
Gut Microbial Characterization of Melon-Headed Whales (Peponocephala electra) Stranded in China




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Extraction and Sequencing
2.3. Microbial Community Analysis
3. Results
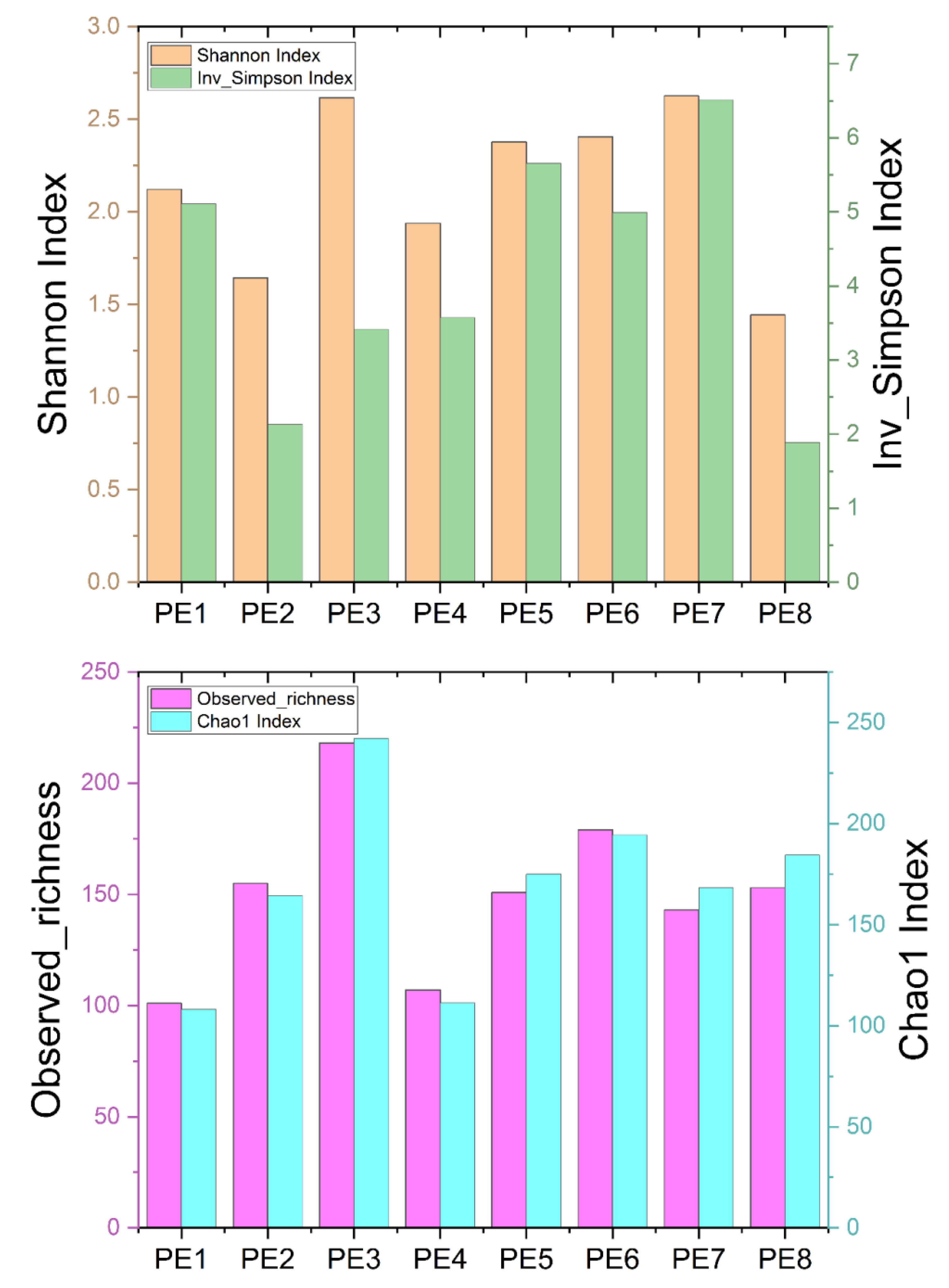
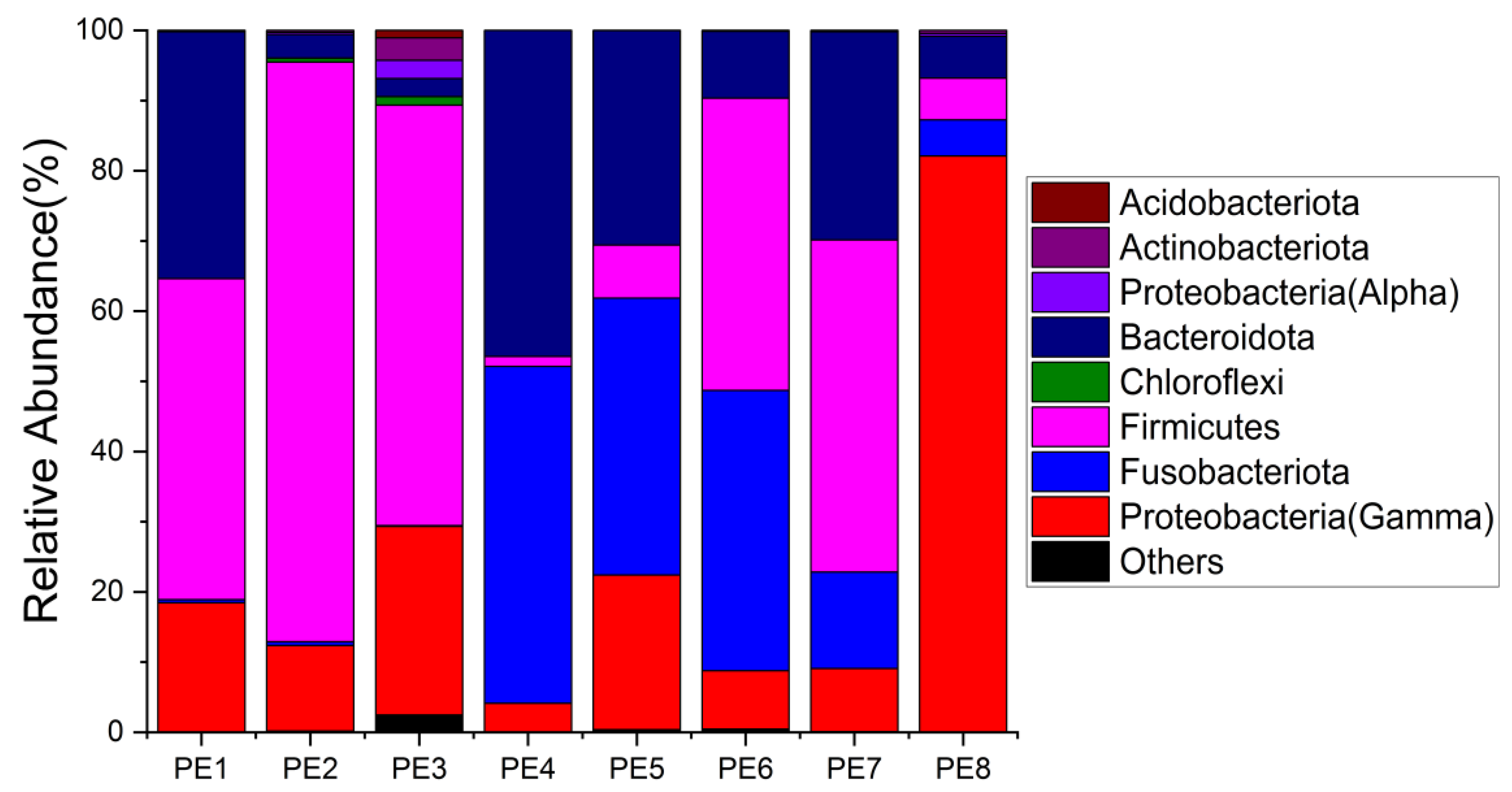
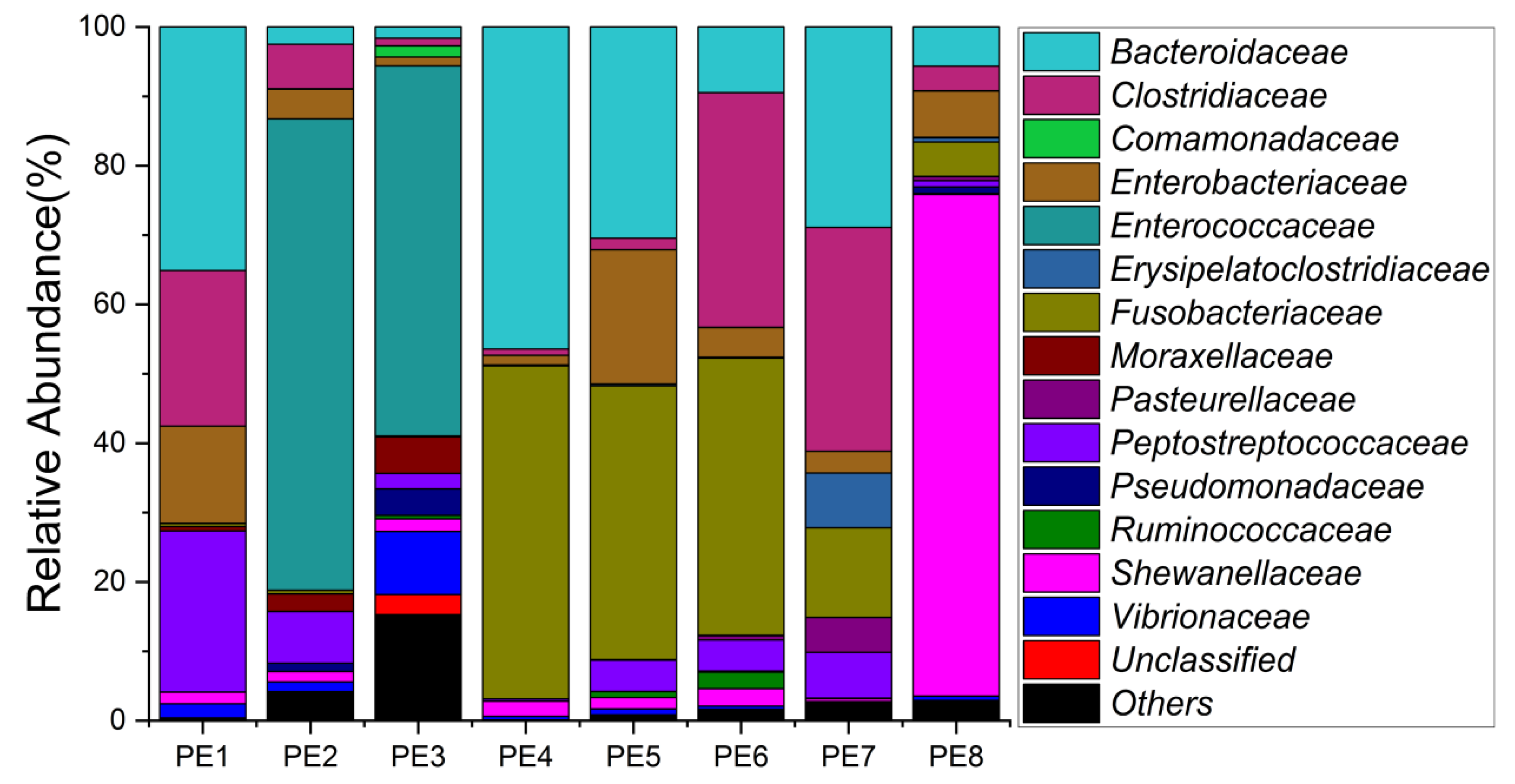
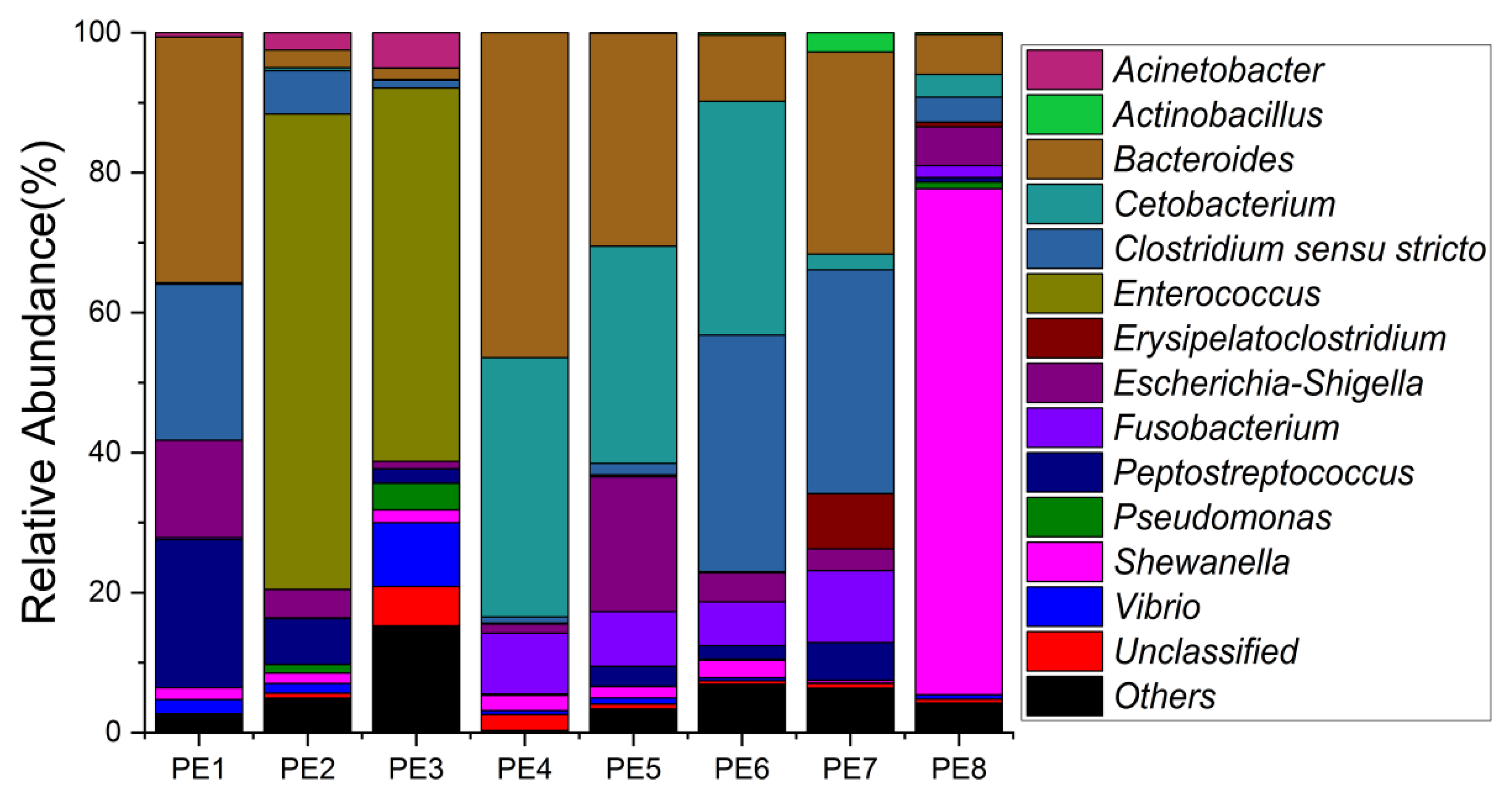
3.1. Sequencing Statistics and Microbial Diversity
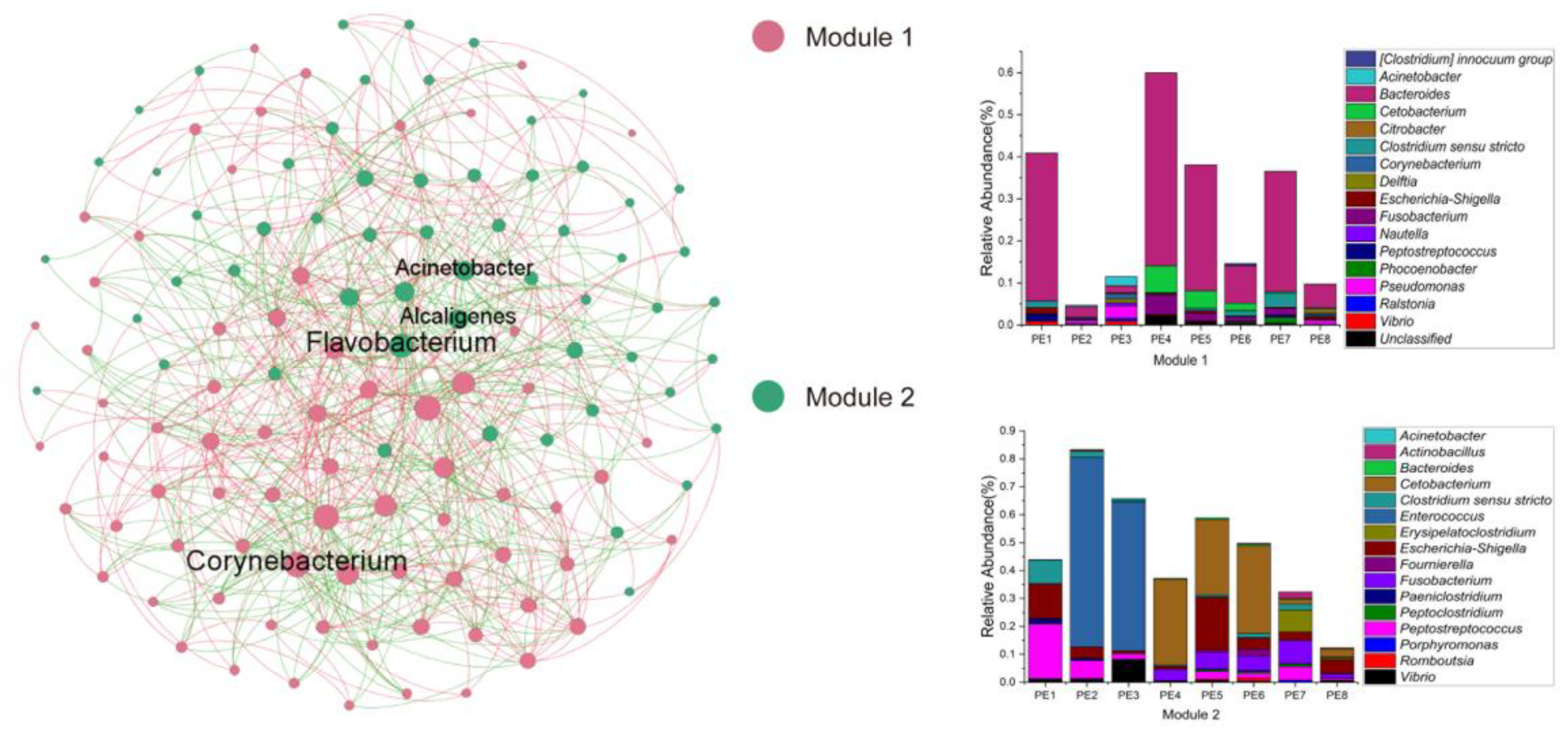
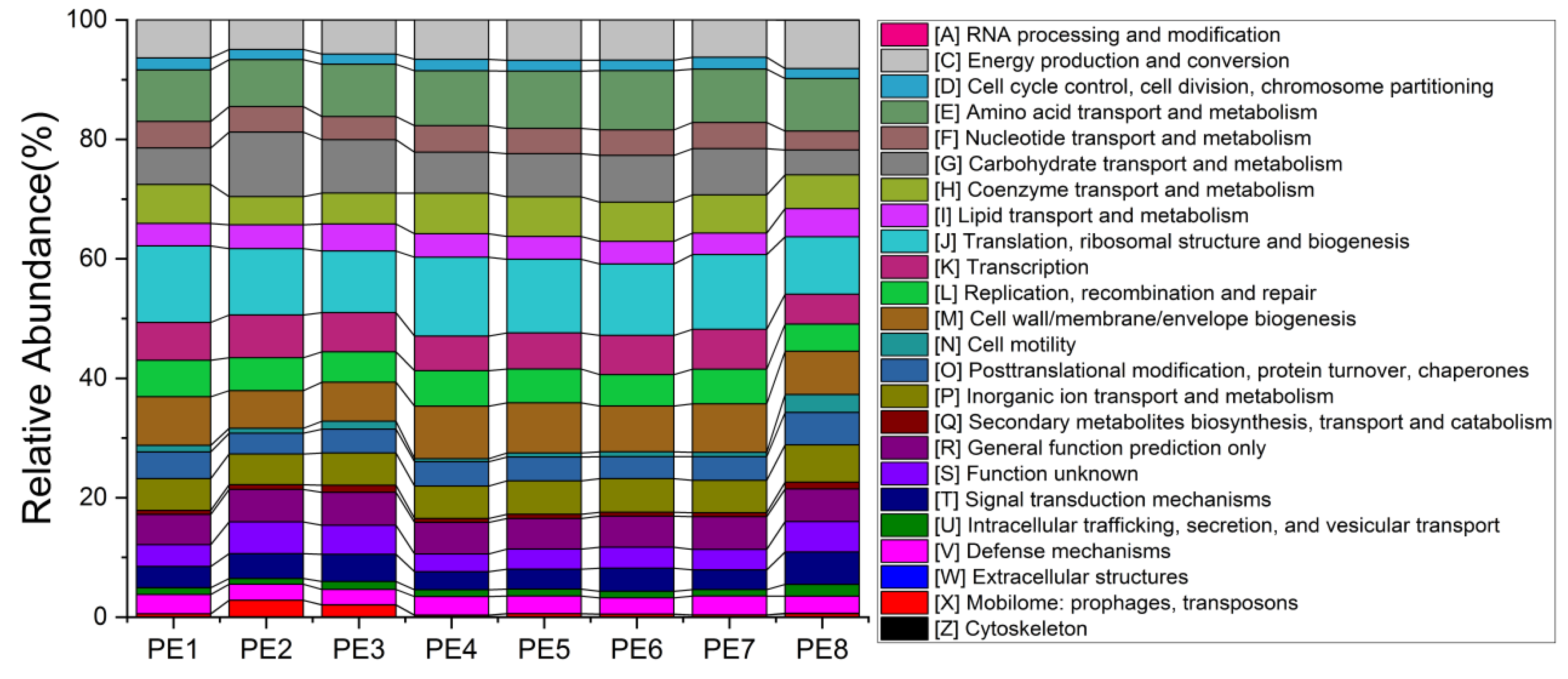
3.2. Co-Occurrence Network and Functional Profile of Gut Microbial Communities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vilstrup, J.T.; Ho, S.Y.; Foote, A.D.; A Morin, P.; Kreb, D.; Krützen, M.; Parra, G.J.; Robertson, K.M.; De Stephanis, R.; Verborgh, P.; et al. Mitogenomic phylogenetic analyses of the Delphinidae with an emphasis on the Globicephalinae. BMC Evol. Biol. 2011, 11, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perryman, W.L.; Danil, K. Melon-Headed Whale. In Encyclopedia of Marine Mammals, 3rd ed.; Wursig, B., Thewissen, J.G.M., Kovacs, K., Eds.; Academy Press: Cambridge, MA, USA, 2018; pp. 593–595. [Google Scholar] [CrossRef]
- Jefferson, T.A.; Webber, M.A.; Pitman, R. Marine Mammals of the World: A Comprehensive Guide to their Identification, 2nd ed.; Academic Press: San Diego, CA, 2015. [Google Scholar]
