webber, J.; Finlay, B.B.; Macpherson, A.J.; Powrie, F.; Lam, L.H. Accurate identification and quantification of commensal microbiota bound by host immunoglobulins. Microbiome 2021, 9, 33. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, A.; Vogelzang, A.; Maruya, M.; Miyajima, M.; Murata, M.; Son, A.; Kuwahara, T.; Tsuruyama, T.; Yamada, S.; Matsuura, M.; et al. IgA regulates the composition and metabolic function of gut microbiota by promoting symbiosis between bacteria. J. Exp. Med. 2018, 215, 2019–2034. [Google Scholar] [CrossRef] [PubMed]
- Joglekar, P.; Ding, H.; Canales-Herrerias, P.; Pasricha, P.J.; Sonnenburg, J.L.; Peterson, D.A. Intestinal IgA Regulates Expression of a Fructan Polysaccharide Utilization Locus in Colonizing Gut Commensal Bacteroides thetaiotaomicron. mBio 2019, 10, e02324-19. [Google Scholar] [CrossRef] [PubMed]
- Henke, M.T.; Brown, E.M.; Cassilly, C.D.; Vlamakis, H.; Xavier, R.J.; Clardy, J. Capsular polysaccharide correlates with immune response to the human gut microbe Ruminococcus gnavus. Proc. Natl. Acad. Sci. USA 2021, 118, e2007595118. [Google Scholar] [CrossRef]
- Bunker, J.J.; Erickson, S.A.; Flynn, T.M.; Henry, C.; Koval, J.C.; Meisel, M.; Jabri, B.; Antonopoulos, D.A.; Wilson, P.C.; Bendelac, A. Natural polyreactive IgA antibodies coat the intestinal microbiota. Science 2017, 358, aan6619. [Google Scholar] [CrossRef]
- Chen, K.; Magri, G.; Grasset, E.K.; Cerutti, A. Rethinking mucosal antibody responses: IgM, IgG and IgD join IgA. Nat. Rev. Immunol. 2020, 20, 427–441. [Google Scholar] [CrossRef]
- Mantis, N.J.; Rol, N.; Corthesy, B. Secretory IgA’s complex roles in immunity and mucosal homeostasis in the gut. Mucosal Immunol. 2011, 4, 603–611. [Google Scholar] [CrossRef]
- Okai, S.; Usui, F.; Yokota, S.; Hori, I.Y.; Hasegawa, M.; Nakamura, T.; Kurosawa, M.; Okada, S.; Yamamoto, K.; Nishiyama, E.; et al. High-affinity monoclonal IgA regulates gut microbiota and prevents colitis in mice. Nat. Microbiol. 2016, 1, 16103. [Google Scholar] [CrossRef]
- Weis, A.M.; Round, J.L. Microbiota-antibody interactions that regulate gut homeostasis. Cell Host Microbe 2021, 29, 334–346. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Chen, H.; Shu, W.; Sun, M.; Fang, L.; Shi, Y.; Pang, Z.; Wu, W.; Liu, Z. Clinical significance of soluble immunoglobulins A and G and their coated bacteria in feces of patients with inflammatory bowel disease. J. Transl. Med. 2018, 16, 359. [Google Scholar] [CrossRef]
- Masu, Y.; Kanazawa, Y.; Kakuta, Y.; Shimoyama, Y.; Onodera, M.; Naito, T.; Moroi, R.; Kuroha, M.; Kimura, T.; Shiga, H.; et al. Immunoglobulin subtype-coated bacteria are correlated with the disease activity of inflammatory bowel disease. Sci. Rep. 2021, 11, 16672. [Google Scholar] [CrossRef] [PubMed]
- Palm, N.W.; de Zoete, M.R.; Cullen, T.W.; Barry, N.A.; Stefanowski, J.; Hao, L.; Degnan, P.H.; Hu, J.; Peter, I.; Zhang, W.; et al. Immunoglobulin A coating identifies colitogenic bacteria in inflammatory bowel disease. Cell 2014, 158, 1000–1010. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.Q.; Huang, H.L.; Peng, W.; Liu, Y.D.; Zhou, Y.L.; Xu, H.M.; Zhang, L.J.; Zhao, C.; Nie, Y.Q. Altered Pattern of Immunoglobulin A-Targeted Microbiota in Inflammatory Bowel Disease After Fecal Transplantation. Front. Microbiol. 2022, 13, 873018. [Google Scholar] [CrossRef]
