
Assessing Onchocerca volvulus Intensity of Infection and Genetic Diversity Using Mitochondrial Genome Sequencing of Single Microfilariae Obtained before and after Ivermectin Treatment

 , , , ,
, , , ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Origin of Microfilariae
2.2. Origin of Adult Worms and Adult Worm Sequences
2.3. DNA Extraction, Amplification and Sequencing
2.4. Variant Calling
2.5. Data Analysis
3. Results
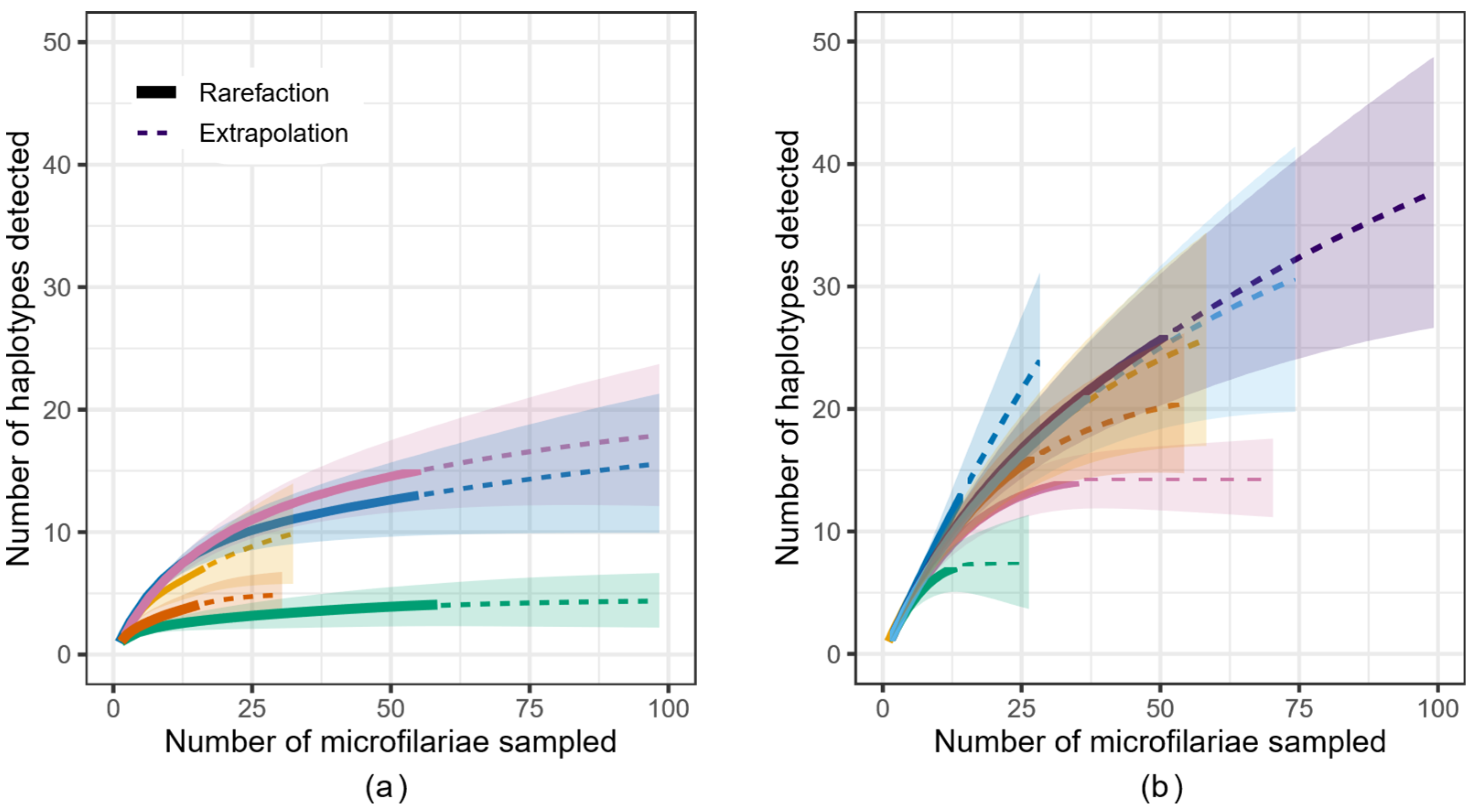
3.1. Sequencing Results
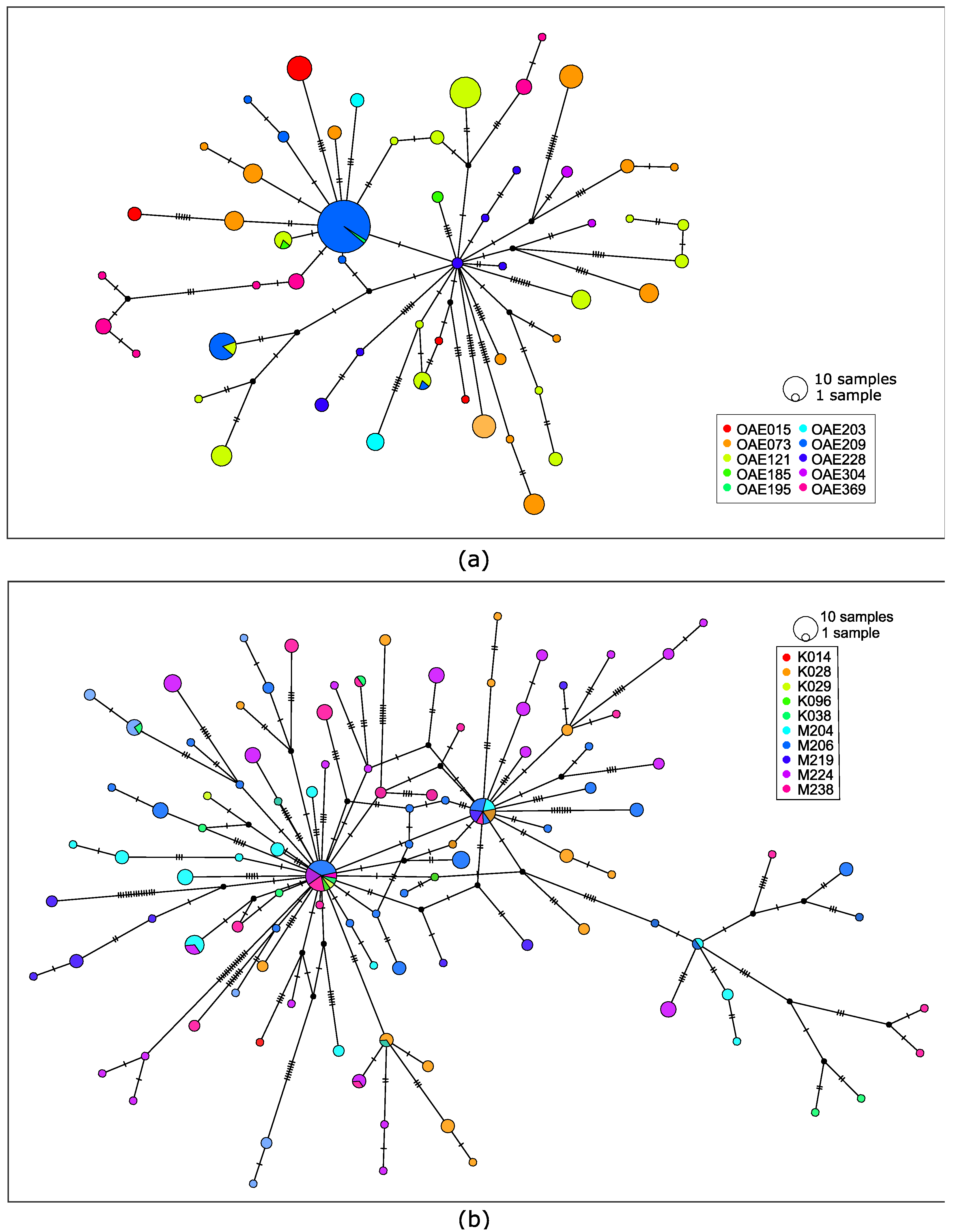
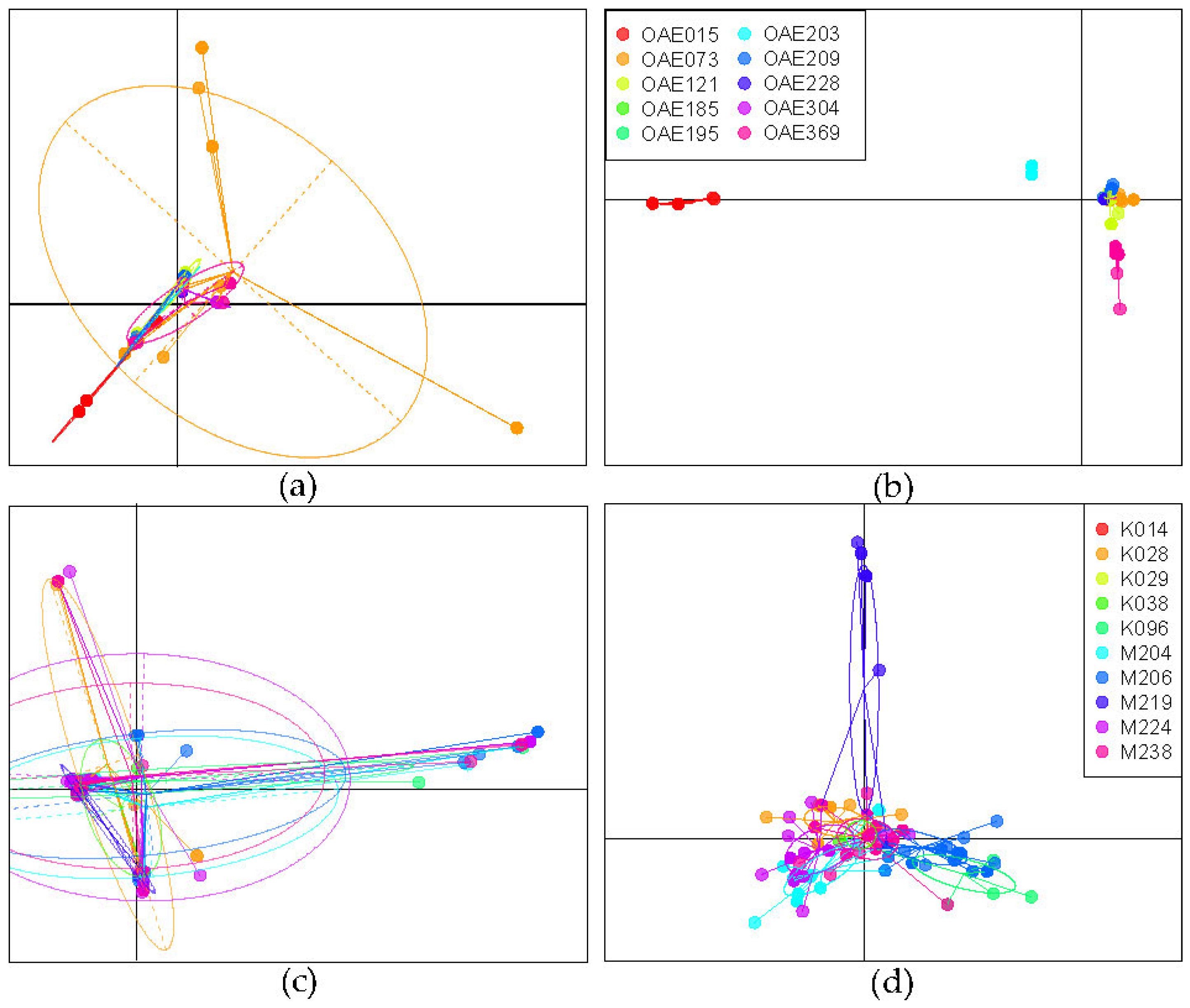
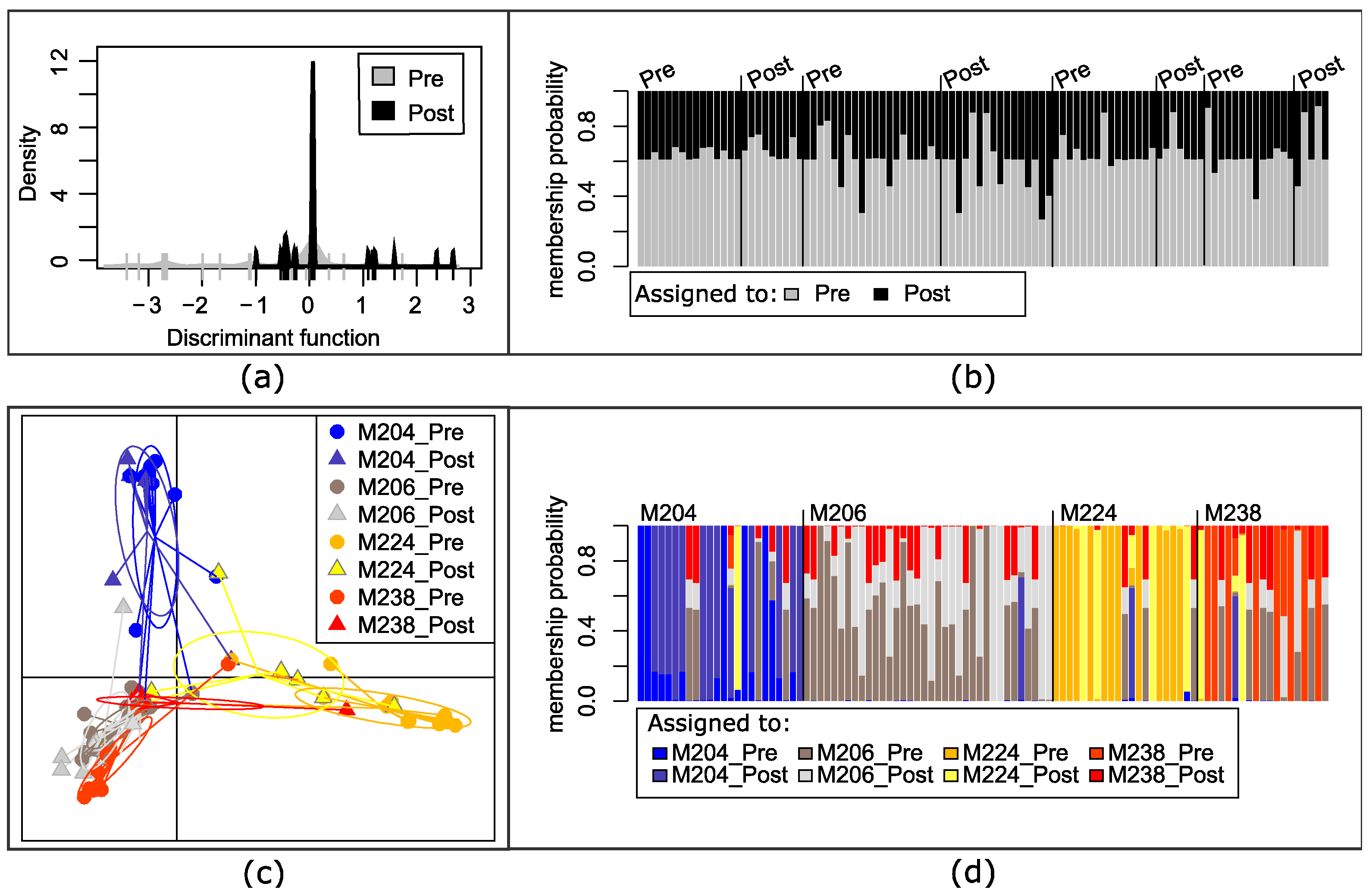
3.2. Genetic Variation of Microfilariae within People

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | Person | Mean Number of Microfilariae Pre/Post [37] | Number of Microfilariae in Dataset Pre/Post | Number of Haplotypes Pre/Post (Total) |
|---|---|---|---|---|
| DRC | OAE015 | 48.5/3 | 0/15 | 0/4 (4) |
| OAE073 | 191/0 | 55/0 | 13/0 (13) | |
| OAE121 | 295.5/0 | 55/0 | 13/0 (13) | |
| OAE185 | 18.5/29 | 2/0 | 1/0 (1) | |
| OAE195 | 12.5/0 | 2/0 | 2/0 (2) | |
| OAE203 | 28/0 | 8/0 | 2/0 (2) | |
| OAE209 | 370.5/0 | 60/0 | 6/0 (6) | |
| OAE228 | 40/0 | 9/0 | 6/0 (6) | |
| OAE304 | 9/4 | 3/0 | 2/0 (2) | |
| OAE369 | 52.5/0 | 16/0 | 7/0 (7) | |
| All DRC | 210/15 | 52/4 (52) | ||
| South Sudan | K014 | 16.5/20.5 | 0/1 | 0/1 (1) |
| K028 | 78.5/20 | 27/0 | 16/0 (16) | |
| K029 | 57/143 | 0/1 | 0/1 (1) | |
| K038 | 23.5/24 | 3/0 | 3/0 (3) | |
| K096 | 108.5/1.5 | 9/0 | 8/0 (8) | |
| M204 | 59.5/46.5 | 27/10 | 13/8 (17) | |
| M206 | 105.5/62 | 33/21 | 19/ 16 (28) | |
| M219 | 45.5/64 | 13/0 | 8/0 (8) | |
| M224 | 23.5/51 | 28/9 | 15/7 (20) | |
| M238 | 23/55 | 23/6 | 14/5 (18) | |
| All South Sudan | 163/48 | 96/38 (103) | ||
| Totals | 373/63 | 148/42 (155) |
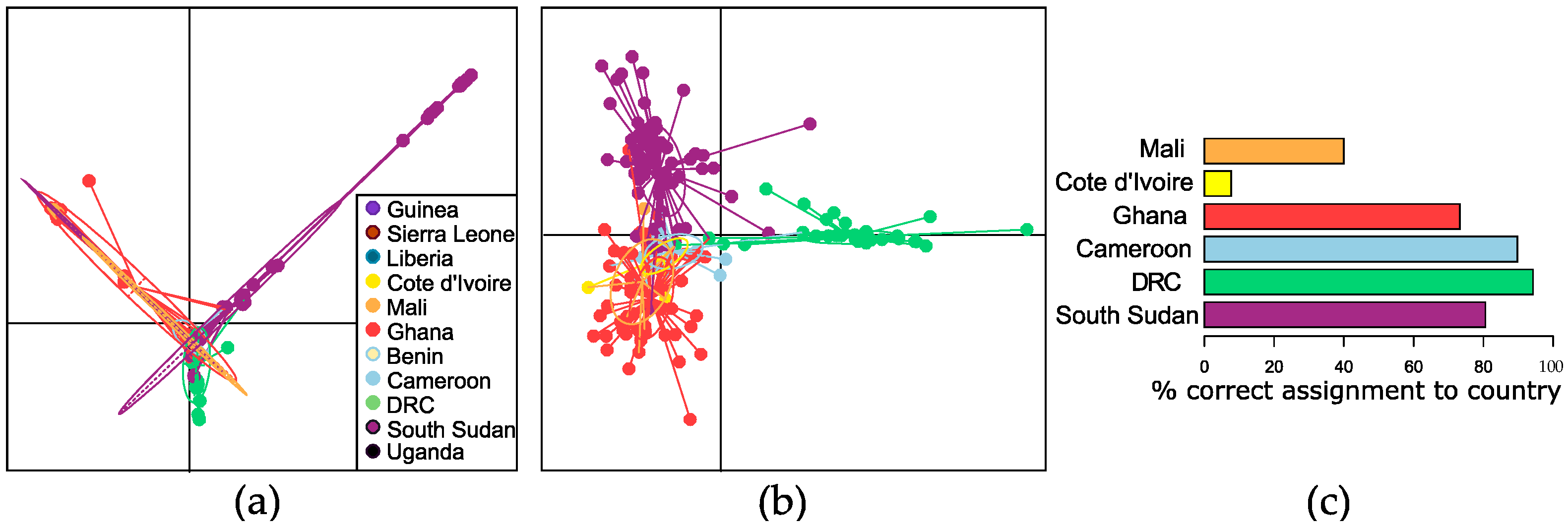
3.3. Genetic Variation in the African Context
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethics Committee Approval
References
- World Health Organization. Elimination of human onchocerciasis: Progress report, 2020. Wkly. Epidemiol. Rec. 2021, 46, 557–567. [Google Scholar]
- Chesnais, C.B.; Nana-Djeunga, H.C.; Njamnshi, A.K.; Lenou-Nanga, C.G.; Boulle, C.; Bissek, A.Z.; Kamgno, J.; Colebunders, R.; Boussinesq, M. The temporal relationship between onchocerciasis and epilepsy: A population-based cohort study. Lancet Infect. Dis. 2018, 18, 1278–1286. [Google Scholar] [CrossRef] [PubMed]