- Jefferson, T.A.; Barros, N.B. Peponocephala electra. Mammalian Species 1997, 553, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Spitz, J.; Cherel, Y.; Bertin, S.; Kiszka, J.; Dewez, A.; Ridoux, V. Prey preferences among the community of deep-diving odontocetes from the Bay of Biscay, Northeast Atlantic. Deep Sea Res. Part I Oceanogr. Res. Pap. 2011, 58, 273–282. [Google Scholar] [CrossRef]
- Gensollen, T.; Iyer, S.S.; Kasper, D.L.; Blumberg, R.S. How colonization by microbiota in early life shapes the immune system. Science 2016, 352, 539–544. [Google Scholar] [CrossRef] [Green Version]
- Dang, A.T.; Marsland, B.J. Microbes, metabolites, and the gut–lung axis. Mucosal Immunol. 2019, 12, 843–850. [Google Scholar] [CrossRef] [Green Version]
- Krajmalnik-Brown, R.; Ilhan, Z.-E.; Kang, D.-W.; DiBaise, J.K. Effects of Gut Microbes on Nutrient Absorption and Energy Regulation. Nutr. Clin. Pr. 2012, 27, 201–214. [Google Scholar] [CrossRef] [Green Version]
- Woting, A.; Blaut, M. The Intestinal Microbiota in Metabolic Disease. Nutrients 2016, 8, 202. [Google Scholar] [CrossRef]
- Dzutsev, A.; Badger, J.H.; Perez-Chanona, E.; Roy, S.; Salcedo, R.; Smith, C.K.; Trinchieri, G. Microbes and Cancer. Annu. Rev. Immunol. 2017, 35, 199–228. [Google Scholar] [CrossRef]
- Quin, C.; Gibson, D.L. Human behavior, not race or geography, is the strongest predictor of microbial succession in the gut bacteriome of infants. Gut Microbes 2020, 11, 1143–1171. [Google Scholar] [CrossRef] [Green Version]