- Michaud, E.; Waeckel, L.; Gayet, R.; Goguyer-Deschaumes, R.; Chanut, B.; Jospin, F.; Bathany, K.; Monnoye, M.; Genet, C.; Prier, A.; et al. Alteration of microbiota antibody-mediated immune selection contributes to dysbiosis in inflammatory bowel diseases. EMBO Mol. Med. 2022, 14, e15386. [Google Scholar] [CrossRef]
- Shapiro, J.M.; de Zoete, M.R.; Palm, N.W.; Laenen, Y.; Bright, R.; Mallette, M.; Bu, K.; Bielecka, A.A.; Xu, F.; Hurtado-Lorenzo, A.; et al. Immunoglobulin A Targets a Unique Subset of the Microbiota in Inflammatory Bowel Disease. Cell Host Microbe 2021, 29, 83–93.e3. [Google Scholar] [CrossRef]
- Bourgonje, A.R.; Roo-Brand, G.; Lisotto, P.; Sadaghian Sadabad, M.; Reitsema, R.D.; de Goffau, M.C.; Faber, K.N.; Dijkstra, G.; Harmsen, H.J.M. Patients With Inflammatory Bowel Disease Show IgG Immune Responses Towards Specific Intestinal Bacterial Genera. Front. Immunol. 2022, 13, 842911. [Google Scholar] [CrossRef]
- De Palma, G.; Nadal, I.; Medina, M.; Donat, E.; Ribes-Koninckx, C.; Calabuig, M.; Sanz, Y. Intestinal dysbiosis and reduced immunoglobulin-coated bacteria associated with coeliac disease in children. BMC Microbiol. 2010, 10, 63. [Google Scholar] [CrossRef]
- Hsieh, C.S.; Rengarajan, S.; Kau, A.; Tarazona-Meza, C.; Nicholson, A.; Checkley, W.; Romero, K.; Hansel, N.N. Altered IgA Response to Gut Bacteria Is Associated with Childhood Asthma in Peru. J. Immunol. 2021, 207, 398–407. [Google Scholar] [CrossRef]
- Dzidic, M.; Abrahamsson, T.R.; Artacho, A.; Bjorksten, B.; Collado, M.C.; Mira, A.; Jenmalm, M.C. Aberrant IgA responses to the gut microbiota during infancy precede asthma and allergy development. J. Allergy Clin. Immunol. 2017, 139, 1017–1025.e1014. [Google Scholar] [CrossRef] [PubMed]
- Scheithauer, T.P.M.; Davids, M.; Winkelmeijer, M.; Verdoes, X.; Aydin, O.; de Brauw, M.; van de Laar, A.; Meijnikman, A.S.; Gerdes, V.E.A.; van Raalte, D.; et al. Compensatory intestinal antibody response against pro-inflammatory microbiota after bariatric surgery. Gut Microbes 2022, 14, 2031696. [Google Scholar] [CrossRef] [PubMed]
- Sterlin, D.; Larsen, M.; Fadlallah, J.; Parizot, C.; Vignes, M.; Autaa, G.; Dorgham, K.; Juste, C.; Lepage, P.; Aboab, J.; et al. Perturbed Microbiota/Immune Homeostasis in Multiple Sclerosis. Neurol.-NeuroImmunol. Neuroinflamm. 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Goedert, J.J.; Hua, X.; Bielecka, A.; Okayasu, I.; Milne, G.L.; Jones, G.S.; Fujiwara, M.; Sinha, R.; Wan, Y.; Xu, X.; et al. Postmenopausal breast cancer and oestrogen associations with the IgA-coated and IgA-noncoated faecal microbiota. Br. J. Cancer 2018, 118, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Viladomiu, M.; Kivolowitz, C.; Abdulhamid, A.; Dogan, B.; Victorio, D.; Castellanos, J.G.; Woo, V.; Teng, F.; Tran, N.L.; Sczesnak, A.; et al. IgA-coated E. coli enriched in Crohn’s disease spondyloarthritis promote TH17-dependent inflammation. Sci. Transl. Med. 2017, 9, eaaf9655. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, A.; Liu, Z.; Joelsson, A.; Zhu, J. Vibrio cholerae virulence regulator-coordinated evasion of host immunity. Proc. Natl. Acad. Sci. USA 2006, 103, 14542–14547. [Google Scholar] [CrossRef] [PubMed]