- Colebunders, R.; Njamnshi, A.K.; van Oijen, M.; Mukendi, D.; Kashama, J.M.; Mandro, M.; Gumisiriza, N.; Preux, P.M.; Suykerbuyk, P.; Idro, R. Onchocerciasis-associated epilepsy: From recent epidemiological and clinical findings to policy implications. Epilepsia Open 2017, 2, 145–152. [Google Scholar] [CrossRef]
- Colebunders, R.; Njamnshi, A.K.; Menon, S.; Newton, C.R.; Hotterbeekx, A.; Preux, P.M.; Hopkins, A.; Vaillant, M.; Siewe Fodjo, J.N. Onchocerca volvulus and epilepsy: A comprehensive review using the Bradford Hill criteria for causation. PLoS Negl. Trop. Dis. 2021, 15, e0008965. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Ending the Neglect to Attain the Sustainable Development Goals: A Road Map for Neglected Tropical Diseases 2021–2030; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Plaisier, A.P.; Alley, E.S.; Boatin, B.A.; Van Oortmarssen, G.J.; Remme, H.; De Vlas, S.J.; Bonneux, L.; Habbema, J.D. Irreversible effects of ivermectin on adult parasites in onchocerciasis patients in the Onchocerciasis Control Programme in West Africa. J. Infect. Dis. 1995, 172, 204–210. [Google Scholar] [CrossRef]
- Basáñez, M.-G.; Pion, S.D.; Boakes, E.; Filipe, J.A.; Churcher, T.S.; Boussinesq, M. Effect of single-dose ivermectin on Onchocerca volvulus: A systematic review and meta-analysis. Lancet Infect. Dis. 2008, 8, 310–322. [Google Scholar] [CrossRef]
- Bottomley, C.; Isham, V.; Collins, R.C.; Basáñez, M.G. Rates of microfilarial production by Onchocerca volvulus are not cumulatively reduced by multiple ivermectin treatments. Parasitology 2008, 135, 1571–1581. [Google Scholar] [CrossRef]
- Turner, H.C.; Walker, M.; Churcher, T.S.; Basáñez, M.G. Modelling the impact of ivermectin on River blindness and its burden of morbidity and mortality in African savannah: EpiOncho projections. Parasit. Vectors 2014, 7, 241. [Google Scholar] [CrossRef]
- Sauerbrey, M.; Rakers, L.J.; Richards, F.O. Progress toward elimination of onchocerciasis in the Americas. Int. Health 2018, 10, i71–i78. [Google Scholar] [CrossRef]
- World Health Organization. Elimination of human onchocerciasis: Progress report, 2021—Élimination de l’onchocercose humaine: Rapport de situation, 2021. Wkly. Epidemiol. Rec. 2022, 96, 591–598. [Google Scholar]
