
Genomic Analysis of Escherichia coli Longitudinally Isolated from Broiler Breeder Flocks after the Application of an Autogenous Vaccine

 , ,
, ,  and
and 
Abstract
1. Introduction
2. Material and Methods
2.1. Study Design
2.2. Bacterial Strain Selection and DNA Isolation
2.3. DNA Sequencing and Deposition
2.4. Phylogenetic Analysis
2.5. Antimicrobial Resistance Gene Analysis
2.6. Statistical Analysis
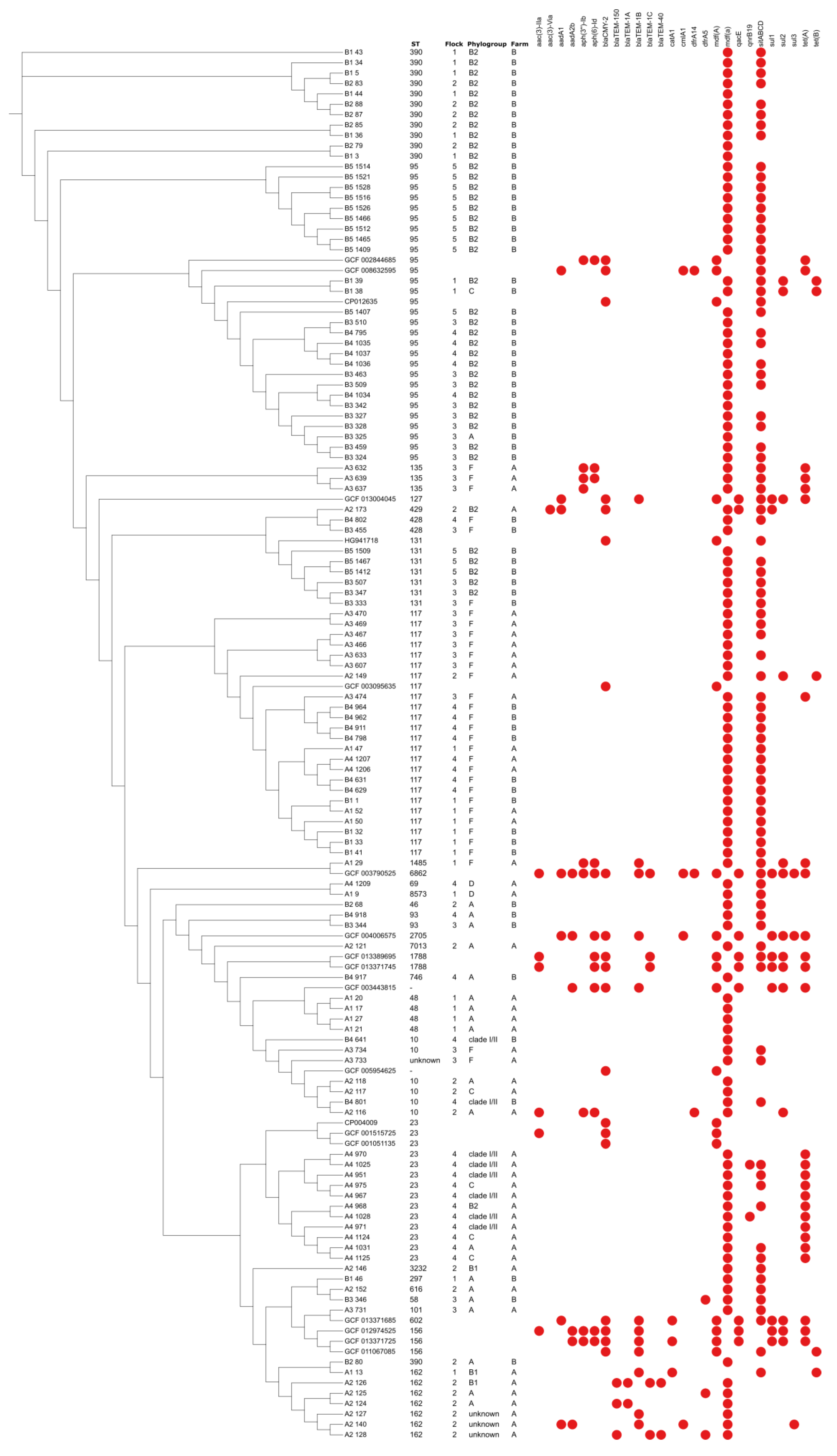
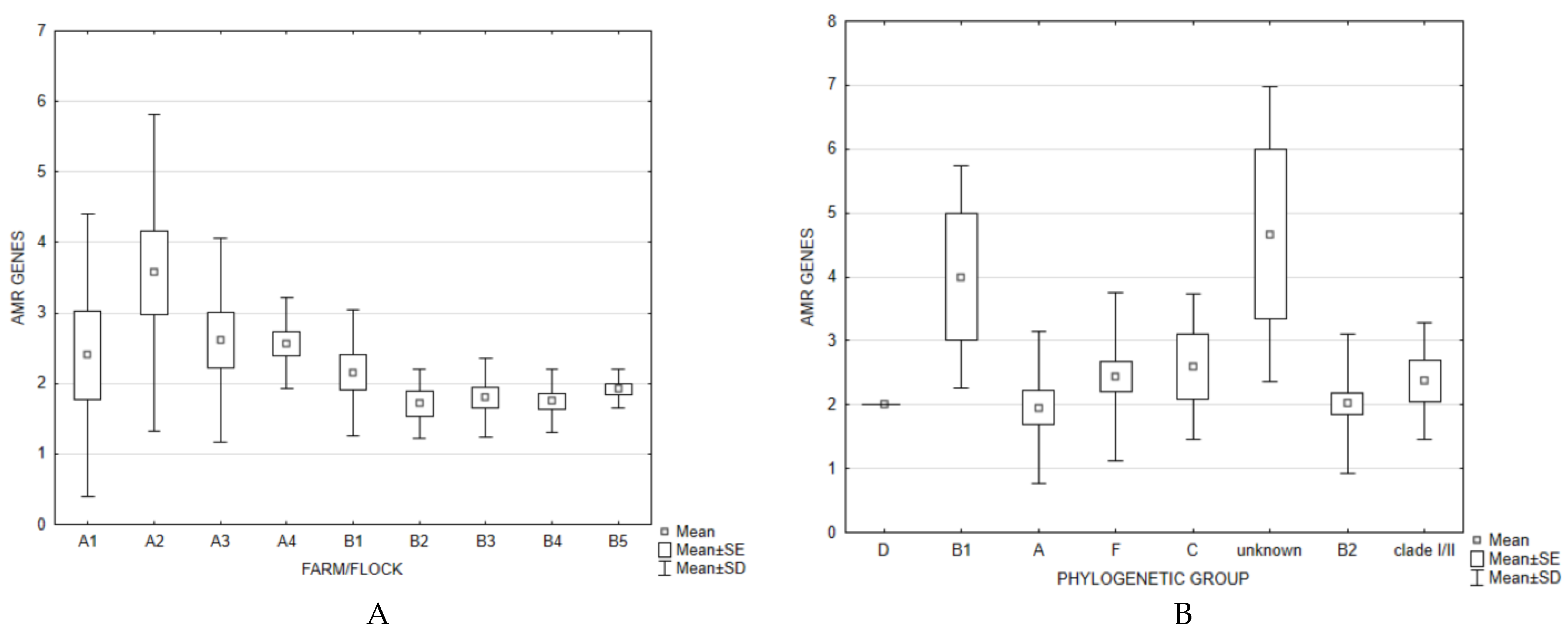
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nolan, L.K.; Vaillancourt, J.-P.; Barbieri, N.L.; Logue, C.M. Colibacillosis. In Diseases of Poultry, 14th ed.; Swayne, D.E., Ed.; Wiley-Blackwell: Ames, IA, USA, 2020; pp. 770–860. [Google Scholar]
- Poulsen, L.L.; Kudirkiene, E.; Jørgensen, S.L.; Djordjevic, S.P.; Cummins, M.L.; Christensen, J.P.; Christensen, H.; Bisgaard, M.; Thøfner, I. Whole genome sequence comparison of avian pathogenic Escherichia coli form acute and chronic salpingitis of egg laying hens. BMC Vet. Res. 2020, 16, 148. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Siek, K.; Giddings, C.W.; Doetkott, C.; Johnson, T.J.; Fakhr, M.K.; Nolan, L.K. Comparison of Escherichia coli isolates implicated in human urinary tract infection and avian colibacillosis. Microbiology 2005, 151, 2097–2110. [Google Scholar] [CrossRef]
- Ewers, C.; Li, G.; Wilking, H.; Kießling, S.; Alt, K.; Antáo, E.M.; Laturnus, C.; Diehl, I.; Glodde, S.; Homeier, T.; et al. Avian pathogenic, uropathogenic, and newborn meningitis-causing Escherichia coli: How closely related are they? Int. J. Med. Microbiol. 2007, 297, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Moulin-Schouleur, M.; Répérant, M.; Laurent, S.; Brée, A.; Mignon-Grasteau, S.; Germon, P.; Rasschaert, D.; Schouler, C. Extraintestinal pathogenic Escherichia coli strains of avian and human origin: Link between phylogenetic relationships and common virulence patterns. J. Clin. Microbiol. 2007, 45, 3366–3376. [Google Scholar] [CrossRef]
- Daga, A.P.; Koga, V.L.; Soncini, J.G.M.; de Matos, C.M.; Perugini, M.R.E.; Pelisson, M.; Kobayashi, R.K.T.; Vespero, E.C. Escherichia coli bloodstream infections in patients at a University hospital: Virulence factors and clinical characteristics. Front. Cell. Infect. Microbiol. 2019, 9, 191. [Google Scholar] [CrossRef]