- Shanker, S.; Czako, R.; Sapparapu, G.; Alvarado, G.; Viskovska, M.; Sankaran, B.; Atmar, R.L.; Crowe, J.E., Jr.; Estes, M.K.; Prasad, B.V. Structural basis for norovirus neutralization by an HBGA blocking human IgA antibody. Proc. Natl. Acad. Sci. USA 2016, 113, E5830–E5837. [Google Scholar] [CrossRef]
- Tokuhara, D.; Yuki, Y.; Nochi, T.; Kodama, T.; Mejima, M.; Kurokawa, S.; Takahashi, Y.; Nanno, M.; Nakanishi, U.; Takaiwa, F.; et al. Secretory IgA-mediated protection against V. cholerae and heat-labile enterotoxin-producing enterotoxigenic Escherichia coli by rice-based vaccine. Proc. Natl. Acad. Sci. USA 2010, 107, 8794–8799. [Google Scholar] [CrossRef]
- Kau, A.L.; Planer, J.D.; Liu, J.; Rao, S.; Yatsunenko, T.; Trehan, I.; Manary, M.J.; Liu, T.C.; Stappenbeck, T.S.; Maleta, K.M.; et al. Functional characterization of IgA-targeted bacterial taxa from undernourished Malawian children that produce diet-dependent enteropathy. Sci. Transl. Med. 2015, 7, 276ra224. [Google Scholar] [CrossRef]
- Huus, K.E.; Rodriguez-Pozo, A.; Kapel, N.; Nestoret, A.; Habib, A.; Dede, M.; Manges, A.; Collard, J.M.; Sansonetti, P.J.; Vonaesch, P.; et al. Immunoglobulin recognition of fecal bacteria in stunted and non-stunted children: Findings from the Afribiota study. Microbiome 2020, 8, 113. [Google Scholar] [CrossRef]
- Huus, K.E.; Bauer, K.C.; Brown, E.M.; Bozorgmehr, T.; Woodward, S.E.; Serapio-Palacios, A.; Boutin, R.C.T.; Petersen, C.; Finlay, B.B. Commensal Bacteria Modulate Immunoglobulin A Binding in Response to Host Nutrition. Cell Host Microbe 2020, 27, 909–921.e905. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.L.; Essigmann, H.T.; Hoffman, K.L.; Palm, N.W.; Gunter, S.M.; Sederstrom, J.M.; Petrosino, J.F.; Jun, G.; Aguilar, D.; Perkison, W.B.; et al. Impact of Diabetes on the Gut and Salivary IgA Microbiomes. Infect. Immun 2020, 88, e00301-20. [Google Scholar] [CrossRef] [PubMed]
- Pearson, J.A.; Ding, H.; Hu, C.; Peng, J.; Galuppo, B.; Wong, F.S.; Caprio, S.; Santoro, N.; Wen, L. IgM-associated gut bacteria in obesity and type 2 diabetes in C57BL/6 mice and humans. Diabetologia 2022, 65, 1398–1411. [Google Scholar] [CrossRef] [PubMed]
- Dzunkova, M.; Moya, A.; Vazquez-Castellanos, J.F.; Artacho, A.; Chen, X.; Kelly, C.; D’Auria, G. Active and Secretory IgA-Coated Bacterial Fractions Elucidate Dysbiosis in Clostridium difficile Infection. mSphere 2016, 1, e00101-16. [Google Scholar] [CrossRef]
- Huus, K.E.; Frankowski, M.; Pucic-Bakovic, M.; Vuckovic, F.; Lauc, G.; Mullish, B.H.; Marchesi, J.R.; Monaghan, T.M.; Kao, D.; Finlay, B.B. Changes in IgA-targeted microbiota following fecal transplantation for recurrent Clostridioides difficile infection. Gut Microbes 2021, 13, 1–12. [Google Scholar] [CrossRef]