- Tekle, A.H.; Zouré, H.G.; Noma, M.; Boussinesq, M.; Coffeng, L.E.; Stolk, W.A.; Remme, J.H. Progress towards onchocerciasis elimination in the participating countries of the African Programme for Onchocerciasis Control: Epidemiological evaluation results. Infect. Dis. Poverty 2016, 5, 66. [Google Scholar] [CrossRef]
- Diawara, L.; Traore, M.O.; Badji, A.; Bissan, Y.; Doumbia, K.; Goita, S.F.; Konate, L.; Mounkoro, K.; Sarr, M.D.; Seck, A.F.; et al. Feasibility of onchocerciasis elimination with ivermectin treatment in endemic foci in Africa: First evidence from studies in Mali and Senegal. PLoS Negl. Trop. Dis. 2009, 3, e497. [Google Scholar] [CrossRef] [PubMed]
- Traore, M.O.; Sarr, M.D.; Badji, A.; Bissan, Y.; Diawara, L.; Doumbia, K.; Goita, S.F.; Konate, L.; Mounkoro, K.; Seck, A.F.; et al. Proof-of-principle of onchocerciasis elimination with ivermectin treatment in endemic foci in Africa: Final results of a study in Mali and Senegal. PLoS Negl. Trop. Dis. 2012, 6, e1825. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization; African Programme for Onchocerciasis Control. Report of the Consultative Meetings on Strategic Options and Alternative Treatment Strategies for Accelerating Onchocerciasis Elimination in Africa. World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Awadzi, K.; Attah, S.K.; Addy, E.T.; Opoku, N.O.; Quartey, B.T.; Lazdins-Helds, J.K.; Ahmed, K.; Boatin, B.A.; Boakye, D.A.; Edwards, G. Thirty-month follow-up of sub-optimal responders to multiple treatments with ivermectin, in two onchocerciasis-endemic foci in Ghana. Ann. Trop. Med. Parasitol. 2004, 98, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Awadzi, K.; Boakye, D.A.; Edwards, G.; Opoku, N.O.; Attah, S.K.; Osei-Atweneboana, M.Y.; Lazdins-Helds, J.K.; Ardrey, A.E.; Addy, E.T.; Quartey, B.T.; et al. An investigation of persistent microfilaridermias despite multiple treatments with ivermectin, in two onchocerciasis-endemic foci in Ghana. Ann. Trop. Med. Parasitol. 2004, 98, 231–249. [Google Scholar] [CrossRef] [PubMed]
- Churcher, T.S.; Pion, S.D.; Osei-Atweneboana, M.Y.; Prichard, R.K.; Awadzi, K.; Boussinesq, M.; Collins, R.C.; Whitworth, J.A.; Basáñez, M.G. Identifying sub-optimal responses to ivermectin in the treatment of River Blindness. Proc. Natl. Acad. Sci. USA 2009, 106, 16716–16721. [Google Scholar] [CrossRef]