- Nolan, L.K.; Logue, C.M.; Barbieri, N.L. APEC: A global one health pathogen and the myths that complicate its control. In Proceedings of the XXIst World Veterinary Poultry Association Congress, Bangkok, Thailand, 16–20 September 2019; pp. 86–90. [Google Scholar]
- Zhang, H.; Chen, X.; Nolan, L.K.; Zhang, W.; Li, G. Identification of host adaptation genes to extraintestinal pathogenic Escherichia coli during infection in different hosts. Infect. Immun. 2019, 87, e00666-19. [Google Scholar] [CrossRef]
- Christensen, H.; Bachmeier, J.; Bisgaard, M. New strategies to prevent and control avian pathogenic Escherichia coli (APEC). Avian Pathol. 2021, 50, 370–381. [Google Scholar] [CrossRef]
- Jordan, F.T.W.; Williams, N.J.; Wattret, A.; Jones, T. Observations on salpingitis, peritonitis and salpingoperitonitis in a layer breeder flock. Vet. Rec. 2005, 157, 573–577. [Google Scholar] [CrossRef]
- Li, L.; Thøfner, I.; Christensen, J.P.; Ronco, T.; Pedersen, K.; Olsen, R.H. Evaluation of the efficacy of an autogenous Escherichia coli vaccine in broiler breeders. Avian Pathol. 2017, 46, 300–308. [Google Scholar] [CrossRef]
- Rodriguez-Siek, K.; Giddings, C.; Doetkott, C.; Johnson, T.; Nolan, L. Characterizing the APEC pathotype. BMC Vet. Res. 2005, 36, 241–256. [Google Scholar] [CrossRef]
- Dziva, F.; Stevens, M.P. Colibacillosis in poultry: Unravelling the molecular basis of virulence of avian pathogenic Escherichia coli in their natural hosts. Avian Pathol. 2008, 37, 355–366. [Google Scholar] [CrossRef]
- Johnson, T.J.; Jordan, D.; Kariyawasam, S.; Stell, A.L.; Bell, N.P.; Wannemuehler, Y.M.; Fernández-Alarcón, C.; Li, G.; Tivendale, K.A.; Logue, C.M.; et al. Sequence analysis and characterization of a transferable hybrid plasmid encoding multidrug resistance and enabling zoonotic potential for extraintestinal Escherichia coli. Infect. Immun. 2010, 78, 1931–1942. [Google Scholar] [CrossRef]
- de Oliveira, A.L.; Rocha, D.A.; Finkler, F.; De Moraes, L.B.; Barbieri, N.L.; Pavanelo, D.B.; Winkler, C.; Grassotti, T.T.; De Brito, K.C.T.; De Brito, B.G.; et al. Prevalence of ColV plasmid-linked genes and in vivo pathogenicity of avian strains of Escherichia coli. Foodborne Pathog. Dis. 2015, 12, 679–685. [Google Scholar] [CrossRef]