- Sanders, J.G.; Beichman, A.C.; Roman, J.; Scott, J.; Emerson, D.; McCarthy, J.J.; Girguis, P.R. Baleen whales host a unique gut microbiome with similarities to both carnivores and herbivores. Nat. Commun. 2015, 6, 8285. [Google Scholar] [CrossRef]
- Bai, S.; Zhang, P.; Zhang, C.; Du, J.; Du, X.; Zhu, C.; Liu, J.; Xie, P.; Li, S. Comparative Study of the Gut Microbiota Among Four Different Marine Mammals in an Aquarium. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Segawa, T.; Sawa, S.; Nishitani, C.; Ueda, K.; Itou, T.; Asahina, K.; Suzuki, M. Comparison of the gut microbiota of captive common bottlenose dolphins Tursiops truncatus in three aquaria. J. Appl. Microbiol. 2018, 126, 31–39. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, R.W.; Chen, M.; Zheng, J.; Zhao, Q.; Wang, D. Analysis of the bacterial diversity in the fecal material of the endangered Yangtze finless porpoise, Neophocaena phocaenoides asiaeorientalis. Mol. Biol. Rep. 2011, 39, 5669–5676. [Google Scholar] [CrossRef]
- Bai, S.; Zhang, P.; Lin, M.; Lin, W.; Yang, Z.; Li, S. Microbial diversity and structure in the gastrointestinal tracts of two stranded short-finned pilot whales (Globicephala macrorhynchus) and a pygmy sperm whale (Kogia breviceps). Integr. Zool. 2021, 16, 324–335. [Google Scholar] [CrossRef]
