Diversity of the Seedborne Fungi and Pathogenicity of Fusarium Species Associated with Intercropped Soybean

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
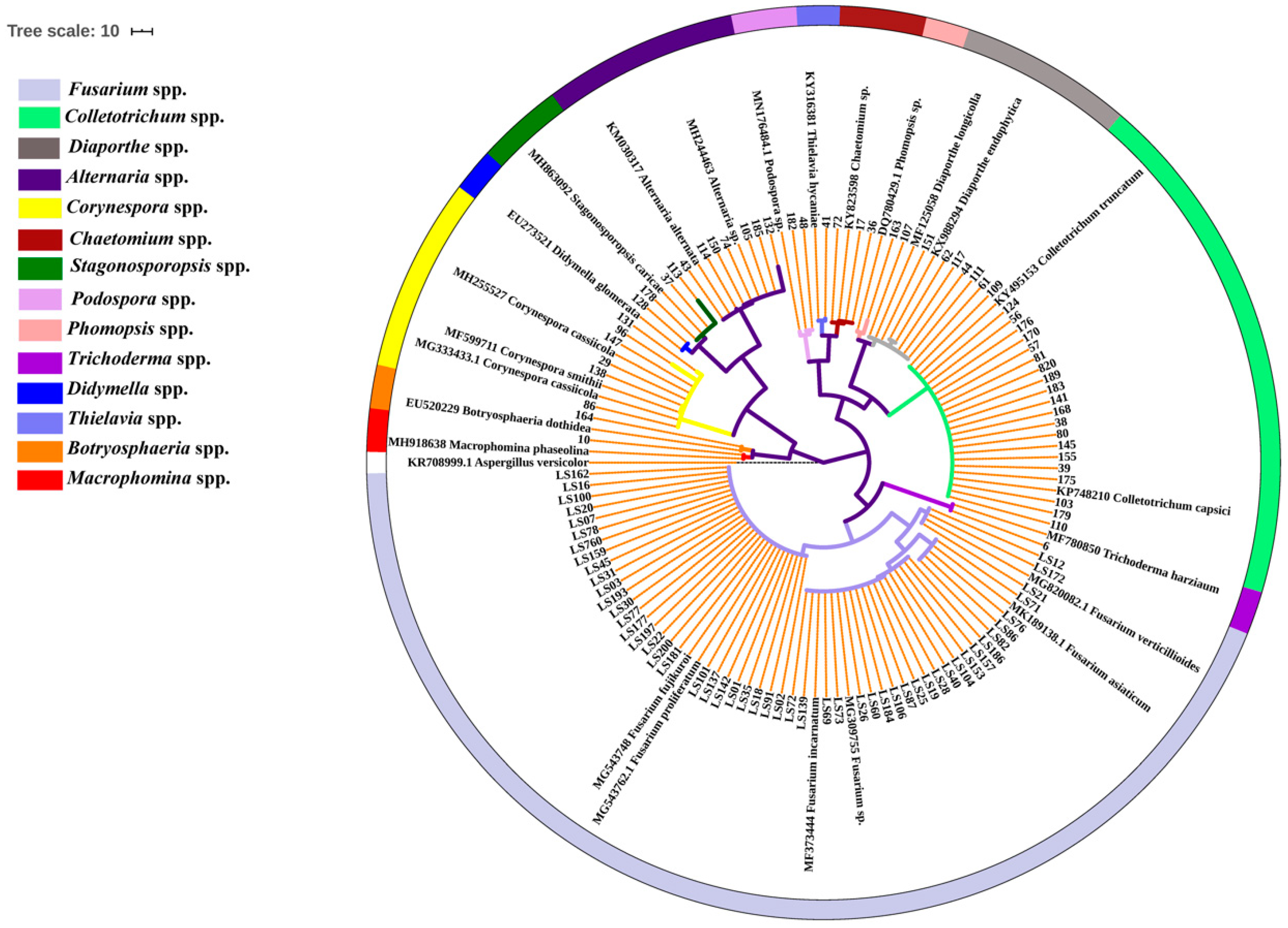
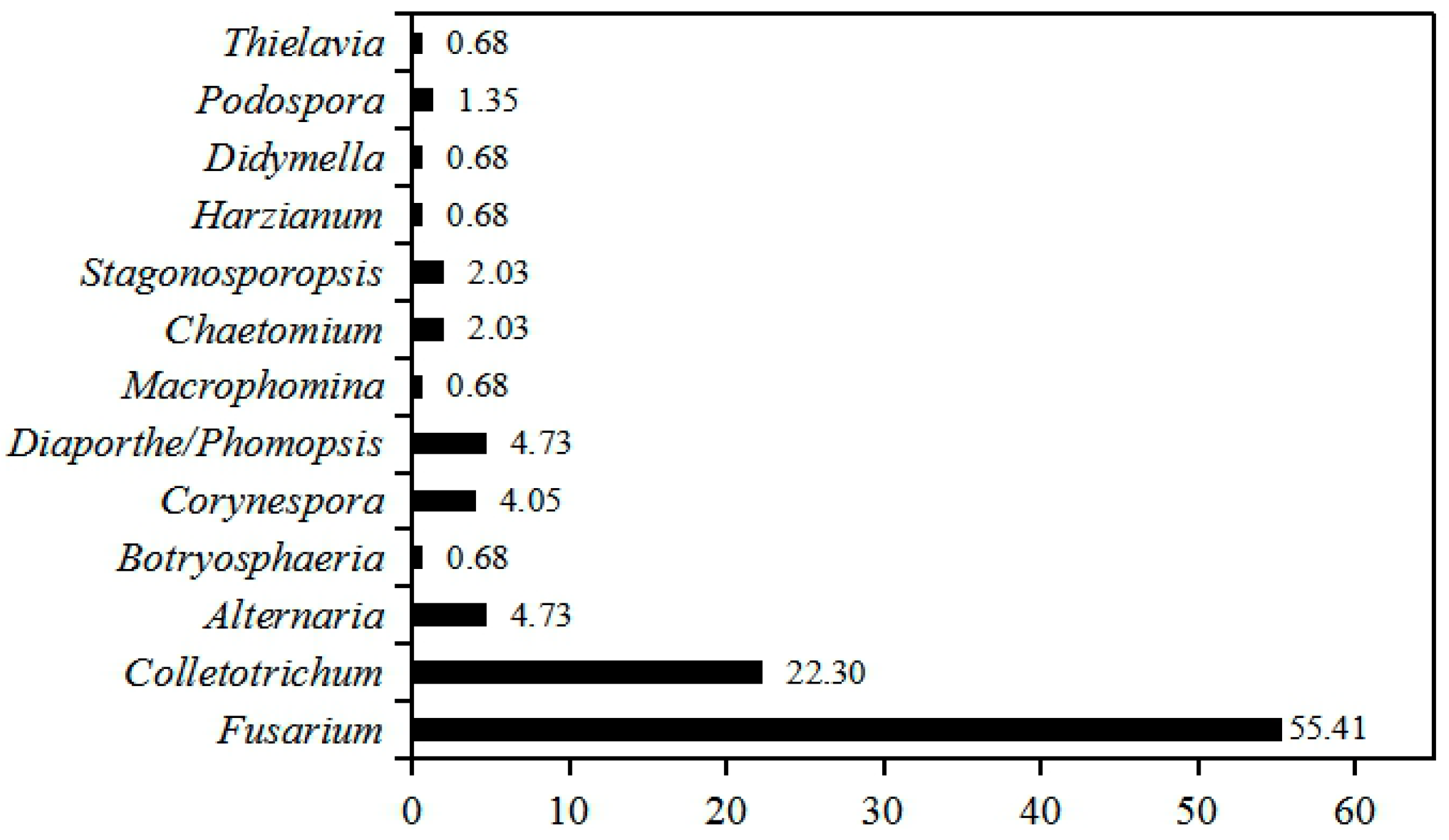
2.1. Population Identification of the Seedborne Fungi Associated with Intercropped Soybean
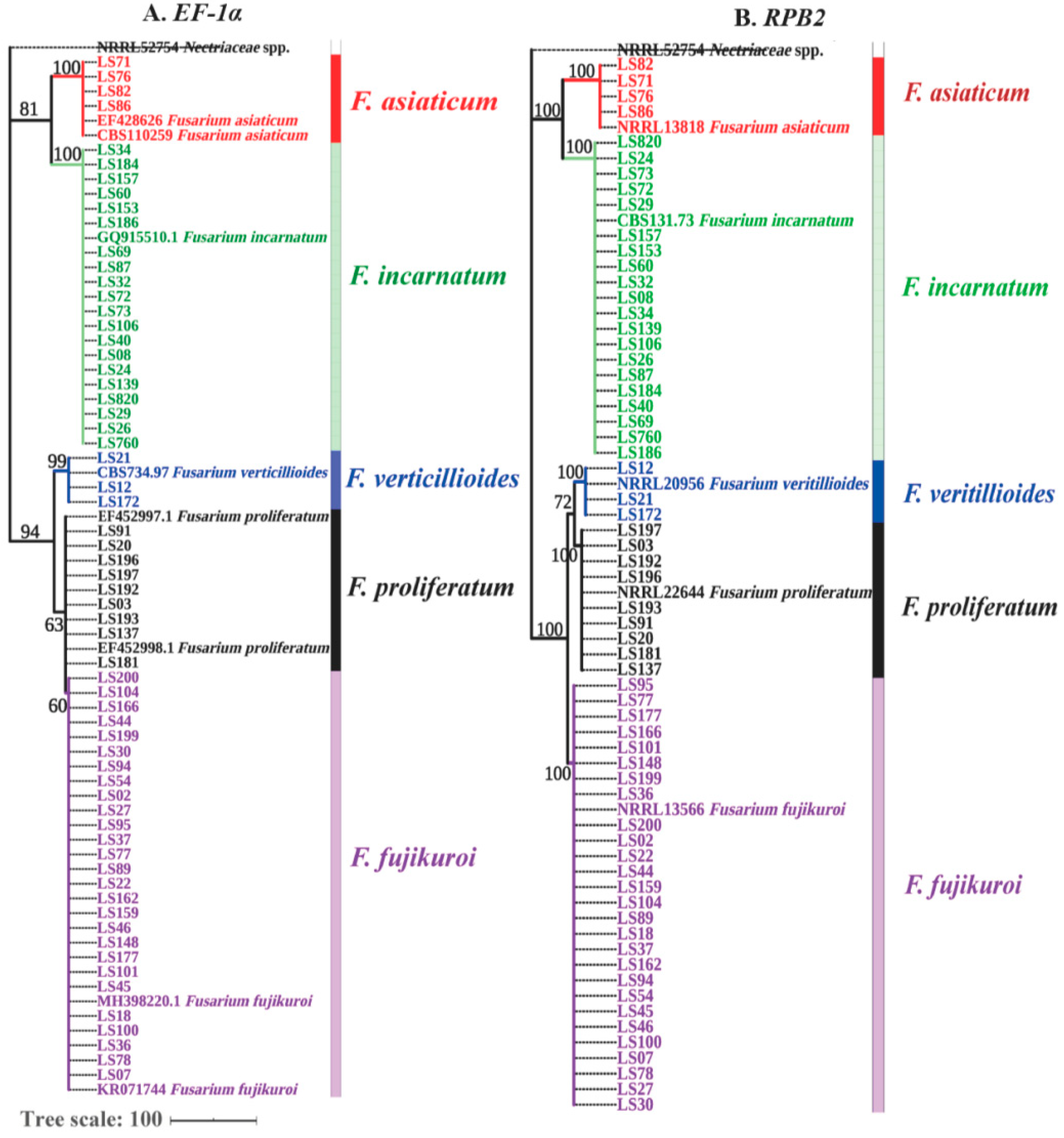
2.2. Identification of Fusarium Species Associated with Soybean Seeds Based on EF-1α and RPB2 Genes
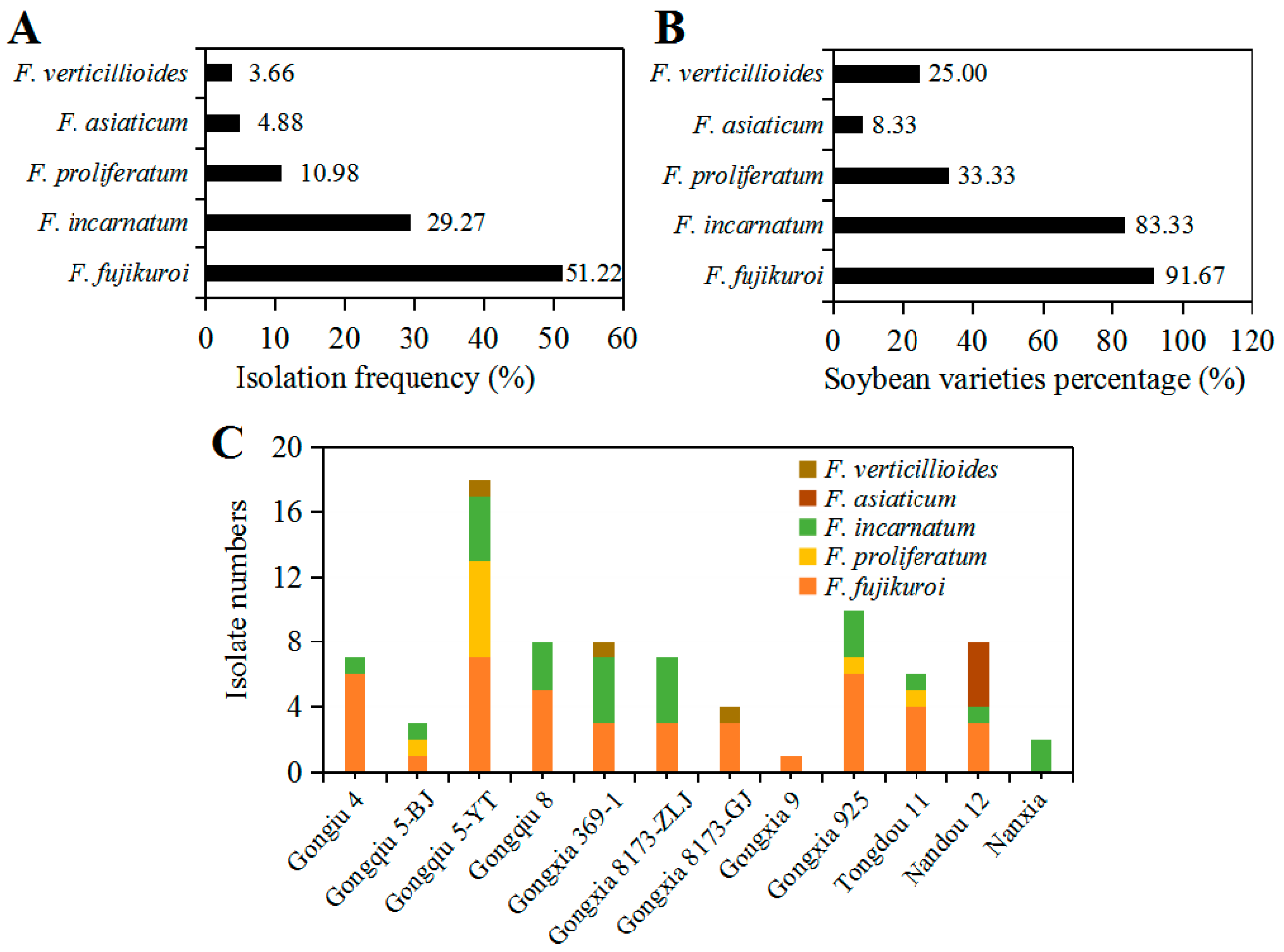
2.3. Isolation Frequency of Fusarium Species and Its Diversity Correlated with Soybean Varieties
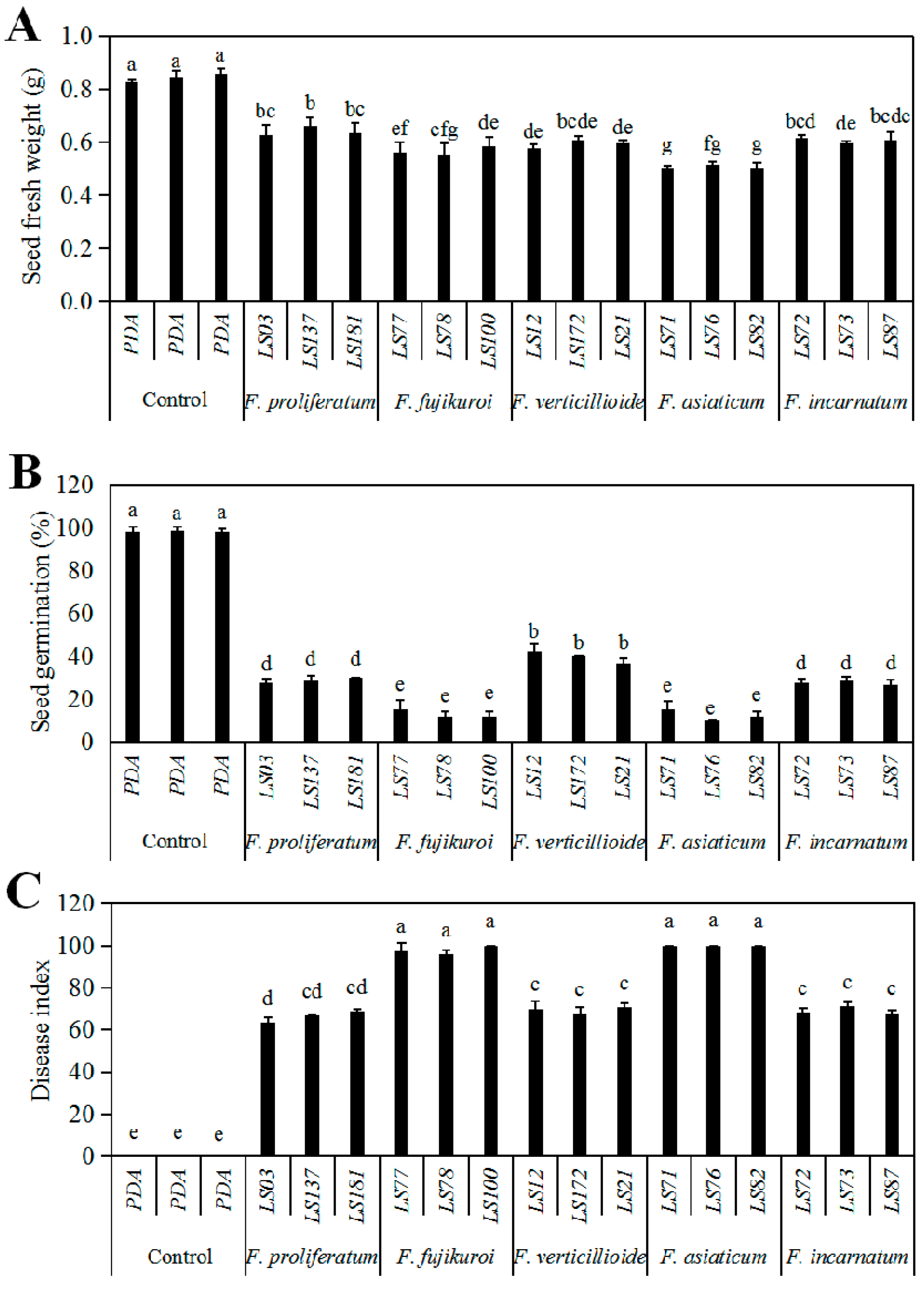
2.4. Pathogenicity of the Seedborne Fusarium Species Associated with Intercropped Soybean
3. Discussion
4. Materials and Methods
4.1. Samples Collection and Isolation of Seedborne Fungi
4.2. PCR Amplification of rDNA ITS, EF-1α and RPB2 Sequences
4.3. Phylogenetic Analysis
4.4. Pathogenicity Assay of Fusarium Species on Soybean Seeds
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Li, S.; Hartman, G.L.; Boykin, D.L. Aggressiveness of Phomopsis longicolla and other Phomopsis spp. on soybean. Plant Dis. 2010, 94, 1035–1040. [Google Scholar] [CrossRef]
- Hartman, G.L. Worldwide importance of soybean pathogens and pests. In Compendium of Soybean Diseases and Pests, 5th ed.; The American Phytopathological Society: St Paul, MN. USA, 2015; pp. 4–5. [Google Scholar] [CrossRef]
- Liu, J.; Deng, J.; Zhang, K.; Wu, H.; Yang, C.; Zhang, X.; Du, J.; Shu, K.; Yang, W. Pod mildew on soybeans can mitigate the damage to the seed arising from field mold at harvest time. J. Agric. Food Chem. 2016, 64, 9135–9142. [Google Scholar] [CrossRef]
