
Probiotic Effects of a Marine Purple Non-Sulfur Bacterium, Rhodovulum sulfidophilum KKMI01, on Kuruma Shrimp (Marsupenaeus japonicus)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Condition
2.2. Small-Scale (3 Days) Aquaria Experiments to Examine the Effects of R. sulfidophilum KKMI01 on the Gene Expression in M. japonicus
2.3. Long-Term (145 Days) 200-Ton Aquaria Experiments
2.4. qRT-PCR
2.5. Statistical Analysis
3. Results
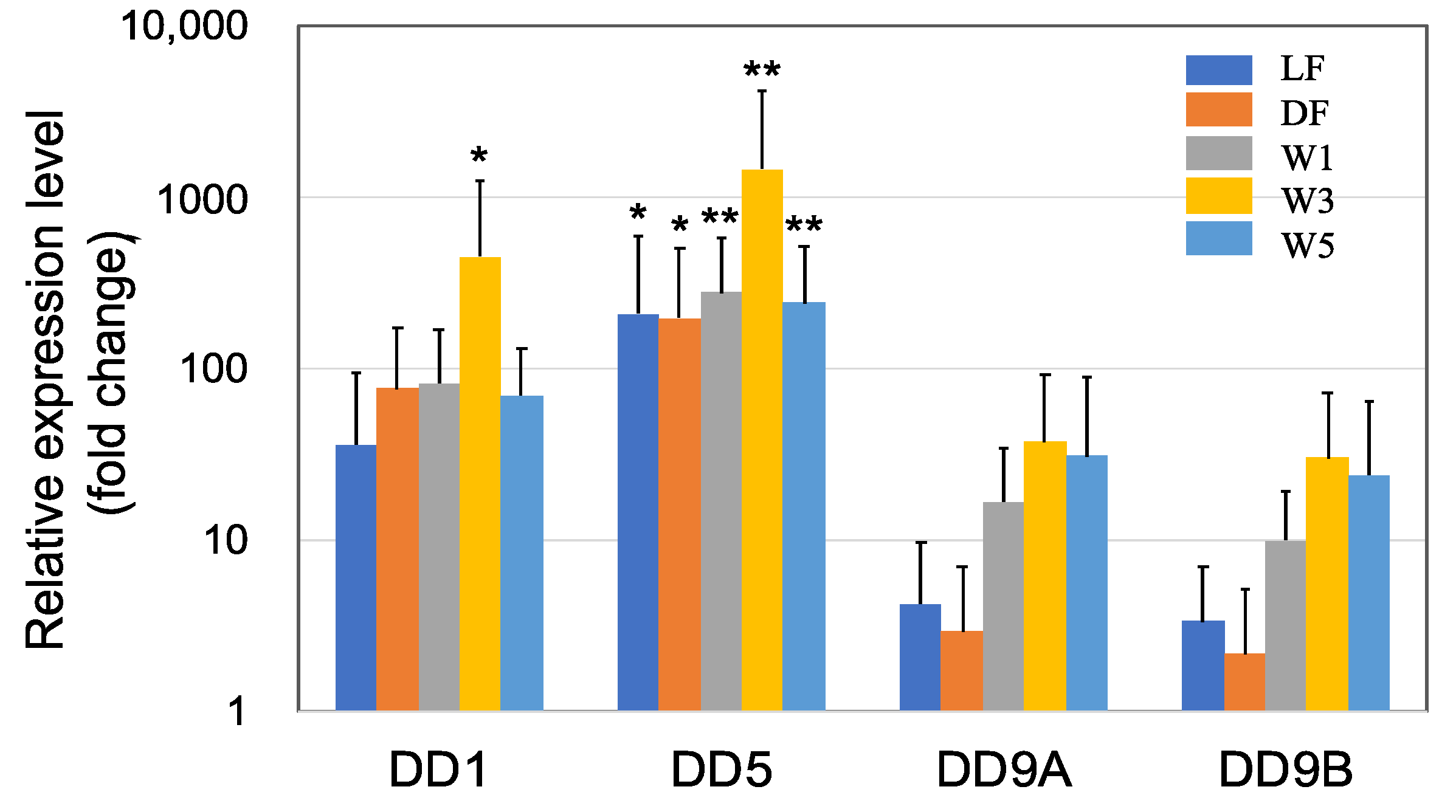
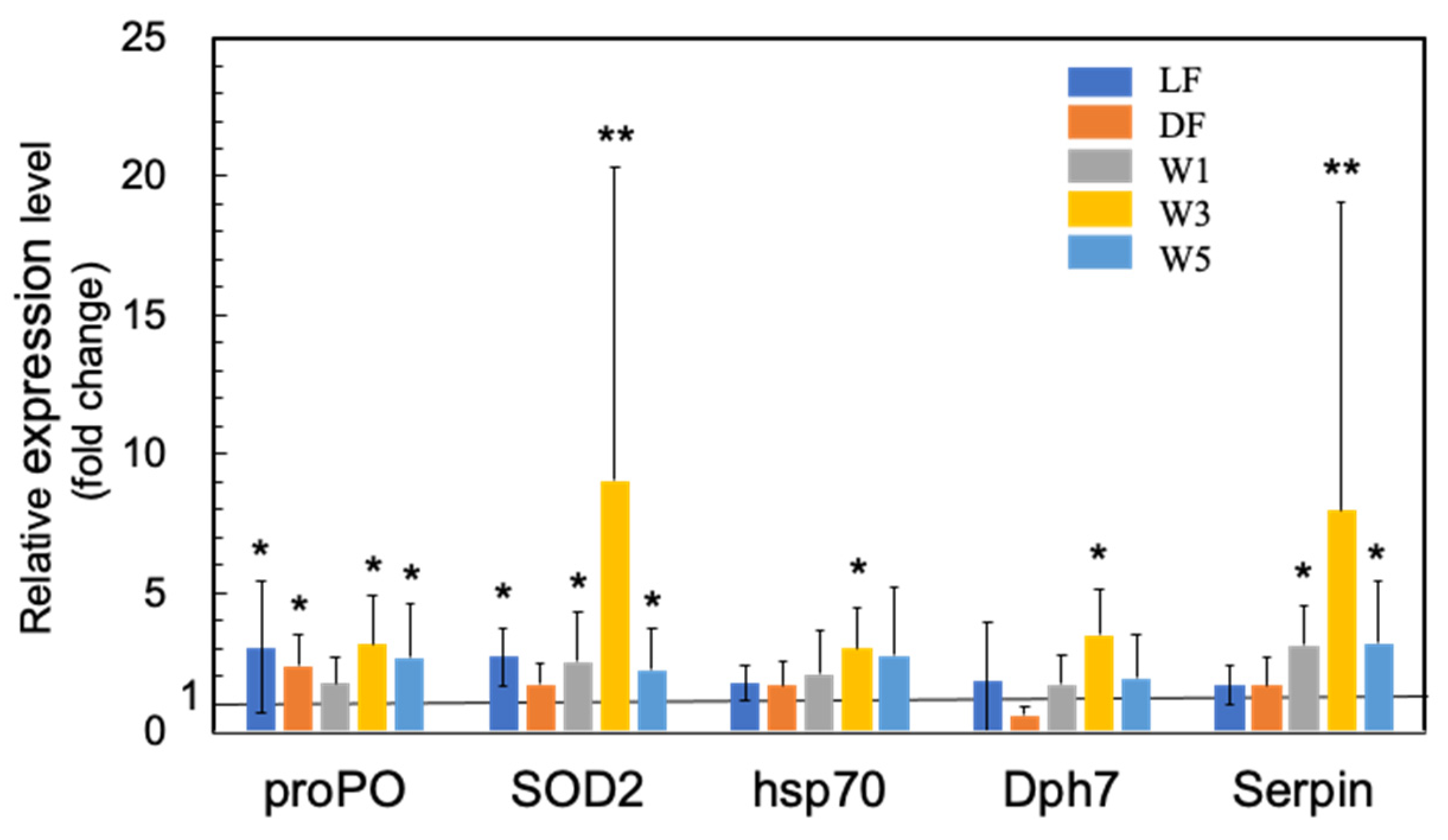
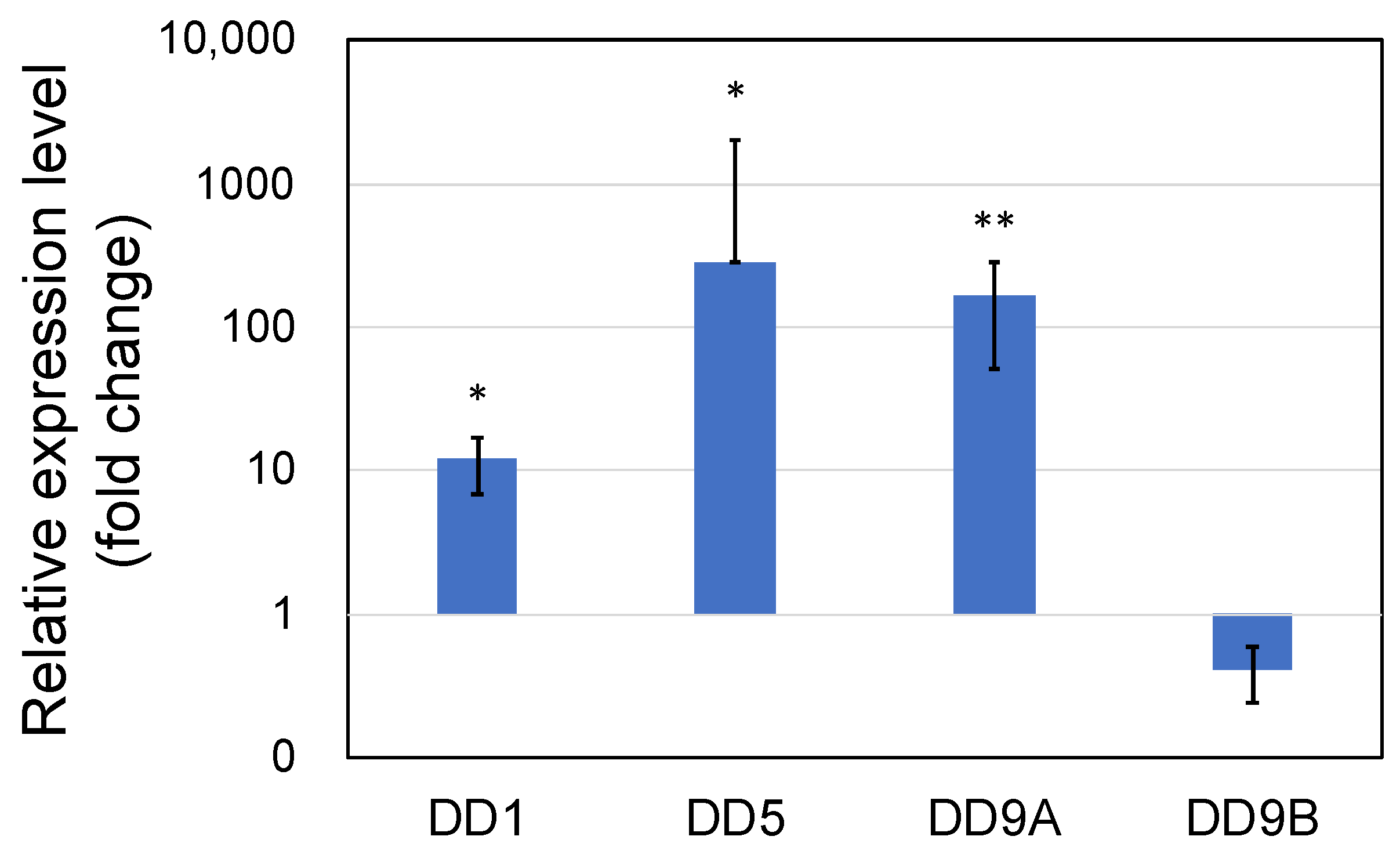
3.1. Genes Upregulated by PNSB Treatment in Short-Term Experiments
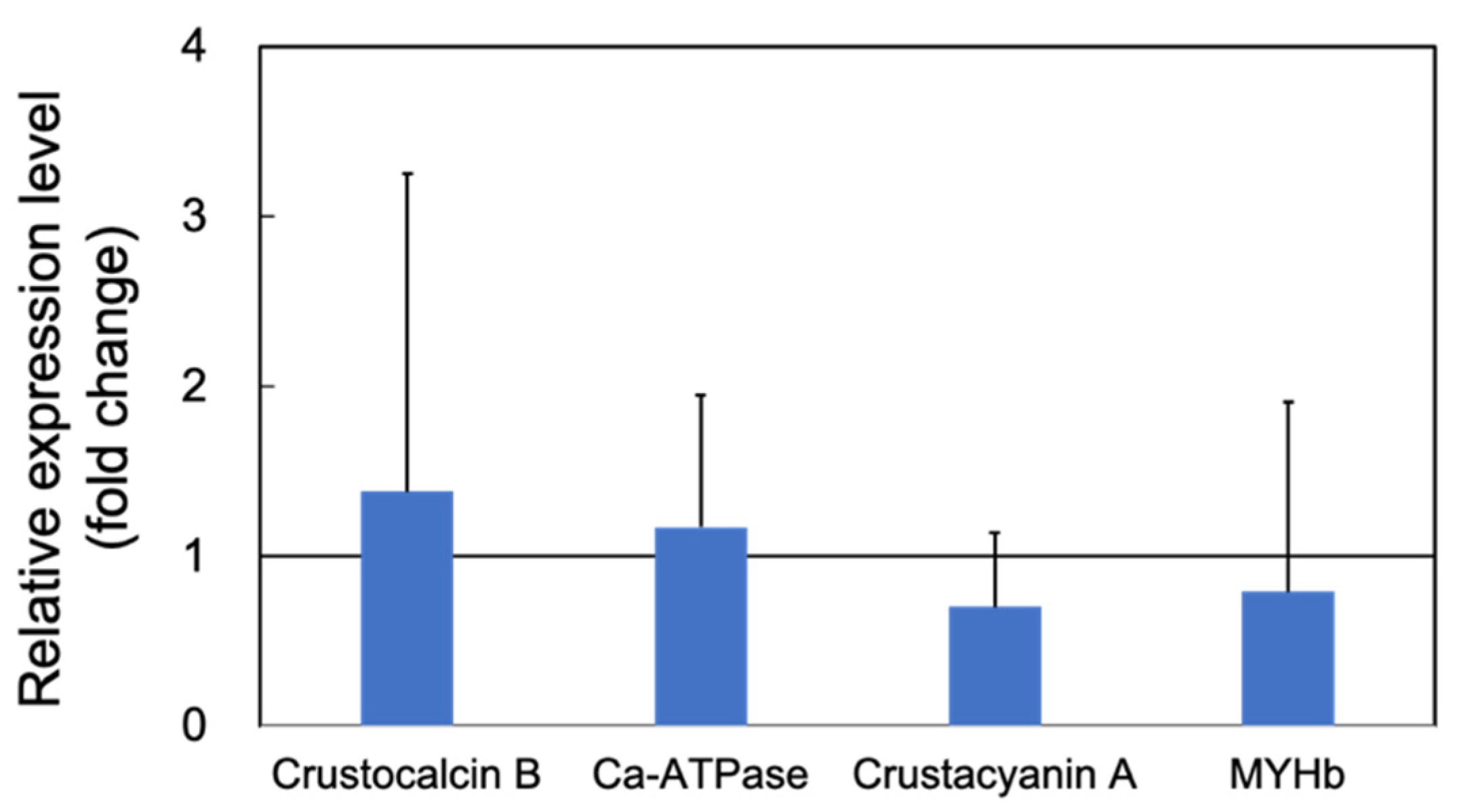
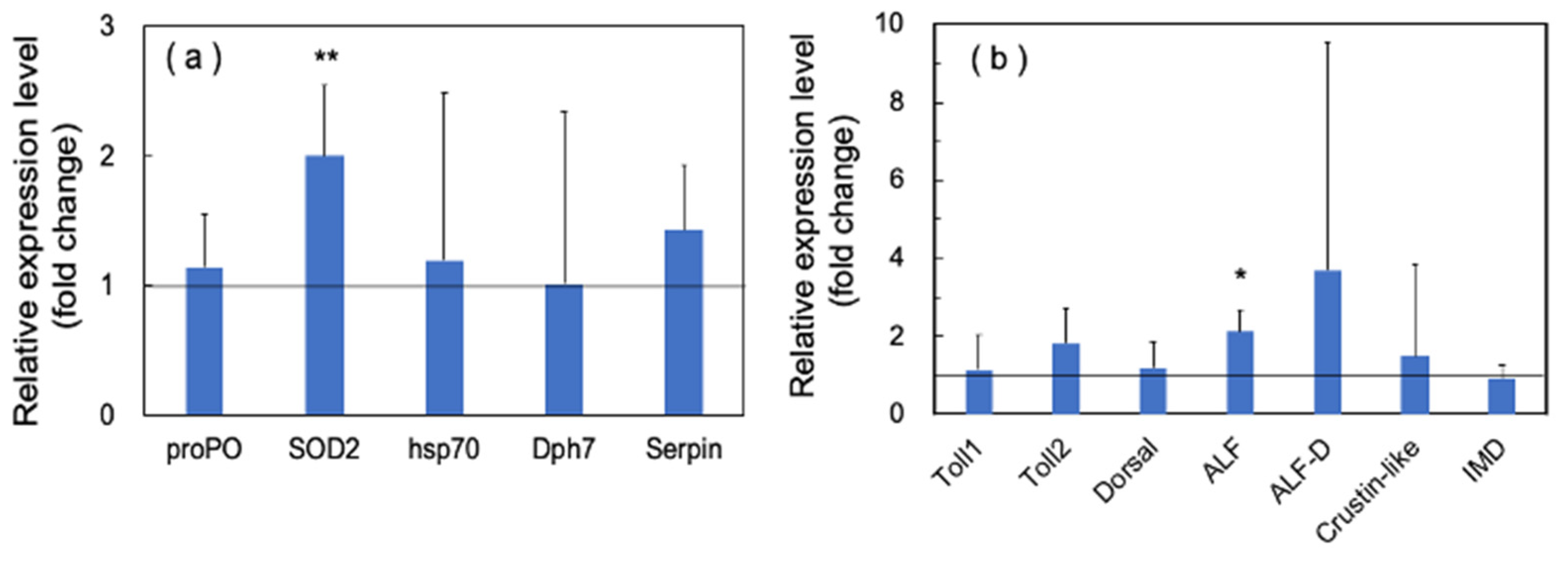
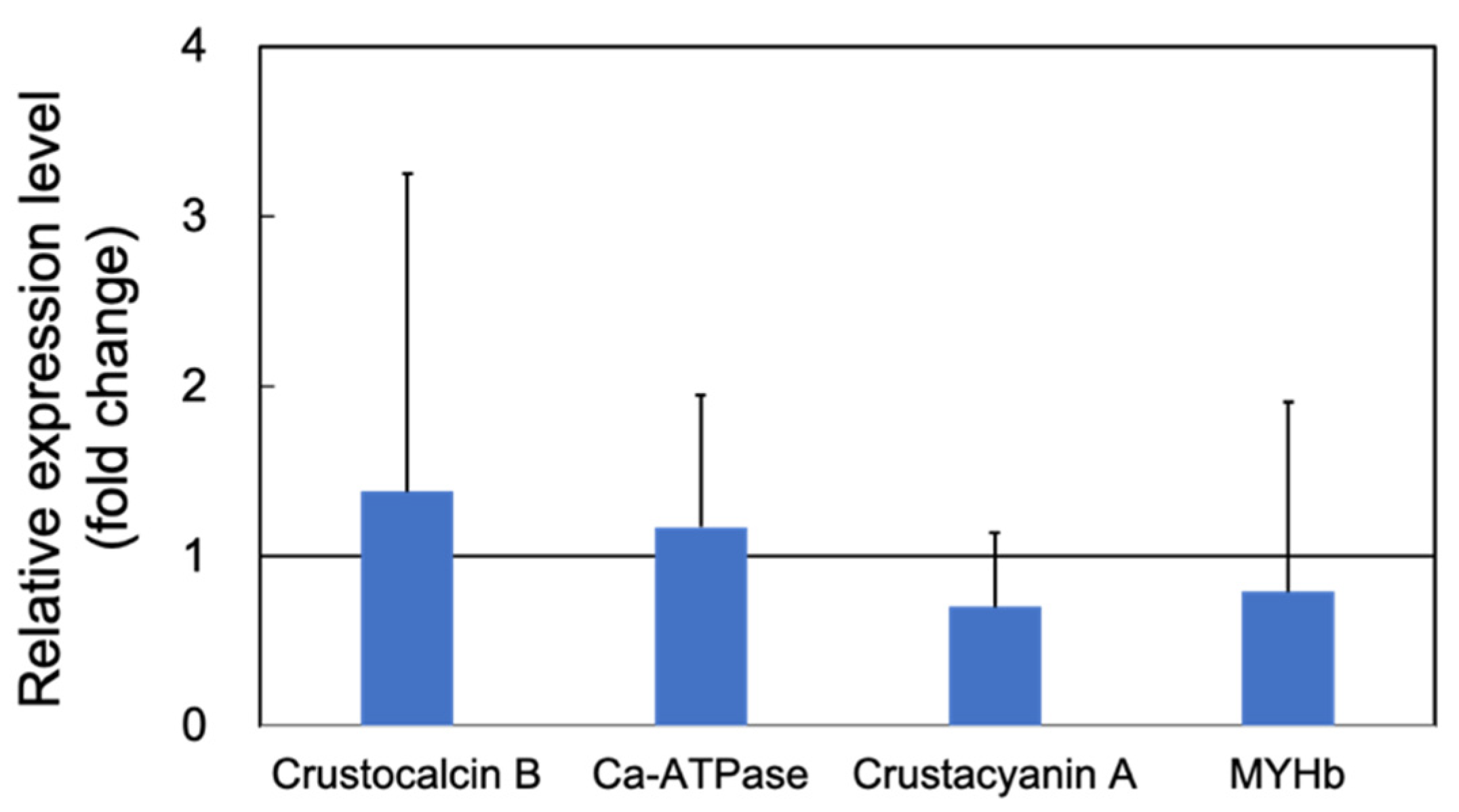
3.1.1. Upregulation of the Genes Related to Molting by PNSB
3.1.2. Upregulation of the Innate Immunity Genes by PNSB Treatment
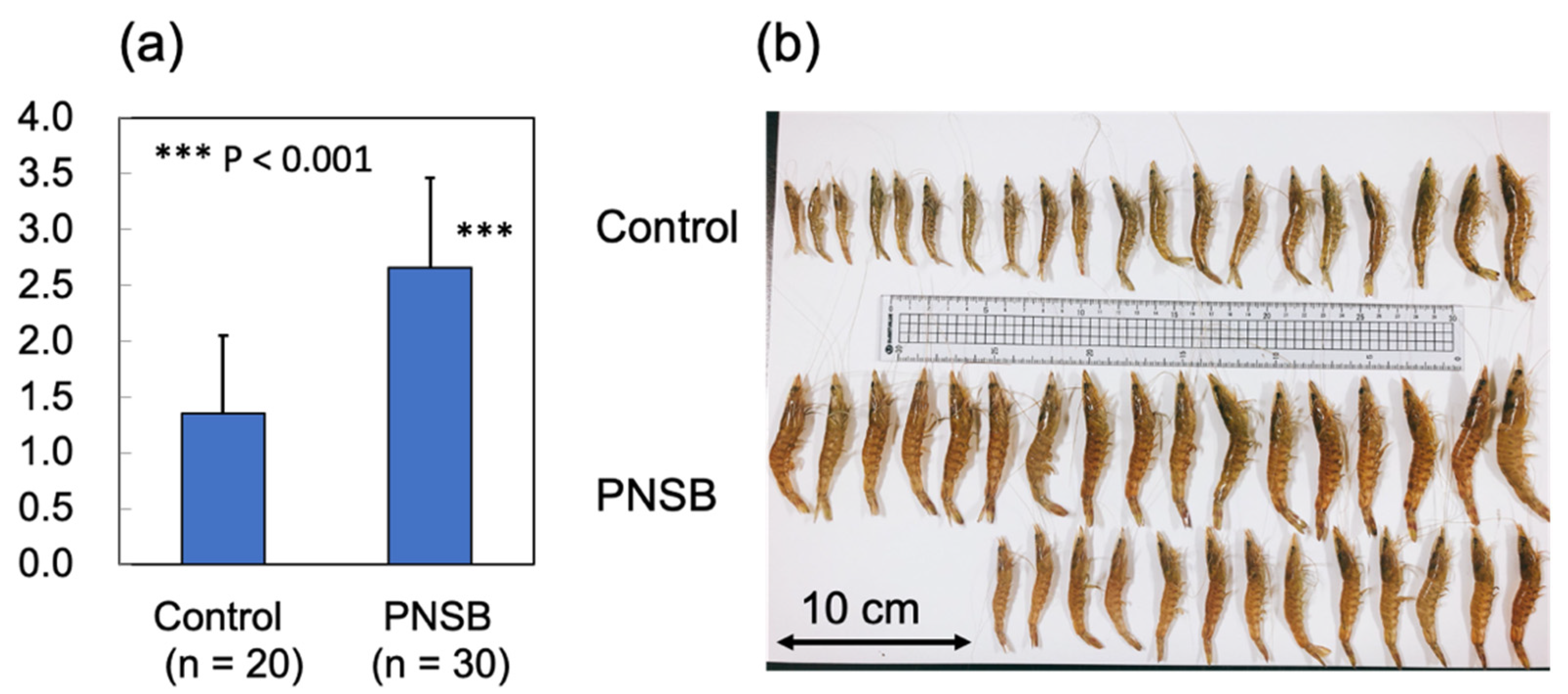
3.2. Outdoor 200-Ton Aquaria Experiments
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thitamadee, S.; Prachumwat, A.; Srisala, J.; Jaroenlak, P.; Salachan, P.V.; Sritunyalucksana, K.; Flegel, T.W.; Itsathitphaisarn, O. Review of current disease threats for cultivated penaeid shrimp in Asia. Aquaculture 2016, 452, 69–87. [Google Scholar] [CrossRef]
- Walker, P.J.; Mohan, C.V. Viral disease emergence in shrimp aquaculture: Origins, impact and the effectiveness of health management strategies. Rev. Aquac. 2009, 1, 125–154. [Google Scholar] [CrossRef]
- Zorriehzahra, M.J.; Delshad, S.T.; Adel, M.; Tiwari, R.; Karthik, K.; Dhama, K.; Lazado, C.C. Probiotics as beneficial microbes in aquaculture: An update on their multiple modes of action: A review. Vet. Q. 2016, 36, 228–241. [Google Scholar] [CrossRef] [Green Version]
- Sakarika, M.; Spanoghe, J.; Sui, Y.; Wambacq, E.; Grunert, O.; Haesaert, G.; Spiller, M.; Vlaeminck, S.E. Purple non-sulphur bacteria and plant production: Benefits for fertilization, stress resistance and the environment. Microb. Biotechnol. 2020, 13, 1336–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.K.; Lur, H.S.; Liu, C. Te From lab to farm: Elucidating the beneficial roles of photosynthetic bacteria in sustainable agriculture. Microorganisms 2021, 9, 2453. [Google Scholar] [CrossRef]