- Ng, Q.X.; Soh, A.Y.S.; Loke, W.; Lim, D.Y.; Yeo, W.S. The role of inflammation in irritable bowel syndrome (IBS). J. Inflamm. Res. 2018, 11, 345–349. [Google Scholar] [CrossRef]
- Liu, Y.; Yuan, X.; Li, L.; Lin, L.; Zuo, X.; Cong, Y.; Li, Y. Increased Ileal Immunoglobulin A Production and Immunoglobulin A-Coated Bacteria in Diarrhea-Predominant Irritable Bowel Syndrome. Clin. Transl. Gastroenterol. 2020, 11, e00146. [Google Scholar] [CrossRef]
- Rengarajan, S.; Knoop, K.A.; Rengarajan, A.; Chai, J.N.; Grajales-Reyes, J.G.; Samineni, V.K.; Russler-Germain, E.V.; Ranganathan, P.; Fasano, A.; Sayuk, G.S.; et al. A Potential Role for Stress-Induced Microbial Alterations in IgA-Associated Irritable Bowel Syndrome with Diarrhea. Cell Rep. Med. 2020, 1, 100124. [Google Scholar] [CrossRef]
- Probstel, A.K.; Zhou, X.; Baumann, R.; Wischnewski, S.; Kutza, M.; Rojas, O.L.; Sellrie, K.; Bischof, A.; Kim, K.; Ramesh, A.; et al. Gut microbiota-specific IgA(+) B cells traffic to the CNS in active multiple sclerosis. Sci. Immunol. 2020, 5, eabc7191. [Google Scholar] [CrossRef]
- Rojas, O.L.; Probstel, A.K.; Porfilio, E.A.; Wang, A.A.; Charabati, M.; Sun, T.; Lee, D.S.W.; Galicia, G.; Ramaglia, V.; Ward, L.A.; et al. Recirculating Intestinal IgA-Producing Cells Regulate Neuroinflammation via IL-10. Cell 2019, 177, 492–493. [Google Scholar] [CrossRef]
- Chu, N.D.; Crothers, J.W.; Nguyen, L.T.T.; Kearney, S.M.; Smith, M.B.; Kassam, Z.; Collins, C.; Xavier, R.; Moses, P.L.; Alm, E.J. Dynamic Colonization of Microbes and Their Functions after Fecal Microbiota Transplantation for Inflammatory Bowel Disease. mBio 2021, 12, e0097521. [Google Scholar] [CrossRef] [PubMed]
- Lima, S.F.; Gogokhia, L.; Viladomiu, M.; Chou, L.; Putzel, G.; Jin, W.B.; Pires, S.; Guo, C.J.; Gerardin, Y.; Crawford, C.V.; et al. Transferable Immunoglobulin A-Coated Odoribacter splanchnicus in Responders to Fecal Microbiota Transplantation for Ulcerative Colitis Limits Colonic Inflammation. Gastroenterology 2022, 162, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Basu, S.; Bal, V.; Rath, S.; George, A. Gut IgA abundance in adult life is a major determinant of resistance to dextran sodium sulfate-colitis and can compensate for the effects of inadequate maternal IgA received by neonates. Immunology 2019, 158, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Fretheim, H.; Chung, B.K.; Didriksen, H.; Baekkevold, E.S.; Midtvedt, O.; Brunborg, C.; Holm, K.; Valeur, J.; Tennoe, A.H.; Garen, T.; et al. Fecal microbiota transplantation in systemic sclerosis: A double-blind, placebo-controlled randomized pilot trial. PLoS ONE 2020, 15, e0232739. [Google Scholar] [CrossRef] [PubMed]
- Stebegg, M.; Silva-Cayetano, A.; Innocentin, S.; Jenkins, T.P.; Cantacessi, C.; Gilbert, C.; Linterman, M.A. Heterochronic faecal transplantation boosts gut germinal centres in aged mice. Nat. Commun. 2019, 10, 2443. [Google Scholar] [CrossRef]
- Xiong, N.; Hu, S. Regulation of intestinal IgA responses. Cell Mol. Life Sci. 2015, 72, 2645–2655. [Google Scholar] [CrossRef]