- Johnson, T.J.; Siek, K.E.; Johnson, S.J.; Nolan, L.K. DNA sequence of a ColV plasmid and prevalence-encoded virulence genes among avian Escherichia coli strains. J. Bacteriol. 2006, 188, 745–758. [Google Scholar] [CrossRef]
- Jørgensen, S.L.; Stegger, M.; Kudirkiene, E.; Lilje, B.; Poulsen, L.L.; Ronco, T.; Pires Dos Santos, T.; Kiil, K.; Bisgaard, M.; Pedersen, K.; et al. Diversity and population overlap between avian and human Escherichia coli belonging to sequence type 95. mSphere 2019, 4, e00333-18. [Google Scholar] [CrossRef]
- Mageiros, L.; Méric, G.; Bayliss, S.C.; Pensar, J.; Pascoe, B.; Mourkas, E.; Calland, J.K.; Yahara, K.; Murray, S.; Wilkinson, T.S.; et al. Genome evaluation and the emergence of pathogenicity in avian Escherichia coli. Nat. Commun. 2021, 12, 765–767. [Google Scholar] [CrossRef]
- Sheppard, S.K.; Guttman, D.S.; Fitzgerald, J.R. Population genomics of bacterial host adaptation. Nat. Rev. Genet. 2018, 19, 549–565. [Google Scholar] [CrossRef]
- Mehat, J.W.; van Vliet, A.H.M.; La Ragione, R.M. The Avian Pathogenic Escherichia coli (APEC) pathotype is comprised of multiple distinct, independent genotypes. Avian Pathol. 2021, 50, 402–406. [Google Scholar] [CrossRef]
- Aworh, M.K.; Kwaga, J.K.P.; Hendriksen, R.S.; Okolocha, E.C.; Thakur, S. Genetic relatedness of multidrug resistant Escherichia coli isolated from humans, chickens and poultry environments. Antimicrob. Resist. Infect. Control 2021, 10, 58. [Google Scholar] [CrossRef]
- Rafique, M.; Potter, R.F.; Ferreiro, A.; Wallace, M.A.; Rahim, A.; Malik, A.A.; Siddique, N.; Abbas, M.A.; D’Souza, A.W.; Burnham, C.-A.D.; et al. Genomic characterization of antibiotic resistant Escherichia coli isolated from domestic chickens in Pakistan. Front. Microbiol. 2020, 10, 3052. [Google Scholar] [CrossRef]
- Vallejos-Vidal, E.; Reyes-Cerpa, S.; Rivas-Pardo, J.A.; Maisey, K.; Yañez, J.M.; Valenzuela, H.; Cea, P.A.; Castro-Fernandez, V.; Tort, L.; Sandino, A.M.; et al. Single-nucleotide polymorphisms (SNP) mining and their effect on the tridimensional protein structure prediction in a set of immunity-related expressed sequence tags (EST) in Atlantic salmon (Salmo salar). Front. Genet. 2020, 10, 1406. [Google Scholar] [CrossRef] [PubMed]
- Bush, S.J.; Foster, D.; Eyre, D.W.; Clark, E.L.; De Maio, N.; Shaw, L.P.; Stoesser, N.; Peto, T.E.A.; Crook, D.W.; Walker, A.S. Genomic diversity affects the accuracy of bacterial single-nucleotide polymorphism-calling pipelines. Gigascience 2020, 9, giaa007. [Google Scholar] [CrossRef] [PubMed]