- Osei-Atweneboana, M.Y.; Eng, J.K.; Boakye, D.A.; Gyapong, J.O.; Prichard, R.K. Prevalence and intensity of Onchocerca volvulus infection and efficacy of ivermectin in endemic communities in Ghana: A two-phase epidemiological study. Lancet 2007, 369, 2021–2029. [Google Scholar] [CrossRef] [PubMed]
- Doyle, S.R.; Bourguinat, C.; Nana-Djeunga, H.C.; Kengne-Ouafo, J.A.; Pion, S.D.S.; Bopda, J.; Kamgno, J.; Wanji, S.; Che, H.; Kuesel, A.C.; et al. Genome-wide analysis of ivermectin response by Onchocerca volvulus reveals that genetic drift and soft selective sweeps contribute to loss of drug sensitivity. PLoS Negl. Trop. Dis. 2017, 11, e0005816. [Google Scholar] [CrossRef]
- Osei-Atweneboana, M.Y.; Awadzi, K.; Attah, S.K.; Boakye, D.A.; Gyapong, J.O.; Prichard, R.K. Phenotypic evidence of emerging ivermectin resistance in Onchocerca volvulus. PLoS Negl. Trop. Dis. 2011, 5, e998. [Google Scholar] [CrossRef]
- Nana-Djeunga, H.C.; Bourguinat, C.; Pion, S.D.; Bopda, J.; Kengne-Ouafo, J.A.; Njiokou, F.; Prichard, R.K.; Wanji, S.; Kamgno, J.; Boussinesq, M. Reproductive status of Onchocerca volvulus after ivermectin treatment in an ivermectin-naive and a frequently treated population from Cameroon. PLoS Negl. Trop. Dis. 2014, 8, e2824. [Google Scholar] [CrossRef]
- Opoku, N.O.; Bakajika, D.K.; Kanza, E.M.; Howard, H.; Mambandu, G.L.; Nyathirombo, A.; Nigo, M.M.; Kasonia, K.; Masembe, S.L.; Mumbere, M.; et al. Single dose moxidectin versus ivermectin for Onchocerca volvulus infection in Ghana, Liberia, and the Democratic Republic of the Congo: A randomised, controlled, double-blind phase 3 trial. Lancet 2018, 392, 1207–1216. [Google Scholar] [CrossRef] [PubMed]
- Bakajika, D.; Kanza, E.M.; Opoku, N.O.; Howard, H.M.; Mambandu, G.L.; Nyathirombo, A.; Nigo, M.M.; Kennedy, K.K.; Masembe, S.L.; Mumbere, M.; et al. Effect of a single dose of 8 mg moxidectin or 150 μg/kg ivermectin on O. volvulus skin microfilariae in a randomized trial: Differences between areas in the Democratic Republic of the Congo, Liberia and Ghana and impact of intensity of infection. PLoS Negl. Trop. Dis. 2022, 16, e0010079. [Google Scholar] [CrossRef]
- Awadzi, K.; Opoku, N.O.; Attah, S.K.; Lazdins-Helds, J.; Kuesel, A.C. A randomized, single-ascending-dose, ivermectin-controlled, double-blind study of moxidectin in Onchocerca volvulus infection. PLoS Negl. Trop. Dis. 2014, 8, e2953. [Google Scholar] [CrossRef]