- Tezuka, H.; Ohteki, T. Regulation of IgA Production by Intestinal Dendritic Cells and Related Cells. Front. Immunol. 2019, 10, 1891. [Google Scholar] [CrossRef]
- Cong, Y.; Feng, T.; Fujihashi, K.; Schoeb, T.R.; Elson, C.O. A dominant, coordinated T regulatory cell-IgA response to the intestinal microbiota. Proc. Natl. Acad. Sci. USA 2009, 106, 19256–19261. [Google Scholar] [CrossRef]
- Wang, W.; Liu, Z.; Yue, W.; Zhu, L.; Zhong, H.; Yang, C.; He, T.; Wan, P.; Geng, J. Mucosa-Colonizing Microbiota Correlate With Host Autophagy Signaling in Patients With Inflammatory Bowel Disease. Front. Microbiol. 2022, 13, 875238. [Google Scholar] [CrossRef]
- Miyauchi, E.; Taida, T.; Kawasumi, M.; Ohkusa, T.; Sato, N.; Ohno, H. Analysis of colonic mucosa-associated microbiota using endoscopically collected lavage. Sci. Rep. 2022, 12, 1758. [Google Scholar] [CrossRef]
- Strobbe, F.; Benard, M.V.; Rossen, N.G.; de Vos, W.M.; Kumar, N.; Lawley, T.D.; Zoetendal, E.G.; Hugenholtz, F.; Ponsioen, C.Y. A novel technique capable of taking ‘protected’ biopsies for reliable assessment of the distribution of microbiota along the colonic mucosa. J. Microbiol. Methods 2021, 185, 106204. [Google Scholar] [CrossRef] [PubMed]
- Jalanka, J.; Mattila, E.; Jouhten, H.; Hartman, J.; de Vos, W.M.; Arkkila, P.; Satokari, R. Long-term effects on luminal and mucosal microbiota and commonly acquired taxa in faecal microbiota transplantation for recurrent Clostridium difficile infection. BMC Med. 2016, 14, 155. [Google Scholar] [CrossRef] [PubMed]
- Zuppi, M.; Hendrickson, H.L.; O’Sullivan, J.M.; Vatanen, T. Phages in the Gut Ecosystem. Front. Cell. Infect. Microbiol. 2021, 11, 822562. [Google Scholar] [CrossRef] [PubMed]
- Sausset, R.; Petit, M.A.; Gaboriau-Routhiau, V.; De Paepe, M. New insights into intestinal phages. Mucosal Immunol. 2020, 13, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Barr, J.J.; Auro, R.; Furlan, M.; Whiteson, K.L.; Erb, M.L.; Pogliano, J.; Stotland, A.; Wolkowicz, R.; Cutting, A.S.; Doran, K.S.; et al. Bacteriophage adhering to mucus provide a non-host-derived immunity. Proc. Natl. Acad. Sci. USA 2013, 110, 10771–10776. [Google Scholar] [CrossRef]
- Majewska, J.; Beta, W.; Lecion, D.; Hodyra-Stefaniak, K.; Klopot, A.; Kazmierczak, Z.; Miernikiewicz, P.; Piotrowicz, A.; Ciekot, J.; Owczarek, B.; et al. Oral Application of T4 Phage Induces Weak Antibody Production in the Gut and in the Blood. Viruses 2015, 7, 4783–4799. [Google Scholar] [CrossRef]
- Majewska, J.; Kazmierczak, Z.; Lahutta, K.; Lecion, D.; Szymczak, A.; Miernikiewicz, P.; Drapala, J.; Harhala, M.; Marek-Bukowiec, K.; Jedruchniewicz, N.; et al. Induction of Phage-Specific Antibodies by Two Therapeutic Staphylococcal Bacteriophages Administered per os. Front. Immunol. 2019, 10, 2607. [Google Scholar] [CrossRef]
- Ott, S.J.; Waetzig, G.H.; Rehman, A.; Moltzau-Anderson, J.; Bharti, R.; Grasis, J.A.; Cassidy, L.; Tholey, A.; Fickenscher, H.; Seegert, D.; et al. Efficacy of Sterile Fecal Filtrate Transfer for Treating Patients With Clostridium difficile Infection. Gastroenterology 2017, 152, 799–811.e797. [Google Scholar] [CrossRef]