- Naeem, M.; Li, H.; Yan, L.; Raza, M.A.; Gong, G.; Chen, H.; Yang, C.; Zhang, M.; Shang, J.; Liu, T.; et al. Characterization and pathogenicity of Fusarium species associated with soybean pods in maize/soybean strip intercropping. Pathogens 2019, 8, 245. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.K.; Sinclair, J.B. Principles of Seed Pathology; CRC Press: Boca Raton, FL, USA, 1996. [Google Scholar]
- Machado, J.C.; Langerak, C.J.; Jaccoud-Filho, D.S. Seed-Borne Fungi: A Contribution to Routine Seed Health Analysis; ISTA/UFLA: Bassersdorf, Switzerland, 2002. [Google Scholar]
- Ellis, M.A.; Galvez, E.G.E. Seed pathology. In Bean Production Problems: Disease, Insect Soil and Climatic Constraints of Phaseolus Vulgaris; Howard, F., Gálvez, E., Guillermo, E., Eds.; Centro Internacional de Agricultura Tropical (CIAT): Cali, CO, USA, 1980; pp. 301–314. [Google Scholar]
- Park, J.S.; Lee, K.R.; Kim, J.C.; Lim, S.H.; Seo, J.A.; Lee, Y.W. A Hemorrhagic factor (apicidin) produced by toxic Fusarium isolates from soybean seeds. Appl. Environ. Microb. 1999, 65, 126–130. [Google Scholar] [CrossRef]
- Barros, G.; Zanon, M.A.; Abod, A.; Oviedo, M.; Ramirez, M.; Reynoso, M.; Torres, A.; Chulze, S. Natural deoxynivalenol occurrence and genotype and chemotype determination of a field population of the Fusarium graminearum complex associated with soybean in Argentina. Food Addit. Contam. Part A 2012, 29, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Garcia, D.; Barros, G.; Chulze, S.; Ramos, A.J.; Sanchis, V.; Marín, S. Impact of cycling temperatures on Fusarium verticillioides and Fusarium graminearum growth and mycotoxins production in soybean. J. Sci. Food Agric. 2012, 92, 2952–2959. [Google Scholar] [CrossRef]