- Frempong, K.K.; Walker, M.; Cheke, R.A.; Tetevi, E.J.; Gyan, E.T.; Owusu, E.O.; Wilson, M.D.; Boakye, D.A.; Taylor, M.J.; Biritwum, N.K.; et al. Does increasing treatment frequency address suboptimal responses to ivermectin for the control and elimination of River blindness? Clin. Infect. Dis. 2016, 62, 1338–1347. [Google Scholar] [CrossRef]
- Abong, R.A.; Amambo, G.N.; Chounna Ndongmo, P.W.; Njouendou, A.J.; Ritter, M.; Beng, A.A.; Esum, M.E.; Deribe, K.; Fru-Cho, J.; Fombad, F.F.; et al. Differential susceptibility of Onchocerca volvulus microfilaria to ivermectin in two areas of contrasting history of mass drug administration in Cameroon: Relevance of microscopy and molecular techniques for the monitoring of skin microfilarial repopulation within six months of direct observed treatment. BMC Infect. Dis. 2020, 20, 726. [Google Scholar] [CrossRef]
- Stolk, W.A.; Walker, M.; Coffeng, L.E.; Basáñez, M.G.; de Vlas, S.J. Required duration of mass ivermectin treatment for onchocerciasis elimination in Africa: A comparative modelling analysis. Parasit. Vectors 2015, 8, 552. [Google Scholar] [CrossRef] [PubMed]
- Basáñez, M.G.; Walker, M.; Turner, H.C.; Coffeng, L.E.; de Vlas, S.J.; Stolk, W.A. River blindness: Mathematical models for control and elimination. Adv. Parasitol. 2016, 94, 247–341. [Google Scholar] [CrossRef]
- Duke, B.O. Onchocerciasis; deep worm bundles close to hip joints. Trans. R. Soc. Trop. Med. Hyg. 1970, 64, 791–792. [Google Scholar] [CrossRef] [PubMed]
- Meyers, W.M.; Neafie, R.C.; Connor, D.H. Onchocerciasis: Invasion of deep organs by Onchocerca volvulus. Am. J. Trop. Med. Hyg. 1977, 26, 650–657. [Google Scholar] [CrossRef]
- Duke, B.O. The population dynamics of Onchocerca volvulus in the human host. Trop. Med. Parasitol. 1993, 44, 61–68. [Google Scholar] [PubMed]
- Duerr, H.P.; Dietz, K.; Schulz-Key, H.; Buttner, D.W.; Eichner, M. The relationships between the burden of adult parasites, host age and the microfilarial density in human onchocerciasis. Int. J. Parasitol. 2004, 34, 463–473. [Google Scholar] [CrossRef]
- Albiez, E.J. Studies on nodules and adult Onchocerca volvulus during a nodulectomy trial in hyperendemic villages in Liberia and Upper Volta. I. Palpable and impalpable onchocercomata. Tropenmed. Parasitol. 1983, 34, 54–60. [Google Scholar]
- Basáñez, M.G.; Boussinesq, M. Population biology of human onchocerciasis. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1999, 354, 809–826. [Google Scholar] [CrossRef]