- Lozica, L.; Repar, J.; Gottstein, Ž. Longitudinal study on the effect of autogenous vaccine application on the sequence type and virulence profiles of Escherichia coli in broiler breeder flocks. Vet. Microbiol. 2021, 259, 109159. [Google Scholar] [CrossRef] [PubMed]
- Gottstein, Ž.; Lozica, L.; Horvatek Tomić, D.; Lukač, M.; Prukner-Radovčić, E.; Vlahek, M.; Dolenčić, N.; Glavak, Ž. Production parameters after autogenous APEC E. coli vaccine application and improvement of vaccination program in broiler breeder flock. In Proceeding of the Poultry Days 2019, Poreč, Croatia, 8–11 May 2019; Balenović, M., Ed.; Croatian Veterinary Institute: Zagreb, Croatia, 2019; pp. 98–103. [Google Scholar]
- Chen, X.; Liu, W.; Li, H.; Yan, S.; Jiand, F.; Cai, W.; Li, G. Whole genome sequencing analysis of avian pathogenic Escherichia coli from China. Vet. Microbiol. 2021, 259, 109158. [Google Scholar] [CrossRef]
- Treangen, T.J.; Ondov, B.D.; Koren, S.; Phillippy, A.M. The Harvest suite for rapid core-genome alignment and visualization of thousands of intraspecific microbial genomes. Genome Biol. 2014, 15, 524–538. [Google Scholar] [CrossRef]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus sequence typing of total genome sequenced bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Ronco, T.; Stegger, M.; Olsen, R.H.; Sekse, C.; Nordstoga, A.B.; Pohjanvirta, T.; Lilje, B.; Lyhs, U.; Andersen, P.S.; Pedersen, K. Spread of avian pathogenic Escherichia coli ST117 O78:H4 in Nordic broiler production. BMC Genom. 2017, 18, 13. [Google Scholar] [CrossRef]
- Papouskova, A.; Masarikova, M.; Valcek, A.; Senk, A.; Cejkova, D.; Jahodarova, E.; Cizek, A. Genomic analysis of Escherichia coli strains isolated from diseased chicken in the Czech Republic. BMC Vet. Res. 2020, 16, 189–199. [Google Scholar] [CrossRef]
- Ramadan, H.; Jackson, C.R.; Frye, J.G.; Hiott, L.M.; Samir, M.; Awad, A.; Woodley, T.A. Antimicrobial resistance, genetic diversity and multilocus sequence typing of Escherichia coli from humans, retail chicken and ground beef in Egypt. Pathogens 2020, 9, 357. [Google Scholar] [CrossRef]
- Smolza, A.; Wami, H.; Dobrindt, U. Comparative genomics of emerging lineages and mobile resistomes of contemporary broiler strains of Salmonella Infantis and E. coli. Front. Microbiol. 2021, 12, 342. [Google Scholar] [CrossRef]