- Fernandez, M.R.; Fernandes, J.M.C. Survival of wheat pathogens in wheat and soybean residues under conservation tillage systems in southern and central Brazil. Can. J. Plant Pathol. 1990, 12, 289–294. [Google Scholar] [CrossRef]
- Wrather, J.; Shannon, J.; Stevens, W.; Sleper, D.; Arelli, A. Soybean cultivar and foliar fungicide effects on Phomopsis sp. seed infection. Plant Dis. 2004, 88, 721–723. [Google Scholar] [CrossRef] [PubMed]
- Wrather, J.A.; Shannon, J.G.; Mengistu, A. Impact of soybean planting date on soil population density of Macrophomina phaseolina. Plant Health Prog. 2007. [Google Scholar] [CrossRef]
- Li, S.; Chen, P. Resistance to Phomopsis seed decay in soybean. ISRN Agron. 2013, 738379. [Google Scholar] [CrossRef]
- Li, S.; Rupe, J.; Chen, P.; Shannon, G.; Wrather, A.; Boykin, D. Evaluation of diverse soybean germplasm for resistance to Phomopsis seed decay. Plant Dis. 2015, 99, 1517–1525. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Smith, J. Evaluation of Soybean Breeding Lines for Resistance to Phomopsis Seed Decay in Stoneville, Mississippi; Plant Disease Management Reports 10:FC045; Crop Genetics Research: Stoneville, MS, USA, 2016. [Google Scholar]
- Miller, W.A.; Roy, K.W. Mycoflora of soybean leaves, pods, and seeds in Mississippi. Can. J. Bot. 1982, 60, 2716–2723. [Google Scholar] [CrossRef]
- Hobbs, T.W.; Schmitthenner, A.; Kuter, G.A. A new Phomopsis species from soybean. Mycologia 1985, 77, 535–544. [Google Scholar] [CrossRef]
- Zhang, A.W.; Riccioni, L.; Pedersen, W.L.; Kollipara, K.P.; Hartman, G.L. Molecular identification and phylogenetic grouping of Diaporthe phaseolorum and Phomopsis longicolla isolates from soybean. Phytopathology 1998, 88, 1306–1314. [Google Scholar] [CrossRef]