- Wu, X.; Xia, Y.; He, F.; Zhu, C.; Ren, W. Intestinal mycobiota in health and diseases: From a disrupted equilibrium to clinical opportunities. Microbiome 2021, 9, 60. [Google Scholar] [CrossRef]
- Doron, I.; Mesko, M.; Li, X.V.; Kusakabe, T.; Leonardi, I.; Shaw, D.G.; Fiers, W.D.; Lin, W.Y.; Bialt-DeCelie, M.; Roman, E.; et al. Mycobiota-induced IgA antibodies regulate fungal commensalism in the gut and are dysregulated in Crohn’s disease. Nat. Microbiol. 2021, 6, 1493–1504. [Google Scholar] [CrossRef]
- Jovel, J.; Patterson, J.; Wang, W.; Hotte, N.; O’Keefe, S.; Mitchel, T.; Perry, T.; Kao, D.; Mason, A.L.; Madsen, K.L.; et al. Characterization of the Gut Microbiome Using 16S or Shotgun Metagenomics. Front. Microbiol. 2016, 7, 459. [Google Scholar] [CrossRef] [PubMed]


| (A) | ||||
|---|---|---|---|---|
| Phylum | Order | Family | Genus | Species |
| Firmicutes (Bacillota) | ||||
| Clostrdiales | Lachnospiraceae | Roseburia | Roseburia spp. | |
| Eubacteriales | Clostridiaceae | Clostridium | Clostridium spp. | |
| Eubacteriales | Oscillospiraceae | Ruminococcus | Ruminococcus spp. | |
| Eubacteriales | Oscillospiraceae | Faecalibacterium | Faecalibacterium prausnitzii | |
| Eubacteriales | Lachnospiraceae | Blautia | Blautia spp. | |
| Eubacteriales | Lachnospiraceae | Coprococcus | Coprococcus spp. | |
| Eubacteriales | Lachnospiraceae | Anaerostipes | Anaerostipes spp. | |
| Lactobacillales | Streptococcaceae | Streptococcus | Streptococcus spp. | |
| Vellionellales | Veillonellaceae | Veillonella | Veillonella spp. | |
| Vellionellales | Veillonellaceae | Dialister | Dialister spp. | |
| Erysipelotrichia | Turicibacteraceae | Turicibacter | Turicibacter spp. | |
| Bacteroidetes (Bacteroidota) | ||||
| Bacteroidales | Bacteroidaceae | Bacteroides | Bacteroides ovatus | |
| Bacteroidales | Rikenellaceae | Too many to list | Too many to list | |
| Proteobacteria (Pseudomonadota) | ||||
| Enterobacterales | Enterobacteriaceae | Too many to list | E. coli, Klebsiella, etc. | |
| Pasteurellales | Pasteurellaceae | Hemophilus | Hemophilus spp. | |
| Actinobacteria (Actinomycetota) | ||||
| Coriobacteriales | Coriobacteriaceae | Collinsella | Collinsella spp. | |
| Bifidobacteriales | Bifidobacteriaceae | Bifidobacterium | Bifidobacterium spp. | |
| Eggerthellales | Eggerthellaceae | Eggerthella | Eggerthella spp. | |
| (B) | ||||
| Phylum | Order | Family | Genus | Species |
| Firmicutes (Bacillota) | ||||
| Clostrdiales | Lachnospiraceae | Roseburia | Roseburia spp. | |
| Lactobacillales | Enterococcaceae | Enterococcus | Enterococcus spp. | |
| Eubacteriales | Clostridiaceae | Clostridium | Clostridium spp. | |
| Eubacteriales | Oscillospiraceae | Ruminococcus | Ruminococcus spp. | |
| Eubacteriales | Oscillospiraceae | Faecalibacterium | Faecalibacterium prausnitzii | |
| Eubacteriales | Lachnospiraceae | Blautia | Blautia spp. | |
| Eubacteriales | Lachnospiraceae | Coprococcus | Coprococcus spp. | |
| Eubacteriales | Lachnospiraceae | Anaerostipes | Anaerostipes spp. | |
