
Extracellular Vesicle Release from Immune Cells in Cutaneous Leishmaniasis: Modulation by Leishmania (V.) braziliensis and Reversal by Antimonial Therapy

 , ,
, ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Cohorts
2.2. Reagents
2.3. Removal of Endogenous Extracellular Vesicles from RPMI 1640 Medium and Fetal Bovine Serum (FBS)
2.4. Blood Collection
2.5. Peripheral Blood Mononuclear Cell Cryopreservation Protocol
2.6. Preparation of L. (V.) braziliensis Promastigotes Extract
2.7. Lymphocyte Proliferation Assay and Harvesting of Extracellular Vesicles from PBMC Culture Supernatants
2.8. EV Labeling Protocol for Nano-Flow Cytometry
2.9. Nano-Flow Cytometry Analysis of Extracellular Vesicles
2.10. Data Analysis and Statistics
3. Results
3.1. Optimization of Extracellular Vesicle Detection in Lymphocyte Proliferative Assay
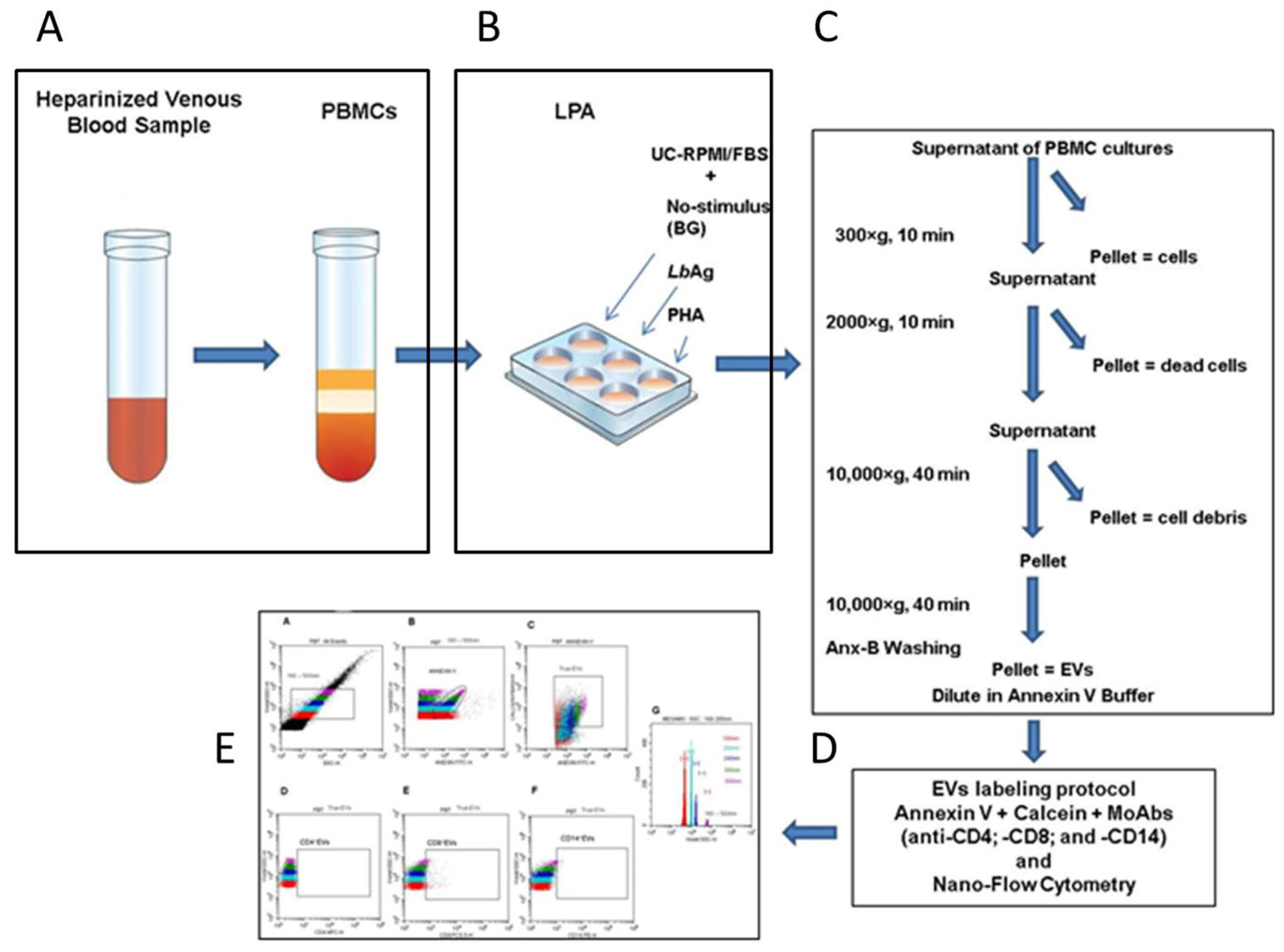
3.2. Definition of a Nano-Flow Cytometry Gating Strategy for EV Quantification and Phenotyping in PBMC Cultures from CL Patients and Healthy Individuals