- Ellis, M.L.; Díaz Arias, M.M.; Leandro, L.F. First report of Fusarium armeniacum causing seed rot and root rot on soybean (Glycine max) in the United States. Plant Dis. 2013, 97, 1557–1562. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.D.; Dias-Neto, J.J.; Santos, M.D.M.; Formento, A.N.; Bizerra, L.V.; Fonseca, M.E.N.; Boiteux, L.S.; Café-Filho, A.C. Current status of soybean anthracnose associated with Colletotrichum truncatum in Brazil and Argentina. Plants 2019, 8, 459. [Google Scholar] [CrossRef]
- Petrović, K.; Riccioni, L.; Vidić, M.; Đorđević, V.; Balešević-Tubić, S.; Đukić, V.; Miladinov, Z. First report of Diaporthe novem, D. foeniculina, and D. rudis associated with soybean seed decay in Serbia. Plant Dis. 2016, 100, 2324. [Google Scholar] [CrossRef]
- Broders, K.D.; Lipps, P.E.; Paul, P.A.; Dorrance, A.E. Evaluation of Fusarium graminearum associated with corn and soybean seed and seedling disease in Ohio. Plant Dis. 2007, 91, 1155–1160. [Google Scholar] [CrossRef]
- Díaz Arias, M.M.; Leandro, L.; Munkvold, G.P. Aggressiveness of Fusarium species and impact of root infection on growth and yield of soybean. Phytopathology 2013, 103, 822–832. [Google Scholar] [CrossRef]
- Pedrozo, R.; Little, C.R. Fusarium verticillioides inoculum potential influences soybean seed quality. Eur. J. Plant Pathol. 2017, 148, 749–754. [Google Scholar] [CrossRef]
- Chang, K.F.; Hwang, S.F.; Conner, R.L.; Gossen, B.D. First report of Fusarium proliferatum causing root rot in soybean (Glycine max L.) in Canada. Crop Prot. 2015, 67, 52–58. [Google Scholar] [CrossRef]
- Chang, X.L.; Dai, H.; Wang, D.P.; Zhou, H.H.; He, W.Q.; Fu, Y.; Ibrahim, F.; Zhou, Y.; Gong, G.S.; Shang, J.; et al. Identification of Fusarium species associated with soybean root rot in Sichuan Province, China. Eur. J. Plant Pathol. 2018, 5, 1–15. [Google Scholar] [CrossRef]