- Edgar, R.; Bibi, E. MdfA, an Escherichia coli multidrug resistance protein with an extraordinarily broad spectrum of drug recognition. J. Bacteriol. 1977, 179, 2274–2280. [Google Scholar] [CrossRef]
- Ong, K.H.; Khor, W.C.; Quek, J.Y.; Low, Z.X.; Arivalan, S.; Humaidi, M.; Chua, C.; Seow, K.L.G.; Guo, S.; Tay, M.Y.F.; et al. Occurrence and antimicrobial resistance traits of Escherichia coli from wild birds and rodents in Singapure. Int. J. Environ. Res. Public Health 2020, 17, 5606. [Google Scholar] [CrossRef]
- Sabri, M.; Léveillé, S.; Dozois, C.M. A SitABCD homologue from an avian pathogenic Escherichia coli strain mediates transport of iron and manganese and resistance to hydrogen peroxide. Microbiology 2006, 152, 745–758. [Google Scholar] [CrossRef]
- Sabri, M.; Caza, M.; Proulx, J.; Lymberopoulos, M.H.; Brée, A.; Moulin-Schouleur, M.; Curtiss, R., III; Dozois, C.M. Contribution of the SitABCD, MntH, and FeoB metal transporters to the virulence of avian pathogenic Escherichia coli O78 strain χ7122. Infec. Immun. 2008, 76, 601–611. [Google Scholar] [CrossRef]
- Maynard, C.; Bekal, S.; Sanschagrin, F.; Levesque, R.C.; Brousseau, R.; Masson, L.; Larivière, S.; Harel, J. Heterogeneity among virulence and antimicrobial resistance gene profiles of extraintestinal Escherichia coli isolates of animal and human origin. J. Clin. Microbiol. 2004, 42, 5444–5452. [Google Scholar] [CrossRef] [PubMed]
- Wilkerson, C.; Samadpour, M.; van Kirk, N.; Roberts, M.C. Antibiotic resistance and distribution of tetracycline resistance genes in Escherichia coli O157:H7 isolates from humans and bovines. Antimicrob. Agents Chemother. 2004, 48, 1066–1067. [Google Scholar] [CrossRef]
- Seifi, S.; Khoshbakht, R. Prevalence of tetracycline resistance determinants in broiler isolated Escherichia coli in Iran. Br. Poult. Sci. 2016, 57, 729–733. [Google Scholar] [CrossRef]
- Jahantigh, M.; Samadi, K.; Dizaji, R.E.; Salari, S. Antimicrobial resistance and prevalence of tetracycline resistance genes in Escherichia coli isolated from lesions of colibacillosis in broiler chickens in Sistan, Iran. BMC Vet. Res. 2020, 16, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Gharaibeh, M.H.; Shatnawi, S.Q. An overview of colistin resistance, mobilized colistin resistance genes dissemination, global responses, and the alternatives to colistin: A review. Vet. World 2019, 12, 1735–1746. [Google Scholar] [CrossRef] [PubMed]