| Clostridiales | Eubacteriaceae | Eubacterium | Eubacterium spp. | |
| Erysipeiotrichia | Erysipelotrichaceae | Too many to list | Too many to list | |
| Lactobacillales | Lactobacillaceae | Lactobacillus | Lactobacillus spp. | |
| Lactobacillales | Streptococcaceae | Streptococcus | Streptococcus spp. | |
| Bacteroidetes (Bacteroidota) | ||||
| Bacteroidales | Bacteroidaceae | Bacteroides | Bacteroides spp. | |
| Bacteroidales | Prevotellaceae | Prevotella | Prevotella spp. | |
| Verrucomicrobia (Verrucomicrobiota) | ||||
| Verrucomicrobiales | Akkermansiaceae | Akkermansia | Akkermansia muciniphila | |
| Proteobacteria (Pseudomonadota) | ||||
| Pseudomonadales | Pseudomonadaceae | Pseudomonas | Pseudomonas spp. | |
| Enterobacterales | Enterobacteriaceae | 53 different genera | E. coli, Klebsiella, etc. | |
| Burkholderiales | Alcaligenaceae | Too many to list | Too many to list | |
| Actinobacteria (Actinomycetota) | ||||
| Coriobacteriales | Coriobacteriaceae | Collinsella | Collinsella spp. | |
| Bifidobacteriales | Bifidobacteriaceae | Bifidobacterium | Bifidobacterium spp. | |
| Eggerthellales | Eggerthellaceae | Eggerthella | Eggerthella spp. | |
| Propionibacteriales | Nocardioidaceae | Aeromicrobium, Marmoricola, Mumia and Nocardioides | More than 50 species | |
| Condition | General Findings | Comment |
|---|---|---|
| Inflammatory Bowel Disease | Ig-coated bacteria were increased in IBD [32,33,34,35,36,37] and correlated directly with inflammation markers and severity of disease and facilitated development of colitis in mice [34]. The diversity of the Ig-coated population was lower than the core Ig-negative microbial population [38]. Treatment with TNF-alpha was associated with a change in pattern of IgA-coated bacteria that predicted response to treatment [37]. | The enhanced production of Ig appears to be a direct result of pathogenic strains of bacteria producing inflammation that are involved in disease pathogenesis. Changes in coated IgA-bacteria may provide predictive value for therapeutic response in IBD. |
| Enteric Infections | IgA binds to viral, bacterial and parasitic pathogens and influences growth and virulence of the strains [46,47,48]. | This is an example of “immune exclusion”. |
| Celiac Disease in Children | IgA-, IgG- and IgM-coated bacterial levels were low in childhood celiac disease, treated or untreated [39]. | Gluten-free microbiome friendly diet may improve the microbiome in these children. |
| Childhood Allergies and Asthma | IgA-coated fecal bacteria were reduced in asthma with the level of reduction corelating with more severe disease [40,41]. | Low Ig-coated bacteria impairs gut health, and a microbiome friendly diet should be evaluated in these children. |
| Undernutrition | IgA-coated fecal bacteria were reduced [51] unless infected by enteric pathogens or colonized by pro-inflammatory strains of Enterobacteriaceae [49,50] | Low Ig-coated bacteria impairs gut health, protein caloric intake and microbiome friendly diets are needed. |
