
Experimental Infection of Captive Red Foxes (Vulpes vulpes) with Mycobacterium bovis

 , ,
, ,  , , and
, , and
Abstract
1. Introduction
2. Material and Methods
2.1. Ethics Statement
2.2. Animals and Samples Collection
2.3. Mycobacterium bovis Strain Used for the Infection
2.4. Infection
2.5. IDEXX M. bovis Ab Test
2.6. MAPIA
2.7. Collection of Tissues Post-Mortem
2.8. Histopathology
2.9. Culture
2.10. Molecular Diagnosis
3. Results
3.1. Clinical Follow Up
3.2. Serological Monitoring
3.3. Infection of Tissues
3.4. Excretion Monitoring
3.5. Pathology and Histopathology
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Santos, N.; Richomme, C.; Nunes, T.; Vicente, J.; Alves, P.C.; de la Fuente, J.; Correia-Neves, M.; Boschiroli, M.-L.; Delahay, R.; Gortázar, C. Quantification of the Animal Tuberculosis Multi-Host Community Offers Insights for Control. Pathogens 2020, 9, 421. [Google Scholar] [CrossRef] [PubMed]
- Haydon, D.T.; Cleaveland, S.; Taylor, L.H.; Laurenson, M.K. Identifying Reservoirs of Infection: A Conceptual and Practical Challenge. Emerging Infect. Dis. 2002, 8, 1468–1473. [Google Scholar] [CrossRef]
- Delahay, R.J.; Smith, G.C.; Barlow, A.M.; Walker, N.; Harris, A.; Clifton-Hadley, R.S.; Cheeseman, C.L. Bovine Tuberculosis Infection in Wild Mammals in the South-West Region of England: A Survey of Prevalence and a Semi-Quantitative Assessment of the Relative Risks to Cattle. Vet. J. 2007, 173, 287–301. [Google Scholar] [CrossRef] [PubMed]
- Matos, A.C.; Figueira, L.; Martins, M.H.; Pinto, M.L.; Matos, M.; Coelho, A.C. New Insights into Mycobacterium Bovis Prevalence in Wild Mammals in Portugal. Transbound Emerg. Dis. 2016, 63, e313–e322. [Google Scholar] [CrossRef]
- Millán, J.; Jiménez, M.A.; Viota, M.; Candela, M.G.; Peña, L.; León-Vizcaíno, L. Disseminated Bovine Tuberculosis in a Wild Red Fox (Vulpes Vulpes) in Southern Spain. J. Wildl. Dis. 2008, 44, 701–706. [Google Scholar] [CrossRef]
- Michelet, L.; De Cruz, K.; Hénault, S.; Tambosco, J.; Richomme, C.; Réveillaud, É.; Gares, H.; Moyen, J.-L.; Boschiroli, M.L. Mycobacterium Bovis Infection of Red Fox, France. Emerging Infect. Dis. 2018, 24, 1150–1153. [Google Scholar] [CrossRef] [PubMed]
- Richomme, C.; Réveillaud, E.; Moyen, J.-L.; Sabatier, P.; de Cruz, K.; Michelet, L.; Boschiroli, M.L. Mycobacterium Bovis Infection in Red Foxes in Four Animal Tuberculosis Endemic Areas in France. Microorganisme 2020, 8, 1070. [Google Scholar] [CrossRef]