- Koutsianos, D.; Athanasiou, L.V.; Dimitriou, T.; Nikolaidis, M.; Tsadila, C.; Amoutzias, G.; Mossialos, D.; Koutoulis, K.C. Antibiotic resistance patterns and mcr-1 detection in avian pathogenic Escherichia coli isolates from commercial layer and layer breeder flocks demonstrating colibacillosis in Greece. Microb. Drug Resist. 2021, 27, 710–720. [Google Scholar] [CrossRef]
- Massella, E.; Giacometty, F.; Bonilauri, P.; Reid, C.J.; Djordjevic, S.P.; Merialdi, G.; Bacci, C.; Fiorentini, L.; Massi, P.; Bardasi, L.; et al. Antimicrobial resistance profile and ExPEC virulence potential in commensal Escherichia coli of multiple sources. Antibiotics 2021, 10, 351. [Google Scholar] [CrossRef]
- Smith, J.L.; Fratamico, P.M.; Gunther, N.W. Extraintestinal pathogenic Escherichia coli. Foodborne Pathog. Dis. 2007, 4, 134–163. [Google Scholar] [CrossRef]
- Chakraborty, A.; Saralaya, V.; Adhikari, P.; Shenoy, S.; Baliga, S.; Hegde, A. Characterization of Escherichia coli phylogenetic groups associated with extraintestinal infections in South Indian population. Ann. Med. Health Sci. Res. 2015, 5, 241–246. [Google Scholar] [CrossRef]
- Lozica, L.; Ekert Kabalin, A.; Dolenčić, N.; Vlahek, M.; Gottstein, Ž. Phylogenetic characterization of avian pathogenic Escherichia coli strains longitudinally isolated from broiler breeder flocks vaccinated with autogenous vaccine. Poult. Sci. J. 2021, 100, 101079. [Google Scholar] [CrossRef]
- Landman, W.J.M.; van Eck, J.H.H. The efficacy of inactivated Escherichia coli autogenous vaccines against the E. coli peritonitis syndrome in layers. Avian Pathol. 2017, 46, 658–665. [Google Scholar] [CrossRef]
- Koutsianos, D.; Gantelet, H.; Franzo, G.; Lecoupeur, M.; Thibault, E.; Cecchinato, M.; Koutoulis, K.C. An assessment of the level of protection against colibacillosis conferred by several autogenous and/or commercial vaccination programs in conventional pullets upon experimental challenge. Vet. Sci. 2020, 7, 80. [Google Scholar] [CrossRef] [PubMed]