- Chumpol, S.; Kantachote, D.; Rattanachuay, P.; Vuddhakul, V.; Nitoda, T.; Kanzaki, H. In vitro and in vivo selection of probiotic purple nonsulphur bacteria with an ability to inhibit shrimp pathogens: Acute hepatopancreatic necrosis disease-causing Vibrio parahaemolyticus and other vibrios. Aquac. Res. 2017, 48, 3182–3197. [Google Scholar] [CrossRef]
- Chang, B.V.; Liao, C.S.; Chang, Y.T.; Chao, W.L.; Yeh, S.L.; Kuo, D.L.; Yang, C.W. Investigation of a farm-scale multitrophic recirculating aquaculture system with the addition of Rhodovulum sulfidophilum for milkfish (Chanos chanos) coastal aquaculture. Sustainability 2019, 11, 1880. [Google Scholar] [CrossRef] [Green Version]
- Ninawe, A.S.; Selvin, J. Probiotics in shrimp aquaculture: Avenues and challenges. Crit. Rev. Microbiol. 2009, 35, 43–66. [Google Scholar] [CrossRef]
- Sánchez-Ortiz, A.C.; Angulo, C.; Luna-González, A.; Álvarez-Ruiz, P.; Mazón-Suástegui, J.M.; Campa-Córdova, Á.I. Effect of mixed-Bacillus spp isolated from pustulose ark Anadara tuberculosa on growth, survival, viral prevalence and immune-related gene expression in shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2016, 59, 95–102. [Google Scholar] [CrossRef]
- Chen, M.; Chen, X.Q.; Tian, L.X.; Liu, Y.J.; Niu, J. Improvement of growth, intestinal short-chain fatty acids, non-specific immunity and ammonia resistance in Pacific white shrimp (Litopenaeus vannamei) fed dietary water-soluble chitosan and mixed probiotics. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2020, 236, 108791. [Google Scholar] [CrossRef] [PubMed]
- Won, S.; Hamidoghli, A.; Choi, W.; Bae, J.; Jang, W.J.; Lee, S.; Bai, S.C. Evaluation of potential probiotics bacillus subtilis WB60, Pediococcus pentosaceus, and Lactococcus lactis on growth performance, immune response, gut histology and immune-related genes in whiteleg shrimp, Litopenaeus vannamei. Microorganisms 2020, 8, 281. [Google Scholar] [CrossRef] [Green Version]
- Zokaeifar, H.; Balcázar, J.L.; Saad, C.R.; Kamarudin, M.S.; Sijam, K.; Arshad, A.; Nejat, N. Effects of Bacillus subtilis on the growth performance, digestive enzymes, immune gene expression and disease resistance of white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2012, 33, 683–689. [Google Scholar] [CrossRef] [Green Version]