- Rupe, J.C.; Ferriss, R.S. Effects of pod moisture on soybean seed infection by Phomopsis sp. Phytopathology 1986, 76, 273–277. [Google Scholar] [CrossRef]
- Balducchi, A.J.; McGee, D.C. Environmental factors influencing infection of soybean seeds by Phomopsis and Diaporthe species during seed maturation. Plant Dis. 1987, 71, 209–212. [Google Scholar] [CrossRef]
- Yang, F.; Wang, X.; Liao, D.; Lu, F.; Gao, R.; Liu, W.; Yong, T.; Wu, X.; Du, J.; Liu, J. Yield response to different planting geometries in maize-soybean relay strip intercropping systems. Agron. J. 2015, 107, 296–304. [Google Scholar] [CrossRef]
- Du, J.; Han, T.; Gai, J.; Yong, T.; Sun, X.; Wang, X.; Yang, F.; Liu, J.; Shu, K.; Liu, W. Maize-soybean strip intercropping: Achieved a balance between high productivity and sustainability. J. Integr. Agric. 2018, 17, 747–754. [Google Scholar] [CrossRef]
- Su, B.; Liu, X.; Cui, L.; Xiang, B.; Yang, W. Suppression of weeds and increases in food production in higher crop diversity planting arrangements: A case study of relay intercropping. Crop Sci. 2018, 58, 1729–1739. [Google Scholar] [CrossRef]
- Munkvold, G.P. Seed pathology progress in academia and industry. Ann. Rev. Phytopathol. 2009, 47, 285–311. [Google Scholar] [CrossRef]
- Escamilla, D.; Rosso, M.L.; Zhang, B. Identification of fungi associated with soybeans and effective seed disinfection treatments. Food Sci. Nutr. 2019, 7, 3194–3205. [Google Scholar] [CrossRef]
- Shovan, L.R.; Bhuiyan, M.K.A.; Sultana, N.; Begum, J.A.; Pervenz, Z. Prevalence of fungi associated with soybean seeds and pathogencity tests of the major seed-borne pathogens. Int. J. Sustain. Crop Prod. 2008, 3, 24–33. [Google Scholar]
- Wei, S.; Zhu, X.; Chen, L.; Wang, Y.; Duan, Y. Seed-borne fungi detection in different soybean cultivars. Soybean Sci. 2014, 33, 890–895. [Google Scholar] [CrossRef]