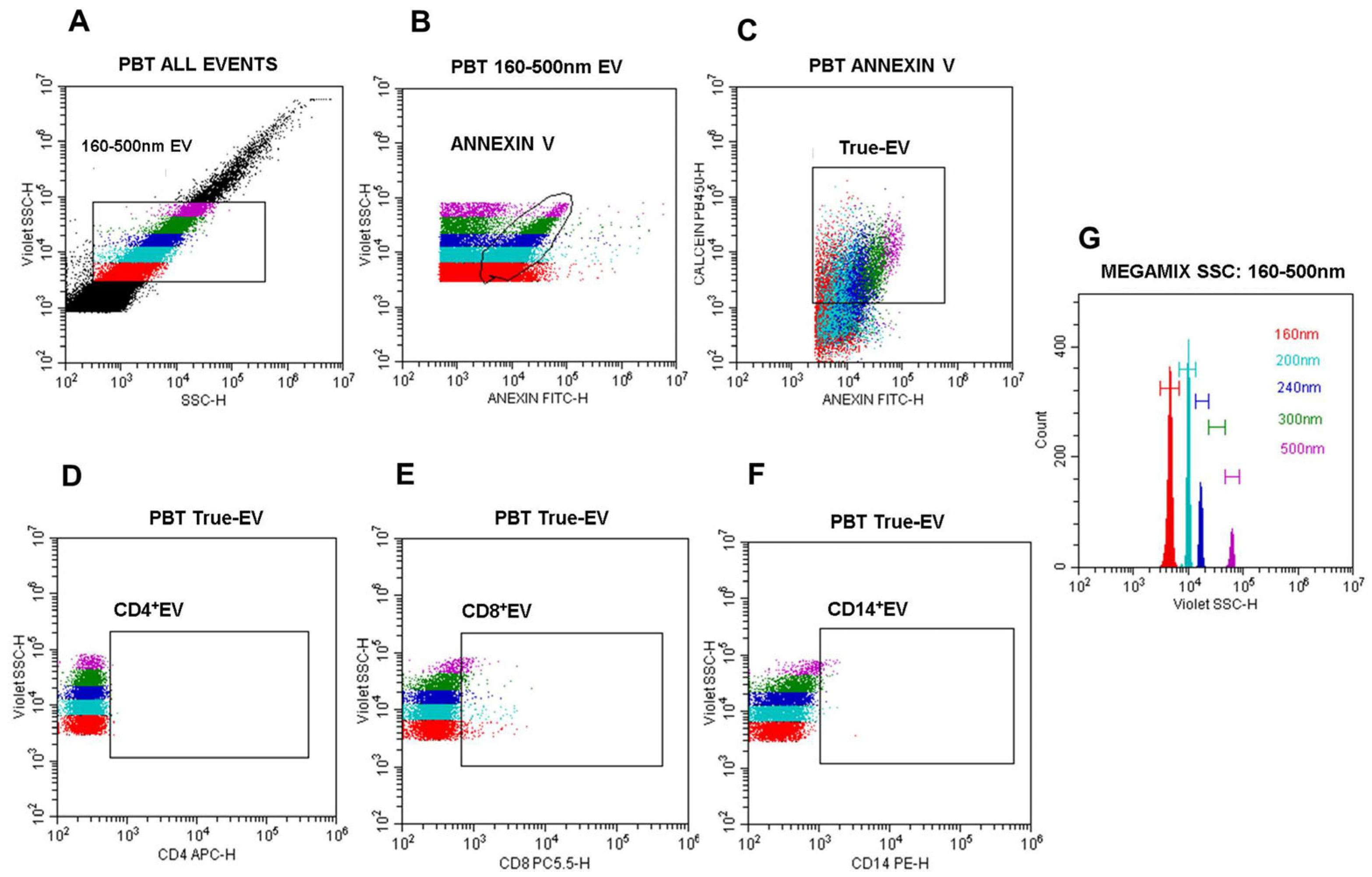
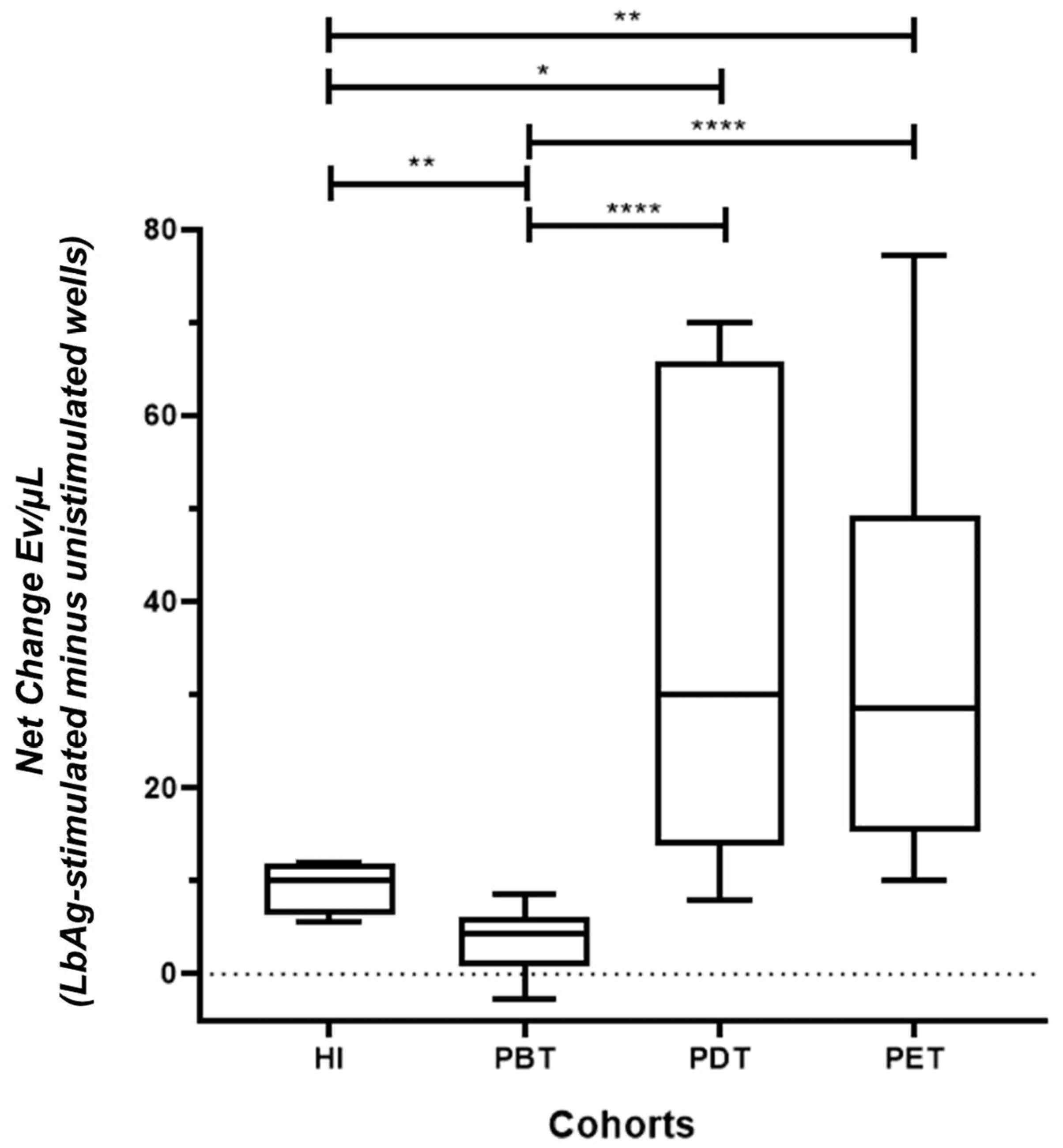
3.3. L. (V.) braziliensis Antigen Stimulation Modulates EV Release in PBMC Cultures
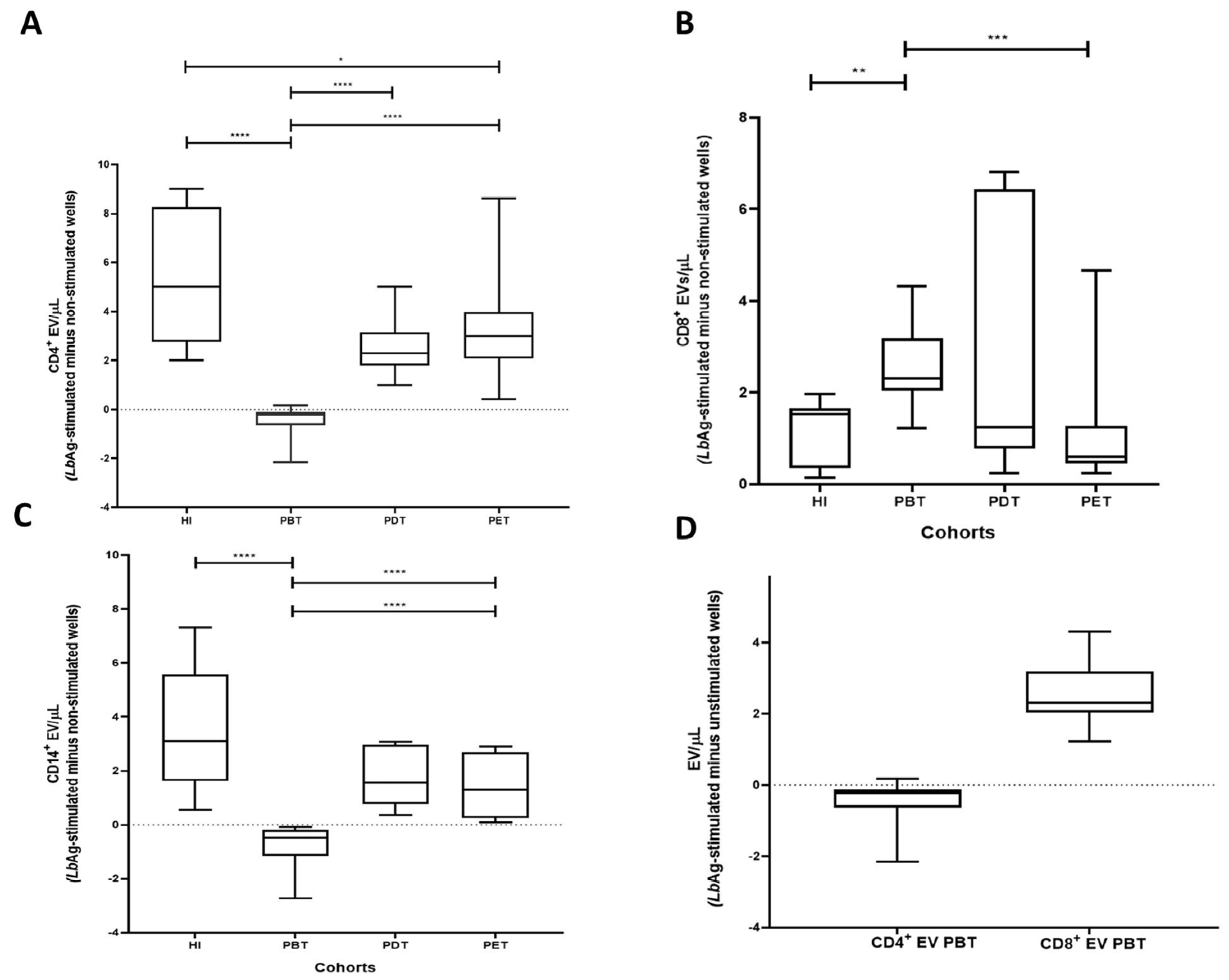
3.4. LbAg-Induced Changes in CD4+, CD8+, and CD14+ EV Subsets
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| nFCM | Nano-flow cytometry |
| LPA | Lymphocyte lymphoproliferative assay |
| EV | Extracellular vesicles |
| PBMCs | Peripheral blood mononuclear cells |
| LbAg | Leishmania (V.) braziliensis antigens |
| Anx-B | Annexin V buffer |
References
- World Health Organization (WHO). Leishmaniasis.Status of Endemicity of Cutaneous Leishmaniasis. Available online: https://apps.who.int/neglected_diseases/ntddata/leishmaniasis/leishmaniasis (accessed on 24 April 2025).
- BRASIL, Ministério da Saúde. Manual de Vigilância da Leishmaniose Tegumentar. 2017. Available online: https://bvsms.saude.gov.br/bvs/publicacoes/manual_vigilancia_leishmaniose_tegumentar.pdf (accessed on 24 April 2025).