- Interaminense, J.A.; Vogeley, J.L.; Gouveia, C.K.; Portela, R.S.; Oliveira, J.P.; Silva, S.M.B.C.; Coimbra, M.R.M.; Peixoto, S.M.; Soares, R.B.; Buarque, D.S.; et al. Effects of dietary Bacillus subtilis and Shewanella algae in expression profile of immune-related genes from hemolymph of Litopenaeus vannamei challenged with Vibrio parahaemolyticus. Fish Shellfish Immunol. 2019, 86, 253–259. [Google Scholar] [CrossRef]
- Antony, S.P.; Singh, I.S.B.; Sudheer, N.S.; Vrinda, S.; Priyaja, P.; Philip, R. Molecular characterization of a crustin-like antimicrobial peptide in the giant tiger shrimp, Penaeus monodon, and its expression profile in response to various immunostimulants and challenge with WSSV. Immunobiology 2011, 216, 184–194. [Google Scholar] [CrossRef]
- Zheng, X.; Duan, Y.; Dong, H.; Zhang, J. Effects of dietary Lactobacillus plantarum in different treatments on growth performance and immune gene expression of white shrimp Litopenaeus vannamei under normal condition and stress of acute low salinity. Fish Shellfish Immunol. 2017, 62, 195–201. [Google Scholar] [CrossRef]
- Chiu, C.H.; Guu, Y.K.; Liu, C.H.; Pan, T.M.; Cheng, W. Immune responses and gene expression in white shrimp, Litopenaeus vannamei, induced by Lactobacillus plantarum. Fish Shellfish Immunol. 2007, 23, 364–377. [Google Scholar] [CrossRef] [PubMed]
- Huynh, T.G.; Cheng, A.C.; Chi, C.C.; Chiu, K.H.; Liu, C.H. A synbiotic improves the immunity of white shrimp, Litopenaeus vannamei: Metabolomic analysis reveal compelling evidence. Fish Shellfish Immunol. 2018, 79, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Wang, B.; Jiang, K.; Wang, M.; Zhou, S.; Liu, M.; Wang, L. Exploring the influence of the surface proteins on probiotic effects performed by Lactobacillus pentosus HC-2 using transcriptome analysis in Litopenaeus vannamei midgut. Fish Shellfish Immunol. 2019, 87, 853–870. [Google Scholar] [CrossRef] [PubMed]
- Chomwong, S.; Charoensapsri, W.; Amparyup, P.; Tassanakajon, A. Two host gut-derived lactic acid bacteria activate the proPO system and increase resistance to an AHPND-causing strain of Vibrio parahaemolyticus in the shrimp Litopenaeus vannamei. Dev. Comp. Immunol. 2018, 89, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Maeda, M.; Shibata, A.; Biswas, G.; Korenaga, H.; Kono, T.; Itami, T.; Sakai, M. Isolation of lactic acid bacteria from kuruma shrimp (Marsupenaeus japonicus) intestine and assessment of immunomodulatory role of a selected strain as probiotic. Mar. Biotechnol. 