- Koutsianos, D.; Athanasiou, L.V.; Spyropoulou, M.; Prentza, Z.; Dedousi, A.; Polizopoulou, Z.; Mossialos, D.; Koutoulis, K. Evaluation of hematological variables in layer pullets after vaccination and challenge with E. coli. Comp. Clin. Pathol. 2021, 30, 113–118. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Farm | Flock | Number of Analysed Strains per Flock | Vaccination | Age at the Time of Vaccination | Treatment |
|---|---|---|---|---|---|
| A | 1 | 10 | Commercial vaccines (live attenuated + inactivated 2x) | 0 d 10 w 18 w | Doxycycline |
| 2 | 14 | Autogenous vaccine 2x | 10 w 18 w | Doxycycline, tiamulin | |
| 3 | 13 | Autogenous vaccine 2x | 12 w 19 w | - | |
| 4 | 14 | Autogenous vaccine 2x | 10 w 19 w | Amoxicillin, doxycycline, enrofloxacin, oxytetracycline | |
| B | 1 | 13 | Commercial vaccines (live attenuated + inactivated 2x) | 0 d 13 w 19 w | Doxycycline, enrofloxacin, tylosin |
| 2 | 7 | Commercial vaccines (live attenuated 2x) Autogenous vaccine 2x | 0 d 5 w 11 w 18 w | Amoxicilllin, polymyxin E, tylosin | |
| 3 | 15 | Autogenous vaccine 2x | 9 w 20 w | Doxycycline, polymyxin E | |
| 4 | 16 | Autogenous vaccine 2x | 10 w 18 w | Doxycycline, enrofloxacin | |
| 5 | 13 | Autogenous vaccine 2x | 9 w 17 w | Amoxicillin, enrofloxacin |
| AMR Gene | Phenotype | Number of AMR Genes on Farm A * n = 51 | Number of AMR Genes on Farm B ** n = 64 | Total Number of AMR Genes |
|---|---|---|---|---|
| aac(3)-IIa | apramycin, gentamicin, tobramycin, dibekacin, netilmicin, sisomicin | 1 (1.96) | 0 | 1 (0.87) |
| aac(3)-VIa | gentamicin | 1 (1.96) | 0 | 1 (0.87) |
| aadA1 | spectinomycin | 2 (3.92) | 0 | 2 (1.74) |
| aadA2b | spectinomycin, streptomycin | 1 (1.96) | 0 | 1 (0.87) |
| aph(3″)-Ib | streptomycin | 5 (9.8) | 0 | 5 (4.35) |
| aph(6)-Id | streptomycin | 5 (9.81) | 0 | 5 (4.35) |
| blaCMY-2 | amoxicillin, amoxicillin + clavulanic acid, ampicillin, ampicillin + clavulanic acid, cefotaxime, cefotxitin, ceftazidime, piperacillin, piperacillin + tazobactam, ticarcillin, ticarcillin + clavulanic acid | 1 (1.96) | 0 | 1 (0.87) |
| blaTEM-1A | amoxicillin, ampicillin, cephalothin, piperacillin, ticarcillin | 3 (5.88) | 0 | 3 (2.61) |
| blaTEM-1B | amoxicillin, ampicillin, cephalothin, piperacillin, ticarcillin | 4 (7.84) | 0 | 4 (3.48) |
| blaTEM-1C | amoxicillin, ampicillin, cephalothin, piperacillin, ticarcillin | 2 (3.92) | 0 | 2 (1.74) |
| blaTEM-40 | amoxicillin, amoxicillin + clavulanic acid, ampicillin, ampicillin + clavulanic acid, piperacillin, piperacillin + tazobactam, ticarcillin, ticarcillin + clavulanic acid | 2 (3.92) | 0 | 2 (1.74) |