- Réveillaud, É.; Desvaux, S.; Boschiroli, M.-L.; Hars, J.; Faure, É.; Fediaevsky, A.; Cavalerie, L.; Chevalier, F.; Jabert, P.; Poliak, S.; et al. Infection of Wildlife by Mycobacterium Bovis in France Assessment Through a National Surveillance System, Sylvatub. Front. Vet. Sci. 2018, 5, 262. [Google Scholar] [CrossRef]
- Lyashchenko, K.P.; Greenwald, R.; Esfandiari, J.; O’Brien, D.J.; Schmitt, S.M.; Palmer, M.V.; Waters, W.R. Rapid Detection of Serum Antibody by Dual-Path Platform VetTB Assay in White-Tailed Deer Infected with Mycobacterium Bovis. Clin. Vaccine Immunol. 2013, 20, 907–911. [Google Scholar] [CrossRef][Green Version]
- Wangoo, A.; Johnson, L.; Gough, J.; Ackbar, R.; Inglut, S.; Hicks, D.; Spencer, Y.; Hewinson, G.; Vordermeier, M. Advanced Granulomatous Lesions in Mycobacterium Bovis-Infected Cattle Are Associated with Increased Expression of Type I Procollagen, Gammadelta (WC1+) T Cells and CD 68+ Cells. J. Comp. Pathol. 2005, 133, 223–234. [Google Scholar] [CrossRef]
- Chambers, M.A.; Aldwell, F.; Williams, G.A.; Palmer, S.; Gowtage, S.; Ashford, R.; Dalley, D.J.; Davé, D.; Weyer, U.; Salguero, F.J.; et al. The Effect of Oral Vaccination with Mycobacterium Bovis BCG on the Development of Tuberculosis in Captive European Badgers (Meles Meles). Front. Cell Infect. Microbiol. 2017, 7, 6. [Google Scholar] [CrossRef]
- King, H.C.; Murphy, A.; James, P.; Travis, E.; Porter, D.; Hung, Y.-J.; Sawyer, J.; Cork, J.; Delahay, R.J.; Gaze, W.; et al. The Variability and Seasonality of the Environmental Reservoir of Mycobacterium Bovis Shed by Wild European Badgers. Sci. Rep. 2015, 5, 12318. [Google Scholar] [CrossRef]
- Lesellier, S.; Boschiroli, M.-L.; Barrat, J.; Wanke, C.; Salguero, F.J.; Garcia-Jimenez, W.L.; Nunez, A.; Godinho, A.; Spiropoulos, J.; Palmer, S.; et al. Detection of Live M. Bovis BCG in Tissues and IFN-Γresponses in European Badgers (Meles Meles) Vaccinated by Oropharyngeal Instillation or Directly in the Ileum. BMC Vet. Res. 2019, 15, 2166. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, T.; Labes, R.E.; Lambeth, M.; Montgomery, H.; Griffin, J.F.; Mackintosh, C.G. Transmission of Mycobacterium Bovis from Experimentally Infected Ferrets to Non-Infected Ferrets (Mustela Furo). N. Zealand Vet. J. 2000, 48, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, T.; Labes, R.E.; Cross, M.L.; Griffin, J.F.; Mackintosh, C.G. Partial Protection against Oral Challenge with Mycobacterium Bovis in Ferrets (Mustela Furo) Following Oral Vaccination with BCG. Int. J. Tuberc. Lung. Dis. 1999, 3, 1025–1033. [Google Scholar] [PubMed]