2014, 16, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Zhang, J.; Huang, J.; Jiang, S. Effects of dietary Clostridium butyricum on the growth, digestive enzyme activity, antioxidant capacity, and resistance to nitrite stress of Penaeus monodon. Probiotics Antimicrob. Proteins 2019, 11, 938–945. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Tian, X.; Zhao, K.; Jiang, W.; Dong, S. Effect of Clostridium butyricum in different forms on growth performance, disease resistance, expression of genes involved in immune responses and mTOR signaling pathway of Litopenaeus vannamai. Fish Shellfish Immunol. 2019, 87, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Zhang, Y.; Dong, H.; Wang, Y.; Zhang, J. Effect of the dietary probiotic Clostridium butyricum on growth, intestine antioxidant capacity and resistance to high temperature stress in kuruma shrimp Marsupenaeus japonicus. J. Therm. Biol. 2017, 66, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Zhang, Y.; Dong, H.; Wang, Y.; Zheng, X.; Zhang, J. Effect of dietary Clostridium butyricum on growth, intestine health status and resistance to ammonia stress in Pacific white shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2017, 65, 25–33. [Google Scholar] [CrossRef]
- Duan, Y.; Wang, Y.; Dong, H.; Ding, X.; Liu, Q.; Li, H.; Zhang, J.; Xiong, D. Changes in the intestine microbial, digestive, and immune-related genes of Litopenaeus vannamei in response to dietary probiotic Clostridium butyricum supplementation. Front. Microbiol. 2018, 9, 2191. [Google Scholar] [CrossRef] [Green Version]
- Sunish, K.S.; Biji, M.; Rosamma, P.; Sudheer, N.S.; Sreedharan, K.; Mohandas, A.; Singh, I.S.B. Marine actinomycetes Nocardiopsis alba MCCB 110 has immunomodulatory property in the tiger shrimp Penaeus monodon. Fish Shellfish Immunol. 2020, 102, 125–132. [Google Scholar] [CrossRef]
- Licona-Jain, A.; Campa-Córdova, Á.; Luna-González, A.; Racotta, I.S.; Tello, M.; Angulo, C. Dietary supplementation of marine yeast Yarrowia lipolytica modulates immune response in Litopenaeus vannamei. Fish Shellfish Immunol. 2020, 105, 469–476. [Google Scholar] [CrossRef]
- Dwyer, D.J.; Kohanski, M.A.; Collins, J.J. Role of reactive oxygen species in antibiotic action and resistance. Curr. Opin. Microbiol. 2009, 12, 482–489. [Google Scholar] [CrossRef] [Green Version]