| Obesity and type 2 diabetes | In a mouse model of obesity, IgM-coated bacteria appeared to be involved in the immunopathogenesis of obesity and type 2 diabetes [53]. In humans with diabetes, IgA commonly coats strains of proinflammatory Enterobacteriaceae that appear to contribute to regulation of obesity-related insulin resistance [42]. | The microbiome and Ig-coated bacteria need more study in obesity and diabetes with potential improvement in microbiome diversity with FMT. |
| C difficile Infection | The total IgA-microbiome was largely depleted in this infection, with proinflammatory strains of IgA-coated Enterobacteriaceae dominating the microbiome [55]. | Antibiotic effects on normally protective microbiota are the driving force leading to dysbiosis and susceptibility to CDI. |
| Irritable Bowel Syndrome | The proportion of IgA-coated bacteria was elevated in IBS-D, was associated with emergence of proinflammatory taxa [57], and in an animal model enhanced enteric symptoms and potentiated bacterial translocation [58]. | Patients with IBS-D show increased proportions of IgA-coated bacteria that may be involved with the pathogenesis of the disease and may represent therapeutic targets. |
| Multiple Sclerosis | IgA-coated bacteria were reduced in multiple sclerosis [43,60]. | Defective IgA responses in MS need further study and the authors suggest that microbiota reconstitution with fecal microbiota transplantation should be evaluated in MS to look for clinical and biologic effects. |
| Breast Cancer | IgA-coated bacteria were reduced in women with breast cancer [44] and the IgA-microbiome appeared unique, with increased proportion of IgA-coated Ruminococcus oscilibacter (p = 0.003). The microbiome changes seen correlated with urinary estrogen levels. | More studies are needed of the Ig-biome in cancer. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
DuPont, H.L.; Jiang, Z.-D.; Alexander, A.S.; DuPont, A.W.; Brown, E.L. Intestinal IgA-Coated Bacteria in Healthy- and Altered-Microbiomes (Dysbiosis) and Predictive Value in Successful Fecal Microbiota Transplantation. Microorganisms 2023, 11, 93. https://doi.org/10.3390/microorganisms11010093
DuPont HL, Jiang Z-D, Alexander AS, DuPont AW, Brown EL. Intestinal IgA-Coated Bacteria in Healthy- and Altered-Microbiomes (Dysbiosis) and Predictive Value in Successful Fecal Microbiota Transplantation. Microorganisms. 2023; 11(1):93. https://doi.org/10.3390/microorganisms11010093
Chicago/Turabian StyleDuPont, Herbert L., Zhi-Dong Jiang, Ashley S. Alexander, Andrew W. DuPont, and Eric L. Brown. 2023. "Intestinal IgA-Coated Bacteria in Healthy- and Altered-Microbiomes (Dysbiosis) and Predictive Value in Successful Fecal Microbiota Transplantation" Microorganisms 11, no. 1: 93. https://doi.org/10.3390/microorganisms11010093
APA StyleDuPont, H. L., Jiang, Z.-D., Alexander, A. S., DuPont, A. W., & Brown, E. L. (2023). Intestinal IgA-Coated Bacteria in Healthy- and Altered-Microbiomes (Dysbiosis) and Predictive Value in Successful Fecal Microbiota Transplantation. Microorganisms, 11(1), 93. https://doi.org/10.3390/microorganisms11010093