- Organización Panamericana de la Salud (OPAS), OMS Américas. Available online: https://www.paho.org/pt/topicos/leishmaniose (accessed on 24 April 2025).
- de Oliveira-Neto, M.P.; Mattos, M.S.; Perez, M.A.; Da-Cruz, A.M.; Fernandes, O.; Moreira, J.; Gonçalves-Costa, S.C.; Brahin, L.R.; Menezes, C.R.; Pirmez, C. American tegumentary leishmaniasis (ATL) in Rio de Janeiro State, Brazil: Main clinical and epidemiologic characteristics. Int. J. Dermatol. 2000, 39, 506–514. [Google Scholar] [CrossRef]
- Schubach, A.O.; Marzochi, K.B.F.; Moreira, J.S.; Schubach, T.M.P.; Araújo, M.L.; Francesconi-do-Vale, A.C.; Passos, S.R.L.; Marzochi, M.C.A. Retrospective study of 151 patients with cutaneous leishmaniasis treated with meglumine antimoniate. Rev. Soc. Bras. Med. Trop. 2005, 38, 213–217. [Google Scholar] [CrossRef]
- Scott, P.; Novais, F.O. Cutaneous leishmaniasis: Immune responses in protection and pathogenesis. Nat. Rev. Immunol. 2016, 16, 581–592. [Google Scholar] [CrossRef]
- Oliveira, W.N.; Ribeiro, L.E.; Schrieffer, A.; Machado, P.; Carvalho, E.M.; Bacellar, O. The role of inflammatory and anti-inflammatory cytokines in the pathogenesis of human tegumentary leishmaniasis. Cytokine 2014, 66, 127–132. [Google Scholar] [CrossRef]
- Alimohmmadian, M.H.; Ajdary, S.; Bahrami, F.A. Historic Review of the Role of CD4+ T-Cell Subsets in Development of the Immune Responses against Cutaneous and Visceral Leishmaniases. Iran. Biomed. J. 2022, 26, 99–109. [Google Scholar] [PubMed]
- Bertho, A.L.; Santiago, M.A.; Da-Cruz, A.M.; Coutinho, S.G. Detection of early apoptosis and cell death in T CD4+ and CD8+ cells from lesions of patients with localized cutaneous leishmaniasis. Braz. J. Med. Biol. Res. 2000, 33, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Da-Cruz, A.M.; Bertho, A.L.; Oliveira-Neto, M.P.; Coutinho, S.G. Flow cytometric analysis of cellular infiltrate from American tegumentary leishmaniasis lesions. Br. J. Dermatol. 2005, 153, 537–543. [Google Scholar] [CrossRef]
- Ferraz, R.; Cunha, C.F.; Gomes-Silva, A.; Schubach, A.O.; Pimentel, M.I.; Lyra, M.R.; Mendonça, S.C.; Valete-Rosalino, C.M.; Da-Cruz, A.M.; Bertho, A.L. Apoptosis and frequency of total and effector CD8+ T lymphocytes from cutaneous leishmaniasis patients during antimonial therapy. BMC Infect. Dis. 2015, 15, 74. [Google Scholar] [CrossRef]
- Cunha, C.F.; Ferraz, R.; Pimentel, M.I.F.; Lyra, M.R.; Schubach, A.O.; Da-Cruz, A.M.; Bertho, A.L. Cytotoxic cell involvement in human cutaneous leishmaniasis: Assessments in active disease, under therapy and after clinical cure. Parasite Immunol. 2016, 38, 244–254. [Google Scholar] [CrossRef]
- Cunha, C.F.; Ferraz-Nogueira, R.; Costa, V.F.A.; Pimentel, M.I.F.; Chometon, T.Q.; Lyra, M.R.; Schubach, A.O.; Da-Cruz, A.M.; Bertho, A.L. Contribution of Leishmania braziliensis antigen specific CD4+ T, CD8+ T, NK and CD3+CD56 +NKT cells in the immunopathogenesis of cutaneous leishmaniasis patients: Cytotoxic, activation and exhaustion profiles. PLoS ONE 2020, 15, e0229400. [Google Scholar] [CrossRef]
- Ferraz, R.; Cunha, C.F.; Pimentel, M.I.F.; Lyra, M.R.; Pereira-Da-Silva, T.; Schubach, A.O.; DaCruz, A.M.; Bertho, A.L. CD3+CD4negCD8neg (double negative) T lymphocytes and NKT cells as the main cytotoxic-related-CD107a+ cells in lesions of cutaneous leishmaniasis caused by Leishmania (Viannia) braziliensis. Parasites Vectors 2017, 10, 219–225. [Google Scholar] [CrossRef]