- Tassanakajon, A.; Rimphanitchayakit, V.; Visetnan, S.; Amparyup, P.; Somboonwiwat, K.; Charoensapsri, W.; Tang, S. Shrimp humoral responses against pathogens: Antimicrobial peptides and melanization. Dev. Comp. Immunol. 2018, 80, 81–93. [Google Scholar] [CrossRef]
- Wang, P.H.; Huang, T.; Zhang, X.; He, J.G. Antiviral defense in shrimp: From innate immunity to viral infection. Antiviral Res. 2014, 108, 129–141. [Google Scholar] [CrossRef]
- Li, F.; Xiang, J. Signaling pathways regulating innate immune responses in shrimp. Fish Shellfish Immunol. 2013, 34, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, S.; Kerkar, S.; D’Costa, A.; Costa, M.; Mishra, A.; Shyama, S.K.; Das, K.R. Immuno-stimulatory effect and toxicology studies of salt pan bacteria as probiotics to combat shrimp diseases in aquaculture. Fish Shellfish Immunol. 2021, 113, 69–78. [Google Scholar] [CrossRef]
- Liu, R.; Wu, W.; Xu, X.; Wang, Y.; Yu, T.; Wang, J.; Zheng, Q.; Changru, X.; Wu, P. Rhodopseudomonas palustris in effluent enhances the disease resistance, TOR and NF-κB signalling pathway, intestinal microbiota and aquaculture water quality of Pelteobagrus vachelli. Aquac. Res. 2020, 51, 3959–3971. [Google Scholar] [CrossRef]
- Qi, Z.; Zhang, X.H.; Boon, N.; Bossier, P. Probiotics in aquaculture of China—Current state, problems and prospect. Aquaculture 2009, 290, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Maki, T. Applications of Rhodobacter capsulata in agriculture, stock raising, environmental technologies, and aquaculture. Seibutsu-kogaku kaishi 2011, 89, 113–116. [Google Scholar]
- Koga, A.; Yamauchi, N.; Imamura, M.; Urata, M.; Kurayama, T.; Iwai, R.; Hayashi, S.; Yamamoto, S.; Miyasaka, H. pufC gene targeted PCR primers for Identification and classification of marine photosynthetic bacterium Rhodovulum sulfidophilum. Acta Sci. Microbiol. 2021, 4, 69–75. [Google Scholar] [CrossRef]
- Sangkharak, K.; Prasertsan, P. Nutrient optimization for production of polyhydroxybutyrate from halotolerant photosynthetic bacteria cultivated under aerobic-dark condition. Electron. J. Biotechnol. 2008, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Hiromichi Nagasawa The crustacean cuticle: Structure, composition and mineralization. Front. Biosci. 2012, E4, 711–720. [CrossRef]