- Corner, L.A.L.; Costello, E.; Lesellier, S.; O’Meara, D.; Sleeman, D.P.; Gormley, E. Experimental Tuberculosis in the European Badger (Meles Meles) after Endobronchial Inoculation of Mycobacterium Bovis: I. Pathology and Bacteriology. Res. Vet. Sci. 2007, 83, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Lesellier, S.; Corner, L.; Costello, E.; Lyashchenko, K.; Greenwald, R.; Esfandiari, J.; Singh, M.; Hewinson, R.G.; Chambers, M.; Gormley, E. Immunological Responses and Protective Immunity in BCG Vaccinated Badgers Following Endobronchial Infection with Mycobacterium Bovis. Vaccine 2009, 27, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Lesellier, S.; Corner, L.; Costello, E.; Sleeman, P.; Lyashchenko, K.; Greenwald, R.; Esfandiari, J.; Singh, M.; Hewinson, R.G.; Chambers, M.; et al. Antigen Specific Immunological Responses of Badgers (Meles Meles) Experimentally Infected with Mycobacterium Bovis. Vet. Immunol. Immunopathol. 2008, 122, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Ashford, R.T.; Anderson, P.; Waring, L.; Davé, D.; Smith, F.; Delahay, R.J.; Gormley, E.; Chambers, M.A.; Sawyer, J.; Lesellier, S. Evaluation of the Dual Path Platform (DPP) VetTB Assay for the Detection of Mycobacterium Bovis Infection in Badgers. Prev. Vet. Med. 2020, 180, 105005. [Google Scholar] [CrossRef]
- Chambers, M.A.; Crawshaw, T.; Waterhouse, S.; Delahay, R.; Hewinson, R.G.; Lyashchenko, K.P. Validation of the BrockTB Stat-Pak Assay for Detection of Tuberculosis in Eurasian Badgers ( Meles Meles ) and Influence of Disease Severity on Diagnostic Accuracy. J. Clin. Microbiol. 2008, 46, 1498–1500. [Google Scholar] [CrossRef]
- Chambers, M.A.; Rogers, F.; Delahay, R.J.; Lesellier, S.; Ashford, R.; Dalley, D.; Gowtage, S.; Davé, D.; Palmer, S.; Brewer, J.; et al. Bacillus Calmette-Guérin Vaccination Reduces the Severity and Progression of Tuberculosis in Badgers. Proc. R. Soc. B. 2011, 278, 1913–1920. [Google Scholar] [CrossRef] [PubMed]
- Corner, L.A.L.; Costello, E.; O’Meara, D.; Lesellier, S.; Aldwell, F.E.; Singh, M.; Hewinson, R.G.; Chambers, M.A.; Gormley, E. Oral Vaccination of Badgers (Meles Meles) with BCG and Protective Immunity against Endobronchial Challenge with Mycobacterium Bovis. Vaccine 2010, 28, 6265–6272. [Google Scholar] [CrossRef] [PubMed]
- Balseiro, A.; Prieto, J.M.; Álvarez, V.; Lesellier, S.; Davé, D.; Salguero, F.J.; Sevilla, I.A.; Infantes-Lorenzo, J.A.; Garrido, J.M.; Adriaensen, H.; et al. Protective Effect of Oral BCG and Inactivated Mycobacterium Bovis Vaccines in European Badgers (Meles Meles) Experimentally Infected With M. Bovis. Front. Vet. Sci. 2020, 7, 41. [Google Scholar] [CrossRef]