- Tian, J.; Du, J.; Lu, Z.; Han, J.; Wang, Z.; Li, D.; Guan, X.; Wang, Z. Distribution of microbiota across different intestinal tract segments of a stranded dwarf minke whale, Balaenoptera acutorostrata. MicrobiologyOpen 2020, 9. [Google Scholar] [CrossRef]
- Wan, X.; Li, J.; Cheng, Z.; Ao, M.; Tian, R.; Mclaughlin, R.W.; Zheng, J.; Wang, D. The intestinal microbiome of an Indo-Pacific humpback dolphin (Sousa chinensis) stranded near the Pearl River Estuary, China. Integr. Zool. 2021, 16, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.-L.; McLaughlin, R.W.; Zheng, J.-S.; Hao, Y.-J.; Fan, F.; Tian, R.-M.; Wang, D. Microbial communities in different regions of the gastrointestinal tract in East Asian finless porpoises (Neophocaena asiaeorientalis sunameri). Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walters, W.; Hyde, E.R.; Berg-Lyons, D.; Ackermann, G.; Humphrey, G.; Parada, A.; Gilbert, J.A.; Jansson, J.K.; Caporaso, J.G.; Fuhrman, J.; et al. Improved Bacterial 16S rRNA Gene (V4 and V4-5) and Fungal Internal Transcribed Spacer Marker Gene Primers for Microbial Community Surveys. mSystems 2016, 1, e00009-15. [Google Scholar] [CrossRef] [Green Version]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Kong, Y. Btrim: A fast, lightweight adapter and quality trimming program for next-generation sequencing technologies. Genomics 2011, 98, 152–153. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UNOISE2: Improved error-correction for Illumina 16S and ITS amplicon sequencing. BioRxiv 2016, 081257. Available online: https://www.biorxiv.org/content/10.1101/081257.abstract (accessed on 29 January 2022).