- Chiotta, M.L.; Alaniz Zanon, M.S.; Palazzini, J.M.; Scandiani, M.M.; Formento, A.N.; Barros, G.G.; Chulze, S.N. Pathogenicity of Fusarium graminearum and F. meridionale on soybean pod blight and trichothecene accumulation. Plant Pathol. 2016, 65, 1492–1497. [Google Scholar] [CrossRef]
- Sharma, S.K.; Gupta, G.K.; Ramteke, R. Colletotrichum truncatum [(Schw.) Andrus & W.D. Moore], the causal agent of anthracnose of soybean [Glycine max (L.) Merrill]—A review. Soybean Res. 2011, 9, 31–52. [Google Scholar]
- Broggi, L.E.; González, H.H.L.; Resnik, S.L.; Pacin, A. Alternaria alternate prevalence in cereal grains and soybean seeds from Entre Rios, Argentina. Rev. Iberoam. Micol. 2007, 24, 47. [Google Scholar] [CrossRef]
- Pedrozo, R.; Fenoglio, J.J.; Little, C.R. The interesting case of soybean seedborne Fusarium spp.: From identify to pathogenicy. In Proceedings of the Annual Meeting of American Phytopathological Society, Pasadena, CA, USA, 1–5 August 2015; Volume 105, p. 109. [Google Scholar] [CrossRef]
- Pedrozo, R.; Fenoglio, J.J.; Little, C.R. First report of seedborne Fusarium fujikuroi and its potential to cause pre- and post- emergent damping-off on soybean (Glycine max) in the United States. Plant Dis. 2015, 99, 1865. [Google Scholar] [CrossRef]
- Leslie, J.F.; Pearson, C.A.S.; Nelson, P.E.; Toussoun, T.A. Fusarium spp. from corn, sorghum, and soybean fields in the central and eastern United States. Phytopathology 1990, 80, 343–350. [Google Scholar] [CrossRef]
- Zhang, H.; Van der Lee, T.; Waalwijk, C.; Chen, W.; Xu, J.; Xu, J.; Zhang, Y.; Feng, J. Population analysis of the Fusarium graminearum species complex from wheat in China show a shift to more aggressive isolates. PLoS ONE 2012, 7, e31722. [Google Scholar] [CrossRef]