| blaTEM-150 | unknown beta-lactam | 3 (5.88) | 0 | 3 (2.61) |
| catA1 | chloramphenicol | 1 (1.96) | 0 | 1 (0.87) |
| cmIA1 | chloramphenicol | 1 (1.96) | 0 | 1 (0.87) |
| dfrA5 | trimethoprim | 2 (3.92) | 1 (1.56) | 3 (2.61) |
| dfrA14 | trimethoprim | 1 (1.96) | 0 | 1 (0.87) |
| mdf(A) | unknown macrolide, aminoglycoside, tetracycline, fluoroquinolone, phenicol and rifamycin | 51 (100) | 64 (100) | 115 (100) |
| qacE | benzylkonium chloride, ethidium bromide, chlorhexidine, cetylpyridinium | 1 (1.96) | 0 | 1 (0.87) |
| qnrB19 | ciprofloxacin | 2 (3.92) | 0 | 2 (1.74) |
| sitABCD | hydrogen peroxide | 31 (60.78) | 51 (79.69) | 82 (71.3) |
| sul1 | sulfamethoxazole | 1 (1.96) | 0 | 1 (0.87) |
| sul2 | sulfamethoxazole | 3 (5.88) | 2 (3.13) | 5 (4.35) |
| sul3 | sulfamethoxazole | 1 (1.96) | 0 | 1 (0.87) |
| tet(A) | doxycycline, tetracycline | 17 (33.33) | 0 | 17 (14.78) |
| tet(B) | doxycycline, tetracycline, minocycline | 2 (3.92) | 2 (3.13) | 4 (3.48) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lozica, L.; Villumsen, K.R.; Li, G.; Hu, X.; Maljković, M.M.; Gottstein, Ž. Genomic Analysis of Escherichia coli Longitudinally Isolated from Broiler Breeder Flocks after the Application of an Autogenous Vaccine. Microorganisms 2022, 10, 377. https://doi.org/10.3390/microorganisms10020377
Lozica L, Villumsen KR, Li G, Hu X, Maljković MM, Gottstein Ž. Genomic Analysis of Escherichia coli Longitudinally Isolated from Broiler Breeder Flocks after the Application of an Autogenous Vaccine. Microorganisms. 2022; 10(2):377. https://doi.org/10.3390/microorganisms10020377
Chicago/Turabian StyleLozica, Liča, Kasper Rømer Villumsen, Ganwu Li, Xiao Hu, Maja Maurić Maljković, and Željko Gottstein. 2022. "Genomic Analysis of Escherichia coli Longitudinally Isolated from Broiler Breeder Flocks after the Application of an Autogenous Vaccine" Microorganisms 10, no. 2: 377. https://doi.org/10.3390/microorganisms10020377
APA StyleLozica, L., Villumsen, K. R., Li, G., Hu, X., Maljković, M. M., & Gottstein, Ž. (2022). Genomic Analysis of Escherichia coli Longitudinally Isolated from Broiler Breeder Flocks after the Application of an Autogenous Vaccine. Microorganisms, 10(2), 377. https://doi.org/10.3390/microorganisms10020377