- Watanabe, T.; Persson, P.; Endo, H.; Fukuda, I.; Furukawa, K.; Kono, M. Identification of a novel cuticular protein in the kuruma prawn Penaeus japonicus. Fish. Sci. 2006, 72, 452–454. [Google Scholar] [CrossRef]
- Ikeya, T.; Persson, P.; Kono, M.; Watanabe, T. The DD5 gene of the decapod crustacean Penaeus japonicus encodes a putative exoskeletal protein with a novel tandem repeat structure. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2001, 128, 379–388. [Google Scholar] [CrossRef]
- Watanabe, T.; Persson, P.; Endo, H.; Kono, M. Molecular analysis of two genes, DD9A and B, which are expressed during the postmolt stage in the decapod crustacean Penaeus japonicus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2000, 125, 127–136. [Google Scholar] [CrossRef]
- Endo, H.; Persson, P.; Watanabe, T. Molecular cloning of the crustacean DD4 cDNA encoding a Ca2+-binding protein. Biochem. Biophys. Res. Commun. 2000, 276, 286–291. [Google Scholar] [CrossRef]
- Ahearn, G.A.; Mandal, P.K.; Mandal, A. Calcium regulation in crustaceans during the molt cycle: A review and update. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2004, 137, 247–257. [Google Scholar] [CrossRef]
- Wade, N.M.; Tollenaere, A.; Hall, M.R.; Degnan, B.M. Evolution of a novel carotenoid-binding protein responsible for crustacean shell color. Mol. Biol. Evol. 2009, 26, 1851–1864. [Google Scholar] [CrossRef] [Green Version]
- Koyama, H.; Akolkar, D.B.; Piyapattanakorn, S.; Watabe, S. Cloning, expression, and localization of two types of fast skeletal myosin heavy chain genes from black tiger and Pacific white shrimps. J. Exp. Zool. A. Ecol. Genet. Physiol. 2012, 317, 608–621. [Google Scholar] [CrossRef]
- Junprung, W.; Supungul, P.; Tassanakajon, A. Litopenaeus vannamei heat shock protein 70 (LvHSP70) enhances resistance to a strain of Vibrio parahaemolyticus, which can cause acute hepatopancreatic necrosis disease (AHPND), by activating shrimp immunity. Dev. Comp. Immunol. 2019, 90, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.R.; Xu, Y.H.; Jiang, H.S.; Xu, S.; Zhao, X.F.; Wang, J.X. Antibacterial activity of serine protease inhibitor 1 from kuruma shrimp Marsupenaeus japonicus. Dev. Comp. Immunol. 2014, 44, 261–269. [Google Scholar] [CrossRef]