- Vallejo, R.; García Marín, J.F.; Juste, R.A.; Muñoz-Mendoza, M.; Salguero, F.J.; Balseiro, A. Immunohistochemical Characterization of Tuberculous Lesions in Sheep Naturally Infected with Mycobacterium Bovis. BMC Vet. Res. 2018, 14, 154. [Google Scholar] [CrossRef] [PubMed]
- Salguero, F.J.; Gibson, S.; Garcia-Jimenez, W.; Gough, J.; Strickland, T.S.; Vordermeier, H.M.; Villarreal-Ramos, B. Differential Cell Composition and Cytokine Expression Within Lymph Node Granulomas from BCG-Vaccinated and Non-Vaccinated Cattle Experimentally Infected with Mycobacterium Bovis. Transbound. Emerg. Dis. 2017, 64, 1734–1749. [Google Scholar] [CrossRef]
- Muñoz-Mendoza, M.; Romero, B.; del Cerro, A.; Gortázar, C.; García-Marín, J.F.; Menéndez, S.; Mourelo, J.; de Juan, L.; Sáez, J.L.; Delahay, R.J.; et al. Sheep as a Potential Source of Bovine TB: Epidemiology, Pathology and Evaluation of Diagnostic Techniques. Transbound Emerg. Dis. 2015, 11, 12325. [Google Scholar] [CrossRef] [PubMed]
- García-Jiménez, W.L.; Fernández-Llario, P.; Gómez, L.; Benítez-Medina, J.M.; García-Sánchez, A.; Martínez, R.; Risco, D.; Gough, J.; Ortiz-Peláez, A.; Smith, N.H.; et al. Histological and Immunohistochemical Characterisation of Mycobacterium Bovis Induced Granulomas in Naturally Infected Fallow Deer (Dama Dama). Vet. Immunol. Immunopathol. 2012, 149, 66–75. [Google Scholar] [CrossRef]
- Gavier-Widén, D.; Cooke, M.M.; Gallagher, J.; Chambers, M.A.; Gortázar, C. A Review of Infection of Wildlife Hosts with Mycobacterium Bovis and the Diagnostic Difficulties of the “no Visible Lesion” Presentation. New Zealand Vet. J. 2009, 57, 122–131. [Google Scholar] [CrossRef]
- Lisle, G.; Yates, G.; Caley, P.; Corboy, R. Surveillance of Wildlife for Mycobacterium Bovis Infection Using Culture of Pooled Tissue Samples from Ferrets ( Mustela Furo ). New Zealand Vet. J. 2005, 53, 14–18. [Google Scholar] [CrossRef]
- García-Jiménez, W.L.; Benítez-Medina, J.M.; Fernández-Llario, P.; Abecia, J.A.; García-Sánchez, A.; Martínez, R.; Risco, D.; Ortiz-Peláez, A.; Salguero, F.J.; Smith, N.H.; et al. Comparative Pathology of the Natural Infections by Mycobacterium Bovis and by Mycobacterium Caprae in Wild Boar ( Sus Scrofa ): Natural Infection of Wild Boar by Mycobacterium Bovis and Mycobacterium Caprae. Transbound Emerg. Dis. 2013, 60, 102–109. [Google Scholar] [CrossRef]