- Schulz-Key, H.; Albiez, E.J. Worm burden of Onchocerca volvulus in a hyperendemic village of the rain-forest in West Africa. Tropenmedizin Parasitol. 1977, 28, 431–438. [Google Scholar]
- Dusabimana, A.; Bhwana, D.; Raimon, S.; Mmbando, B.P.; Hotterbeekx, A.; Tepage, F.; Mandro, M.; Siewe Fodjo, J.N.; Abrams, S.; Colebunders, R. Ivermectin treatment response in Onchocerca volvulus infected persons with epilepsy: A three-country short cohort study. Pathogens 2020, 9, 617. [Google Scholar] [CrossRef]
- Fodjo, J.N.S.; Mandro, M.; Mukendi, D.; Tepage, F.; Menon, S.; Nakato, S.; Nyisi, F.; Abhafule, G.; Wonya’rossi, D.; Anyolito, A.; et al. Onchocerciasis-associated epilepsy in the Democratic Republic of Congo: Clinical description and relationship with microfilarial density. PLoS Negl. Trop. Dis. 2019, 13, e0007300. [Google Scholar] [CrossRef]
- Mandro, M.; Siewe Fodjo, J.N.; Dusabimana, A.; Mukendi, D.; Haesendonckx, S.; Lokonda, R.; Nakato, S.; Nyisi, F.; Abhafule, G.; Wonya’rossi, D.; et al. Single versus multiple dose ivermectin regimen in onchocerciasis-infected persons with epilepsy treated with phenobarbital: A randomized clinical trial in the Democratic Republic of Congo. Pathogens 2020, 9, 205. [Google Scholar] [CrossRef] [PubMed]
- Abd-Elfarag, G.; Carter, J.Y.; Raimon, S.; Sebit, W.; Suliman, A.; Fodjo, J.N.S.; Olore, P.C.; Biel, K.P.; Ojok, M.; Logora, M.Y.; et al. Persons with onchocerciasis-associated epilepsy and nodding seizures have a more severe form of epilepsy with more cognitive impairment and higher levels of Onchocerca volvulus infection. Epileptic Disord. 2020, 22, 301–308. [Google Scholar] [CrossRef]
- Colebunders, R.; Carter, J.Y.; Olore, P.C.; Puok, K.; Bhattacharyya, S.; Menon, S.; Abd-Elfarag, G.; Ojok, M.; Ensoy-Musoro, C.; Lako, R.; et al. High prevalence of onchocerciasis-associated epilepsy in villages in Maridi County, Republic of South Sudan: A community-based survey. Seizure 2018, 63, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Post, R.J.; Laudisoit, A.; Mandro, M.; Lakwo, T.; Laemmer, C.; Pfarr, K.; Hoerauf, A.; Tortosa, P.; Gomard, Y.; Ukety, T.; et al. Identification of the onchocerciasis vector in the Kakoi-Koda focus of the Democratic Republic of Congo. PLoS Negl. Trop. Dis. 2022, 16, e0010684. [Google Scholar] [CrossRef] [PubMed]