- Wang, Z.; Sun, B.; Zhu, F. Molecular characterization of diphthamide biosynthesis protein 7 in Marsupenaeus japonicus and its role in white spot syndrome virus infection. Fish Shellfish Immunol. 2018, 75, 8–16. [Google Scholar] [CrossRef]
- Santos, C.A.; Andrade, S.C.S.; Teixeira, A.K.; Farias, F.; Guerrelhas, A.C.; Rocha, J.L.; Freitas, P.D. Transcriptome differential expression analysis reveals the activated genes in Litopenaeus vannamei shrimp families of superior growth performance. Aquaculture 2021, 531, 735871. [Google Scholar] [CrossRef]
- Toledo, A.; Frizzo, L.; Signorini, M.; Bossier, P.; Arenal, A. Impact of probiotics on growth performance and shrimp survival: A meta-analysis. Aquaculture 2019, 500, 196–205. [Google Scholar] [CrossRef]
- Wang, Y.; Gu, Q. Effect of probiotics on white shrimp (Penaeus vannamei) growth performance and immune response. Mar. Biol. Res. 2010, 6, 327–332. [Google Scholar] [CrossRef]
- Chai, P.-C.; Song, X.-L.; Chen, G.-F.; Xu, H.; Huang, J. Dietary supplementation of probiotic Bacillus PC465 isolated from the gut of Fenneropenaeus chinensis improves the health status and resistance of Litopenaeus vannamei against white spot syndrome virus. Fish Shellfish Immunol. 2016, 54, 602–611. [Google Scholar] [CrossRef] [PubMed]
- Miandare, H.K.; Yarahmadi, P.; Abbasian, M. Immune related transcriptional responses and performance of Litopenaeus vannamei post-larvae fed on dietary probiotic PrimaLac®. Fish Shellfish Immunol. 2016, 55, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.Q.; Xue, M.; Liang, H.F.; Wu, Y.; Li, X. Beneficial effects of Ectothiorhodospira shaposhnikovii WF on larval cultivation of Litopenaeus vannamei. Benef. Microbes 2015, 6, 525–533. [Google Scholar] [CrossRef]
- Jamali, H.; Imani, A.; Abdollahi, D.; Roozbehfar, R.; Isari, A. Use of probiotic Bacillus spp. in rotifer (Brachionus plicatilis) and artemia (Artemia urmiana) enrichment: Effects on growth and survival of Pacific white shrimp, Litopenaeus vannamei, larvae. Probiotics Antimicrob. Proteins 2015, 7, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.F.; Chiu, C.H.; Shiu, Y.L.; Cheng, W.; Liu, C.H. Effects of the probiotic, Bacillus subtilis E20, on the survival, development, stress tolerance, and immune status of white shrimp, Litopenaeus vannamei larvae. Fish Shellfish Immunol. 