- Corner, L.A.; Barrett, R.H.; Lepper, A.W.; Lewis, V.; Pearson, C.W. A Survey of Mycobacteriosis of Feral Pigs in the Northern Territory. Aust. Vet. J. 1981, 57, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Perrett, S.; Lesellier, S.; Rogers, F.; Williams, G.A.; Gowtage, S.; Palmer, S.; Dalley, D.; Davé, D.; Weyer, U.; Wood, E.; et al. Assessment of the Safety of Bacillus Calmette-Guérin Vaccine Administered Orally to Badgers (Meles Meles). Vaccine 2018, 36, 1990–1995. [Google Scholar] [CrossRef] [PubMed]
- Naranjo, V.; Gortazar, C.; Vicente, J.; de la Fuente, J. Evidence of the Role of European Wild Boar as a Reservoir of Mycobacterium Tuberculosis Complex. Vet. Microbiol. 2008, 127, 1–9. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fox ID | Sex | Age (in Years) | ELISA Idexx | MAPIA | End-point (T3–12 wpi) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| T0 | T1 | T2 | T3 | Antigens Recognized (T1–T3) | Tissues with | ||||||
| (4 wpi) | (8 wpi) | (12 wpi) | IgG Response | IgM Response | Granuloma | Culture Positive | PCR Positive | ||||
| A | M | 1 | - | + | - | + | B-PPD | - | 0 | 2 | 4 |
| B | F | 7 | - | + | - | + | - | DID38, DID65 | 4 | 4 | 7 |
| C | M | 1 | - | + | + | + | - | DID65 | 0 | 3 | 4 |
| D | F | 1 | - | - | - | - | None | 2 | 5 | 5 | |
| E | F | 3 | - | + | + | + | E6/P10, DID65, B-PPD | - | 1 | 3 | 5 |
| F | M | 1 | - | + | - | + | None | 0 | 0 | 3 | |
| G | M | 3 | + | + | + | + | E6/P10, DID38, B-PPD | MPB83, DID38 | 2 | 6 | 7 |
| H | F | 1 | - | + | + | - | None | 2 | 3 | 4 | |
| I | F | 7 | - | + | + | + | E6/P10, DID38, DID65, B-PPD | - | 3 | 6 | 7 |
| J | M | 3 | - | - | - | - | - | B-PPD | 1 | 3 | 6 |
| K | F | 3 | - | + | - | + | - | DID65 | 0 | 1 | 4 |
| L | M | 3 | - | + | - | + | MPB83, DID38, DID65 | MPB83, E6/P10, DID38, DID65 | 3 | 4 | 5 |
| Type of Sample | A | B | C | D | E | F | G | H | I | J | K | L | Total | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bac | PCR | Bac | PCR | Bac | PCR | Bac | PCR | Bac | PCR | Bac | PCR | Bac | PCR | Bac | PCR | Bac | PCR | Bac | PCR | Bac | PCR | Bac | PCR | BAC | PCR | |
| Tonsils | + | + | + | + | + | + | + | + | + | + | - | - | + | + | + | + | + | + | - | + | - | + | - | - | 8 | 10 |
| Right retropharyngeal LN | - | + | + | + | + | + | + | + | + | + | - | + | - | + | + | + | + | + | + | + | - | + | - | - | 7 | 11 |
| Left retropharyngeal LN | + | + | + | + | + | + | + | + | - | + | - | + | + | + | + | + | + | + | - | - | - | + | + | + | 8 | 11 |
| Right mandibular LN | - | - | - | + | - | - | + | + | - | + | - | - | + | + | - | - | + | + | - | + | - | - | - | - | 3 | 6 |