- Faria, D.R.; Souza, P.E.A.; Durães, F.V.; Carvalho, E.M.; Gollob, K.J.; Machado, P.R.; Dutra, W.O. Recruitment of CD8+ T cells expressing granzyme A is associated with lesion progression in human cutaneous leishmaniasis. Parasite Immunol. 2009, 31, 432–439. [Google Scholar] [CrossRef]
- da Silva Santos, C.; Brodskyn, C.I. The Role of CD4 and CD8 T Cells in Human Cutaneous Leishmaniasis. Front Public Health 2014, 2, 165–180. [Google Scholar] [CrossRef]
- Novais, F.O.; Wong, A.C.; Villareal, D.O.; Beiting, D.P.; Scott, P. CD8+ T cells lack local signals to produce IFNγ in the skin during Leishmania infection. J. Immunol. 2018, 200, 1737–1745. [Google Scholar] [CrossRef] [PubMed]
- Yáñez-Mó, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borras, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Ves. 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Ostrowski, M.; Segura, E. Membrane vesicles as conveyors of immune responses. Nature Rev. Immunol. 2009, 9, 581–593. [Google Scholar] [CrossRef]
- Yuana, Y.; Sturk, A.; Nieuwland, R. Extracellular vesicles in physiological and pathological conditions. Blood Rev. 2013, 27, 31–39. [Google Scholar] [CrossRef]
- Kalluri, R. The biology and function of extracellular vesicles in immune response and immunity. Immunity 2024, 57, 1752–1768. [Google Scholar] [CrossRef]
- van Niel, G.; Carter, D.R.F.; Clayton, A.; Lambert, D.W.; Raposo, G.; Vader, P. Challenges and directions in studying cell-cell communication by extracellular vesicles. Nature Rev. Mol. Cell Biol. 2022, 23, 369–382. [Google Scholar] [CrossRef]
- Menay, F.; Cocozza, F.; Gravisaco, M.J.; Elisei, A.; Re, J.I.; Ferella, A.; Waldner, C.; Mongini, C. Extracellular vesicles derived from antigen-presenting cells pulsed with foot and mouth virus vaccine-antigens act as carriers of viral proteins and stimulate B cell response. Front. Immunol. 2024, 15, 1440667. [Google Scholar] [CrossRef]
- Marcilla, A.; Martin-Jaular, L.; Trelis, M.; Souza-Almeida, W.A.; Bernal, D.; de Menezes-Neto, A.; Osuna, A.; Fernandez-Becerra, C.; Almeida, I.C.; Del Portillo, H.A. Extracellular vesicles in parasitic diseases. J. Extracell. Ves. 2014, 3, 25040. [Google Scholar] [CrossRef] [PubMed]
- Martínez-López, M.; Soto, M.; Iborra, S.; Sancho, D. Leishmania Hijacks Myeloid Cells for Immune Escape. Front. Microbiol. 2018, 9, 883. [Google Scholar] [CrossRef]
- Rossi, I.V.; de Almeida, R.F.; Sabatke, B.; de Godoy, L.M.F.; Ramirez, M.I. Trypanosoma cruzi interaction with host tissues modulates the composition of large extracellular vesicles. Sci. Rep. 2024, 14, 5000. [Google Scholar] [CrossRef]
- Sabatke, B.; Rossi, I.V.; Bonato, L.; Fucio, S.; Cortés, A.; Marcilla, A.; Ramirez, M.I. Host-Pathogen Cellular Communication: The Role of Dynamin, Clathrin, and Macropinocytosis in the Uptake of Giardia-Derived Extracellular Vesicles. ACS Infect. Dis. 2025, 11, 954–962. [Google Scholar] [CrossRef]
- Ferreira, L.C.; Quintella, L.P.; Schubach, A.O.; Miranda, L.F.C.; Madeira, M.F.; Pimentel, M.I.F.; Vasconcellos, É.C.F.E.; Lyra, M.R.; Oliveira, R.V.C.; Menezes, R.C. Comparison between Colorimetric In Situ Hybridization, Histopathology, and Immunohistochemistry for the Diagnosis of New World Cutaneous Leishmaniasis in Human Skin Samples. Trop. Med. Infect. Dis. 2022, 7, 344. [Google Scholar] [CrossRef]