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Chao, A. Nonparametric-estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 29 January 2022).
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.; Jiang, Y.-H.; Yang, Y.; He, Z.; Luo, F.; Zhou, J. Molecular ecological network analyses. BMC Bioinform. 2012, 13, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.; Zhang, P.; Qin, Y.; Tu, Q.; Yang, Y.; He, Z.; Schadt, C.; Zhou, J. Network succession reveals the importance of competition in response to emulsified vegetable oil amendment for uranium bioremediation. Environ. Microbiol. 2015, 18, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Caspi, R.; Billington, R.; Keseler, I.M.; Kothari, A.; Krummenacker, M.; Midford, P.E.; Ong, W.K.; Paley, S.; Subhraveti, P.; Karp, P.D. The MetaCyc database of metabolic pathways and enzymes—A 2019 update. Nucleic Acids Res. 2020, 48, D445–D453. [Google Scholar] [CrossRef] [Green Version]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Soverini, M.; Quercia, S.; Biancani, B.; Furlati, S.; Turroni, S.; Biagi, E.; Consolandi, C.; Peano, C.; Severgnini, M.; Rampelli, S.; et al. The bottlenose dolphin (Tursiops truncatus) faecal microbiota. FEMS Microbiol. Ecol. 2016, 92, fiw055. [Google Scholar] [CrossRef] [Green Version]