- Nana-Djeunga, H.C.; Sicard, C.M.; Mogoung-Wafo, A.E.; Chesnais, C.B.; Deleglise, H.; Touka-Nounkeu, R.; Domche, A.; Golden, A.; Klion, A.D.; Nutman, T.B.; et al. Changes in onchocerciasis Ov16 IgG4 rapid diagnostic test results over one-month follow-up: Lessons for reading timeframe and decision-making. Am. J. Trop. Med. Hyg. 2022, 107, 658–661. [Google Scholar] [CrossRef]
- Hotterbeekx, A.; Perneel, J.; Mandro, M.; Abhafule, G.; Siewe Fodjo, J.N.; Dusabimana, A.; Abrams, S.; Kumar-Singh, S.; Colebunders, R. Comparison of diagnostic tests for Onchocerca volvulus in the Democratic Republic of Congo. Pathogens 2020, 9, 435. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Report of the Proceedings of the Fifth Meeting of the Onchocerciasis Technical Advisory Subgroup, Virtual Meeting, 9–10 December 2021; World Health Organization: Geneva, Switzerland, 2023. [Google Scholar]
- World Health Organization. Report on the Fourth Meeting of the WHO Onchocerciasis Technical Advisory Subgroup: Virtual Meeting, 28–29 October 2020; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Dusabimana, A.; Siewe Fodjo, J.N.; Ndahura, M.M.; Mmbando, B.P.; Jada, S.R.; Boven, A.; De Smet, E.; Ukety, T.; Njamnshi, A.K.; Laudisoit, A.; et al. Surveillance for onchocerciasis-associated epilepsy and OV16 IgG4 testing of children 6–10 years old should be used to identify areas where onchocerciasis elimination programs need strengthening. Pathogens 2022, 11, 281. [Google Scholar] [CrossRef]
- Lakwo, T.L.; Raimon, S.; Tionga, M.; Siewe Fodjo, J.N.; Alinda, P.; Sebit, W.J.; Carter, J.Y.; Colebunders, R. The role of the Maridi Dam in causing an onchocerciasis-associated epilepsy epidemic in Maridi, South Sudan: An epidemiological, sociological, and entomological study. Pathogens 2020, 9, 315. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Tyagi, R.; McNulty, S.N.; Rosa, B.A.; Ozersky, P.; Martin, J.; Hallsworth-Pepin, K.; Unnasch, T.R.; Norice, C.T.; Nutman, T.B.; et al. Genomic diversity in Onchocerca volvulus and its Wolbachia endosymbiont. Nat. Microbiol. 2016, 2, 16207. [Google Scholar] [CrossRef]
- Crawford, K.E.; Hedtke, S.M.; Kuesel, A.C.; Doyle, S.D.; Armoo, S.; Osei-Atweneboana, M.; Grant, W.N. Genome-based tools for onchocerciasis elimination: Utility of the mitochondrial genome for delineating Onchocerca volvulus transmission zones. bioRxiv 2019, 732446. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Burns, M.; Valdivia, H. Modelling the limit of detection in real-time quantitative PCR. Eur. Food Res. Technol. 