- Filgueira, C.P.B.; Moreira, O.C.; Cantanhêde, L.M.; de Farias, H.M.T.; Porrozzi, R.; Britto, C.; Boité, M.C.; Cupolillo, E. Comparison and clinical validation of qPCR assays targeting Leishmania 18S rDNA and HSP70 genes in patients with American Tegumentary Leishmaniasis. PLoS Neglected Trop. Dis. 2020, 14, e0008750. [Google Scholar] [CrossRef]
- Cupolillo, E.; Grimaldi, G., Jr.; Momen, H. A general classification of New World Leishmania using numerical zymotaxonomy. Am. J. Trop. Med. Hyg. 1994, 50, 296–311. [Google Scholar] [CrossRef]
- Shekari, F.; Alibhai, F.J.; Baharvand, H.; Börger, V.; Bruno, S.; Davies, O.; Giebel, B.; Gimona, M.; Salekdeh, G.H.; Martin-Jaular, L.; et al. Cell culture-derived extracellular vesicles: Considerations for reporting cell culturing parameters. J. Extracell Biol. 2023, 2, e115. [Google Scholar] [CrossRef]
- Costa, V.F.A.; Chometon, T.Q.; de Castro, K.K.G.; Ponte, M.S.G.; Pimentel, M.I.F.; Lyra, M.R.; Bertho, A.L. Platelet-Derived Microvesicles Contribute to the Pathophysiogenesis of Human Cutaneous Leishmaniasis: A Nano-Flow Cytometric Approach in Plasma Samples from Patients before and under Antimonial Treatment. Microorganisms 2024, 12, 526. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr. Protoc. Cell Biol. 2006, 30, 3.22.1–3.22.29. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.A.; Van Der Pol, E.; Arkesteijn, G.J.A.; Bremer, M.; Brisson, A.; Coumans, F.; Dignat-George, F.; Duggan, E.; Ghiran, I.; Giebel, B.; et al. MIFlowCyt-EV: A framework for standardized reporting of extracellular vesicle flow cytometry experiments. J. Extracell. Ves. 2020, 9, 1713526. [Google Scholar] [CrossRef]
- Welsh, J.A.; Goberdhan, D.C.I.; O’Driscoll, L.; Buzas, E.I.; Blenkiron, C.; Bussolati, B.; Cai, H.; Di Vizio, D.; Driedonks, T.A.P.; Erdbrügger, U.; et al. Minimal information for studies of extracellular vesicles (MISEV2023): From basic to advanced approaches. J. Extracell. Ves. 2024, 13, e12404. [Google Scholar] [CrossRef]
- Welsh, J.A.; Arkesteijn, G.J.A.; Bremer, M.; Cimorelli, M.; Dignat-George, F.; Giebel, B.; Görgens, A.; Hendrix, A.; Kuiper, M.; Lacroix, R.; et al. A compendium of single extracellular vesicle flow cytometry. J. Extracell. Ves. 2023, 12, e12299. [Google Scholar] [CrossRef]
- Cruz, C.G.; Sodawalla, H.M.; Mohanakumar, T.; Bansal, S. Extracellular Vesicles as Biomarkers in Infectious Diseases. Biology 2025, 14, 182. [Google Scholar] [CrossRef]
- Abida; Alhuthali, H.M.; Alshehri, J.M.; Alkathiri, A.; Almaghrabi, R.O.M.; Alsaeed, S.S.; Albebi, S.A.H.; Almethn, R.M.; Alfuraydi, B.A.; Alharbi, S.B.; et al. Exosomes in infectious diseases: Insights into leishmaniasis pathogenesis, immune modulation, and therapeutic potential. Naunyn Schmiedebergs Arch. Pharmacol. 2025, 398, 4913–4931. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, A.A.S.; Torrecilhas, A.C.; Souza, B.S.F.; Cruz, F.F.; Guedes, H.L.M.; Ramos, T.D.; Lopes-Pacheco, M.; Caruso-Neves, C.; Rocco, P.R.M. Potential of extracellular vesicles in the pathogenesis, diagnosis and therapy for parasitic diseases. J. Extracell. Ves. 2024, 13, e12496. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.; Weber, J.I.; Durães-Oliveira, J.; Moreno, C.; Ferla, M.; Aires Pereira, M.; Valério-Bolas, A.; Freitas, B.E.; Nunes, T.; Antunes, W.T.; et al. Extracellular Vesicles Derived from Trypanosomatids: The Key to Decoding Host-Parasite Communication. Int. J. Mol. Sci. 2025, 26, 4302. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Andrade, J.M.; Salgado-Bautista, D.; Ramirez-Acosta, K.; Cadena-Nava, R.D.; Riquelme, M. A practical protocol for correlative confocal fluorescence and transmission electron microscopy characterization of extracellular vesicles. Microbiol. Spectr. 2025, 22, e0302624. [Google Scholar] [CrossRef]