- Marón, C.F.; Kohl, K.; Chirife, A.; Di Martino, M.; Fons, M.P.; Navarro, M.A.; Beingesser, J.; McAloose, D.; Uzal, F.A.; Dearing, M.D.; et al. Symbiotic microbes and potential pathogens in the intestine of dead southern right whale (Eubalaena australis) calves. Anaerobe 2019, 57, 107–114. [Google Scholar] [CrossRef]
- Hooper, L.V.; Midtvedt, T.; Gordon, J.I. How host-microbial interactions shape the nutrient environment of the mammalian intestine. Annu. Rev. Nutr. 2002, 22, 283–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wexler, H.M. Bacteroides: The Good, the Bad, and the Nitty-Gritty. Clin. Microbiol. Rev. 2007, 20, 593–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erwin, P.M.; Rhodes, R.G.; Kiser, K.B.; Keenan-Bateman, T.F.; McLellan, W.A.; Pabst, D.A. High diversity and unique composition of gut microbiomes in pygmy (Kogia breviceps) and dwarf (K. sima) sperm whales. Sci. Rep. 2017, 7, 7205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Z.; Sun, J.; Lv, A.; Sung, Y.; Shi, H.; Hu, X.; Xing, K. Isolation, identification and characterization of Shewanella algae from reared tongue sole, Cynoglossus semilaevis Günther. Aquaculture 2017, 468, 356–362. [Google Scholar] [CrossRef]