| Left mandibular LN | - | - | + | + | - | - | - | - | + | + | - | - | + | + | - | - | + | + | - | - | - | - | - | + | 4 | 5 |
| Left parotid LN | - | - | - | - | - | + | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + | 1 | 2 |
| Heart | - | - | - | + | - | - | - | - | Abs | Abs | Abs | Abs | Abs | Abs | Abs | Abs | Abs | Abs | Abs | Abs | Abs | Abs | Abs | Abs | 0 | 1 |
| Hepatic LN | - | - | - | + | Abs | Abs | - | - | - | - | - | - | + | + | - | - | - | - | + | + | - | - | + | + | 3 | 4 |
| Liver | - | - | - | - | - | - | + | + | - | - | - | - | - | - | - | - | - | + | - | + | - | - | + | + | 2 | 4 |
| Mesenteric LN | - | + | - | - | - | - | - | - | - | - | - | + | + | + | - | - | + | + | + | + | + | + | - | - | 4 | 6 |
| Kidneys (L & R) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - | - | - | - | 0 | 1 |
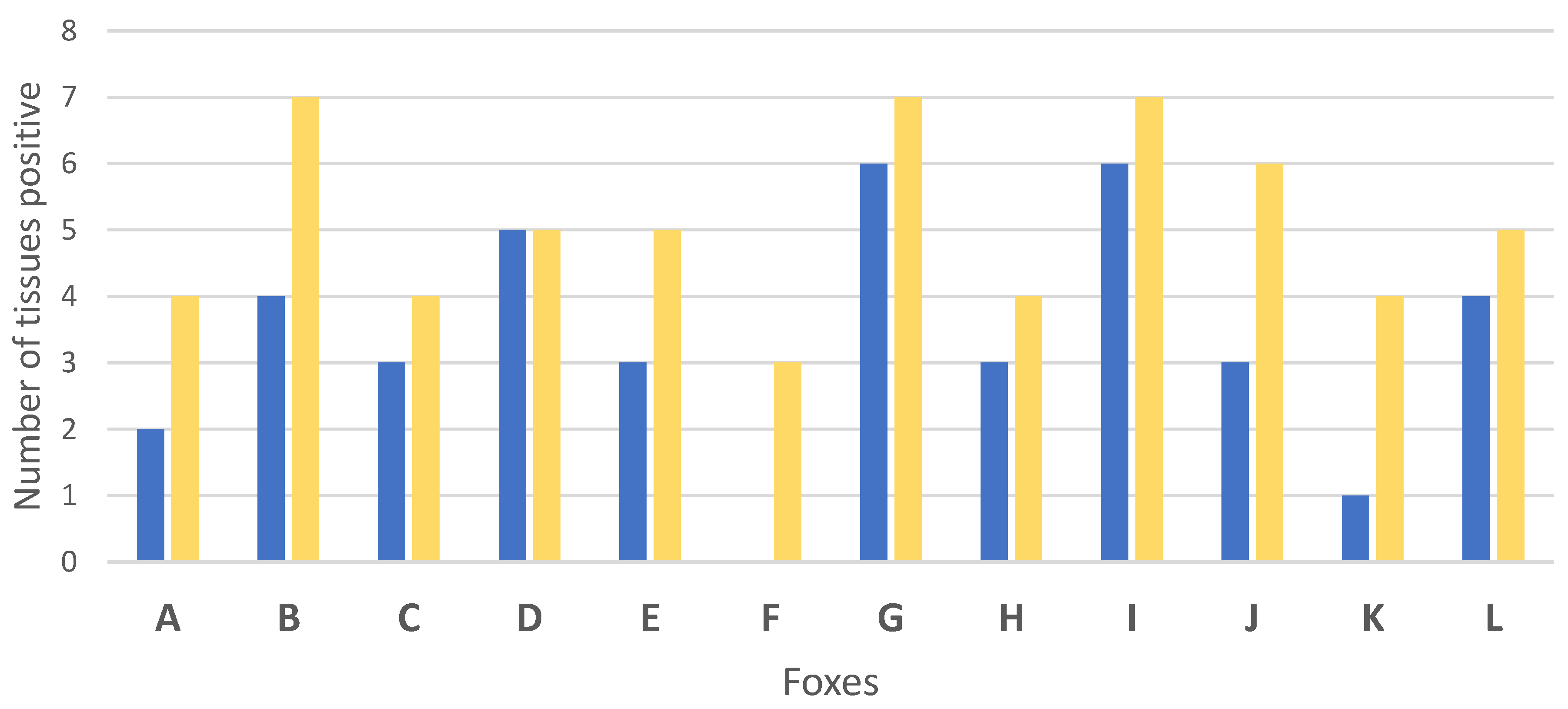
| Total | 2 | 4 | 4 | 7 | 3 | 4 | 5 | 5 | 3 | 5 | 0 | 3 | 6 | 7 | 3 | 4 | 6 | 7 | 3 | 6 | 1 | 4 | 4 | 5 | 40 | 61 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Richomme, C.; Lesellier, S.; Salguero, F.J.; Barrat, J.L.; Boucher, J.-M.; Reyes-Reyes, J.D.; Hénault, S.; De Cruz, K.; Tambosco, J.; Michelet, L.; et al. Experimental Infection of Captive Red Foxes (Vulpes vulpes) with Mycobacterium bovis. Microorganisms 2022, 10, 380. https://doi.org/10.3390/microorganisms10020380
Richomme C, Lesellier S, Salguero FJ, Barrat JL, Boucher J-M, Reyes-Reyes JD, Hénault S, De Cruz K, Tambosco J, Michelet L, et al. Experimental Infection of Captive Red Foxes (Vulpes vulpes) with Mycobacterium bovis. Microorganisms. 2022; 10(2):380. https://doi.org/10.3390/microorganisms10020380
Chicago/Turabian StyleRichomme, Céline, Sandrine Lesellier, Francisco Javier Salguero, Jacques Laurent Barrat, Jean-Marc Boucher, Jennifer Danaidae Reyes-Reyes, Sylvie Hénault, Krystel De Cruz, Jennifer Tambosco, Lorraine Michelet, and et al. 2022. "Experimental Infection of Captive Red Foxes (Vulpes vulpes) with Mycobacterium bovis" Microorganisms 10, no. 2: 380. https://doi.org/10.3390/microorganisms10020380
APA StyleRichomme, C., Lesellier, S., Salguero, F. J., Barrat, J. L., Boucher, J.-M., Reyes-Reyes, J. D., Hénault, S., De Cruz, K., Tambosco, J., Michelet, L., Boutet, J., Elahi, R., Lyashchenko, K. P., O’Halloran, C., Balseiro, A., & Boschiroli, M. L. (2022). Experimental Infection of Captive Red Foxes (Vulpes vulpes) with Mycobacterium bovis. Microorganisms, 10(2), 380. https://doi.org/10.3390/microorganisms10020380