- Pedrozo, R.; Little, C.R. First report of seedborne Fusarium thapsinum and its pathogenicity on soybean (Glycine max) in the United States. Plant Dis. 2014, 98, 1745. [Google Scholar] [CrossRef]
- Wicklow, D.T.; Bennett, G.A.; Shotwell, O.L. Secondary invasion of soybeans by Fusarium graminearum and resulting mycotoxin contamination. Plant Dis. 1987, 71, 1146. [Google Scholar] [CrossRef]
- Goswami, R.S.; Kistler, H.C. Heading for disaster: Fusarium graminearum on cereal crops. Mol. Plant Pathol. 2004, 5, 515–525. [Google Scholar] [CrossRef]
- Bakker, M.G.; Brown, D.W.; Kely, A.C.; Kim, H.S.; Kurtzman, C.P.; Mccormick, S.P.; O’Donnell, K.L.; Proctor, R.H.; Vaughan, M.M.; Ward, T.J. Fusarium mycotoxins: A trans-disciplinary overviwe. Can. J. Plant Pathol. 2018, 40, 161–171. [Google Scholar] [CrossRef]
- Perincherry, L.; Lalak-Kańczugowska, J.; Stępień, L. Fusarium-produced mycotoxins in plant-pathogen interactions. Toxins 2019, 11, 664. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S. Analysis of phylogenetic relationships by amplification and direct sequencing of ribosomal RNA genes. In PCR Protocols: A Guide to Methods and Applications; Jnnis, M.A., Ed.; Academic Press: New York, NY, USA, 1990; pp. 15–22. [Google Scholar]
- O’Donnell, K.; Ward, T.J.; Robert, V.A.R.; Crous, P.W.; Geiser, D.W.; Kang, S. DNA sequence-based identification of Fusarium: Current status and future directions. Phytoparasitica 2015, 43, 583–595. [Google Scholar] [CrossRef]
- Aoki, T.; O’Donnell, K.; Geiser, D.M. Systematics of key phytopathogenic Fusarium species: Current status and future challenges. J. Gen. Plant Pathol. 2014, 80, 189–201. [Google Scholar] [CrossRef]