- Holt, H.M.; Gahrn-Hansen, B.; Bruun, B. Shewanella algae and Shewanella putrefaciens: Clinical and microbiological characteristics. Clin. Microbiol. Infect. 2005, 11, 347–352. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, N.N.N.; Nasir, N.M.; Sahrani, F.K.; Ahmad, A.; Sairi, F. Characterization of putative pathogenic Shewanella algae isolated from ballast water. Veter-World 2021, 14, 678–688. [Google Scholar] [CrossRef]
- Lemaire, O.N.; Méjean, V.; Iobbi-Nivol, C. The Shewanella genus: Ubiquitous organisms sustaining and preserving aquatic ecosystems. FEMS Microbiol. Rev. 2020, 44, 155–170. [Google Scholar] [CrossRef]
- Paccalin, M.; Grollier, G.; Le Moal, G.; Rayeh, F.; Camiade, C. Rupture of a Primary Aortic Aneurysm Infected with Shewanella Alga. Scand. J. Infect. Dis. 2001, 33, 774–775. [Google Scholar] [CrossRef]
- Shanmuganathan, M.; Goh, B.L.; Lim, C.; NorFadhlina, Z.; Fairol, I. Shewanella algae Peritonitis in Patients on Peritoneal Dialysis. Perit. Dial. Int. J. Int. Soc. Perit. Dial. 2016, 36, 574–575. [Google Scholar] [CrossRef]
- Yang, Y. Emerging Patterns of Microbial Functional Traits. Trends Microbiol. 2021, 29, 874–882. [Google Scholar] [CrossRef]
- Bernard, K. The Genus Corynebacterium and Other Medically Relevant Coryneform-Like Bacteria. J. Clin. Microbiol. 2012, 50, 3152–3158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joung, Y.; Kim, H.; Joh, K. Flavobacterium jumunjinense sp. nov., isolated from a lagoon, and emended descriptions of Flavobacterium cheniae, Flavobacterium dongtanense and Flavobacterium gelidilacus. Int. J. Syst. Evol. Microbiol. 