2010, 28, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Verhasselt, V.; Buelens, C.; Willems, F.; De Groote, D.; Haeffner-Cavaillon, N.; Goldman, M. Bacterial lipopolysaccharide stimulates the production of cytokines and the expression of costimulatory molecules by human peripheral blood dendritic cells: Evidence for a soluble CD14-dependent pathway. J. Immunol. 1997, 158, 2919–2925. [Google Scholar] [PubMed]
- Takahashi, Y.; Kondo, M.; Itami, T.; Honda, T.; Inagawa, H.; Nishizawa, T.; Soma, G.I.; Yokomizo, Y. Enhancement of disease resistance against penaeid acute viraemia and induction of virus-inactivating activity in haemolymph of kuruma shrimp, Penaeus japonicus, by oral administration of Pantoea agglomerans lipopolysaccharide (LPS). Fish Shellfish Immunol. 2000, 10, 555–558. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Average Body Weight (g) | Final Number of Shrimp 1 | Survival Rate (%) | Total Body Weight (g) | Total Feed Amount (kg) | FCE 2 | |
|---|---|---|---|---|---|---|
| Control | 7.73 | 589 | 58.9 | 4552 | 7995 | 1.76 |
| R. sulfidophilum-fed | 11.9 *** | 672 *** | 67.2 | 8024 | 12,650 | 1.58 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koga, A.; Goto, M.; Hayashi, S.; Yamamoto, S.; Miyasaka, H. Probiotic Effects of a Marine Purple Non-Sulfur Bacterium, Rhodovulum sulfidophilum KKMI01, on Kuruma Shrimp (Marsupenaeus japonicus). Microorganisms 2022, 10, 244. https://doi.org/10.3390/microorganisms10020244
Koga A, Goto M, Hayashi S, Yamamoto S, Miyasaka H. Probiotic Effects of a Marine Purple Non-Sulfur Bacterium, Rhodovulum sulfidophilum KKMI01, on Kuruma Shrimp (Marsupenaeus japonicus). Microorganisms. 2022; 10(2):244. https://doi.org/10.3390/microorganisms10020244
Chicago/Turabian StyleKoga, Aoi, Midori Goto, Shuhei Hayashi, Shinjiro Yamamoto, and Hitoshi Miyasaka. 2022. "Probiotic Effects of a Marine Purple Non-Sulfur Bacterium, Rhodovulum sulfidophilum KKMI01, on Kuruma Shrimp (Marsupenaeus japonicus)" Microorganisms 10, no. 2: 244. https://doi.org/10.3390/microorganisms10020244
APA StyleKoga, A., Goto, M., Hayashi, S., Yamamoto, S., & Miyasaka, H. (2022). Probiotic Effects of a Marine Purple Non-Sulfur Bacterium, Rhodovulum sulfidophilum KKMI01, on Kuruma Shrimp (Marsupenaeus japonicus). Microorganisms, 10(2), 244. https://doi.org/10.3390/microorganisms10020244