- McVey, M.J.; Spring, C.M.; Kuebler, W.M. Improved resolution in extracellular vesicle populations using 405 instead of 488 nm side scatter. J. Extracell. Ves. 2018, 7, 1454776. [Google Scholar] [CrossRef]
- Brittain, G.C.; Chen, Y.Q.; Martinez, E.; Tang, V.A.; Renner, T.M.; Langlois, M.A.; Gulnik, S. A Novel Semiconductor-Based Flow Cytometer with Enhanced Light-Scatter Sensitivity for the Analysis of Biological Nanoparticles. Sci. Rep. 2019, 9, 16039. [Google Scholar] [CrossRef]
- Gray, W.D.; Mitchell, A.J.; Searles, C.D. An accurate, precise method for general labeling of extracellular vesicles. MethodsX 2015, 2, 360–367. [Google Scholar] [CrossRef]
- Austermann, J.; Roth, J.; Barczyk-Kahlert, K. The Good and the Bad: Monocytes and Macrophages Diverse Functions in Inflammation. Cells 2022, 11, 1979. [Google Scholar] [CrossRef]
- Silverman, J.M.; Clos, J.; Horakova, E.; Wang, A.Y.; Wiesgigl, M.; Kelly, I.; Lynn, M.A.; McMaster, W.R.; Foster, L.J.; Levings, M.K.; et al. Leishmania Exosomes Modulate Innate and Adaptive Immune Responses through Effects on Monocytes and Dendritic Cells. J. Immunol. 2010, 185, 5011–5022. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cohorts | ||||
|---|---|---|---|---|
| Variable | PBT | PDT | PET | HI |
| (n) | 17 | 10 | 10 | 6 |
| Sex | ||||
| Female | 7 | 5 | 5 | 3 |
| Male | 10 | 5 | 5 | 3 |
| Age (mean±SD) | 34.06 ± 14.25 | 42.83 ± 15.60 | 31.60 ± 14.40 | 29.6 ± 10.10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, V.F.d.A.; Chometon, T.Q.; Gomes de Castro, K.K.; Ponte, M.S.G.; Pimentel, M.I.F.; Lyra, M.R.; Nieuwland, R.; Bertho, A.L. Extracellular Vesicle Release from Immune Cells in Cutaneous Leishmaniasis: Modulation by Leishmania (V.) braziliensis and Reversal by Antimonial Therapy. Pathogens 2025, 14, 771. https://doi.org/10.3390/pathogens14080771
Costa VFdA, Chometon TQ, Gomes de Castro KK, Ponte MSG, Pimentel MIF, Lyra MR, Nieuwland R, Bertho AL. Extracellular Vesicle Release from Immune Cells in Cutaneous Leishmaniasis: Modulation by Leishmania (V.) braziliensis and Reversal by Antimonial Therapy. Pathogens. 2025; 14(8):771. https://doi.org/10.3390/pathogens14080771
Chicago/Turabian StyleCosta, Vanessa Fernandes de Abreu, Thaize Quiroga Chometon, Katherine Kelda Gomes de Castro, Melissa Silva Gonçalves Ponte, Maria Inês Fernandes Pimentel, Marcelo Rosandiski Lyra, Rienk Nieuwland, and Alvaro Luiz Bertho. 2025. "Extracellular Vesicle Release from Immune Cells in Cutaneous Leishmaniasis: Modulation by Leishmania (V.) braziliensis and Reversal by Antimonial Therapy" Pathogens 14, no. 8: 771. https://doi.org/10.3390/pathogens14080771
APA StyleCosta, V. F. d. A., Chometon, T. Q., Gomes de Castro, K. K., Ponte, M. S. G., Pimentel, M. I. F., Lyra, M. R., Nieuwland, R., & Bertho, A. L. (2025). Extracellular Vesicle Release from Immune Cells in Cutaneous Leishmaniasis: Modulation by Leishmania (V.) braziliensis and Reversal by Antimonial Therapy. Pathogens, 14(8), 771. https://doi.org/10.3390/pathogens14080771