2013, 63, 3937–3943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batt, C. Alcaligenes. In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C., Tortorello, M., Eds.; Academic Press: Cambridge, MA, USA, 2014; Volume 1, pp. 38–42. [Google Scholar] [CrossRef]
- Paterson, D.L.; Peleg, A.Y. Acinetobacter. In Antimicrobial Drug Resistance, Mayers, D. L, Eds.; Humana Press: Totowa, NJ, USA, 2009; Volume 2, pp. 819–823. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, S.; Zhang, P.; Zhang, X.; Yang, Z.; Li, S. Gut Microbial Characterization of Melon-Headed Whales (Peponocephala electra) Stranded in China. Microorganisms 2022, 10, 572. https://doi.org/10.3390/microorganisms10030572
Bai S, Zhang P, Zhang X, Yang Z, Li S. Gut Microbial Characterization of Melon-Headed Whales (Peponocephala electra) Stranded in China. Microorganisms. 2022; 10(3):572. https://doi.org/10.3390/microorganisms10030572
Chicago/Turabian StyleBai, Shijie, Peijun Zhang, Xianfeng Zhang, Zixin Yang, and Songhai Li. 2022. "Gut Microbial Characterization of Melon-Headed Whales (Peponocephala electra) Stranded in China" Microorganisms 10, no. 3: 572. https://doi.org/10.3390/microorganisms10030572
APA StyleBai, S., Zhang, P., Zhang, X., Yang, Z., & Li, S. (2022). Gut Microbial Characterization of Melon-Headed Whales (Peponocephala electra) Stranded in China. Microorganisms, 10(3), 572. https://doi.org/10.3390/microorganisms10030572