- O’Donnell, K.; Rooney, A.P.; Proctor, R.H.; Brown, D.W.; McCormick, S.P.; Ward, T.J.; Frandsen, R.J.N.; Lysøe, E.; Rehner, S.A.; Aoki, T.; et al. Phylogenetic analyses of RPB1 and RPB2 support a middle Cretaceous origin for a clade comprising all agriculturally and medically important fusaria. Fungal Genet. Biol. 2013, 52, 20–31. [Google Scholar] [CrossRef]
- Reeb, V.; Lutzoni, F.; Roux, C. Contribution of RPB2 to multilocus phylogenetic studies of the euascomycetes (Pezizomycotina, Fungi) with special emphasis on the lichen-forming Acarosporaceae and evolution of polyspory. Mol. Phylogenet. Evol. 2004, 32, 1036–1060. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, X.; Li, H.; Naeem, M.; Wu, X.; Yong, T.; Song, C.; Liu, T.; Chen, W.; Yang, W. Diversity of the Seedborne Fungi and Pathogenicity of Fusarium Species Associated with Intercropped Soybean. Pathogens 2020, 9, 531. https://doi.org/10.3390/pathogens9070531
Chang X, Li H, Naeem M, Wu X, Yong T, Song C, Liu T, Chen W, Yang W. Diversity of the Seedborne Fungi and Pathogenicity of Fusarium Species Associated with Intercropped Soybean. Pathogens. 2020; 9(7):531. https://doi.org/10.3390/pathogens9070531
Chicago/Turabian StyleChang, Xiaoli, Hongju Li, Muhammd Naeem, Xiaoling Wu, Taiwen Yong, Chun Song, Taiguo Liu, Wanquan Chen, and Wenyu Yang. 2020. "Diversity of the Seedborne Fungi and Pathogenicity of Fusarium Species Associated with Intercropped Soybean" Pathogens 9, no. 7: 531. https://doi.org/10.3390/pathogens9070531
APA StyleChang, X., Li, H., Naeem, M., Wu, X., Yong, T., Song, C., Liu, T., Chen, W., & Yang, W. (2020). Diversity of the Seedborne Fungi and Pathogenicity of Fusarium Species Associated with Intercropped Soybean. Pathogens, 9(7), 531. https://doi.org/10.3390/pathogens9070531