2008, 226, 1513–1524. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Cotton, J.A.; Bennuru, S.; Grote, A.; Harsha, B.; Tracey, A.; Beech, R.; Doyle, S.R.; Dunn, M.; Hotopp, J.C.; Holroyd, N.; et al. The genome of Onchocerca volvulus, agent of river blindness. Nat. Microbiol. 2016, 2, 16216. [Google Scholar] [CrossRef]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Quinlan, A.R. BEDTools: The Swiss-Army Tool for Genome Feature Analysis. Curr. Protoc. Bioinforma. 2014, 47, 11–12. [Google Scholar] [CrossRef]
- Price, A.L.; Jones, N.C.; Pevzner, P.A. De novo identification of repeat families in large genomes. Bioinformatics 2005, 21 (Suppl. 1), i351–i358. [Google Scholar] [CrossRef]
- Garrison, E.; Marth, G. Haplotype-based variant detection from short-read sequencing. arXiv 2012, arXiv:1207.3907. [Google Scholar] [CrossRef]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef] [PubMed]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve years of SAMtools and BCFtools. Gigascience 2021, 10, giab008. [Google Scholar] [CrossRef] [PubMed]
- Garrison, E.; Kronenberg, Z.N.; Dawson, E.T.; Pedersen, B.S.; Prins, P. Vcflib and tools for processing the VCF variant call format. bioRxiv 2021. [Google Scholar] [CrossRef]
- Jombart, T.; Ahmed, I. adegenet 1.3-1: New tools for the analysis of genome-wide SNP data. Bioinformatics 2011, 27, 3070–3071. [Google Scholar] [CrossRef]
- R Development Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2008.
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 2010, 11, 94. [Google Scholar] [CrossRef]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Chao, A. Estimating the population size for capture-recapture data with unequal catchability. Biometrics 1987, 43, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.H.; Wang, Y.T.; Walther, B.A.; Chao, A. An improved nonparametric lower bound of species richness via a modified good-turing frequency formula. Biometrics 2014, 70, 671–682. [Google Scholar] [CrossRef]
- O’Hara, R.B. Species richness estimators: How many species can dance on the head of a pin? J. Anim. Ecol. 2005, 74, 375–386. [Google Scholar] [CrossRef]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef]
- Lischer, H.E.; Excoffier, L. PGDSpider: An automated data conversion tool for connecting population genetics and genomics programs. Bioinformatics 2012, 28, 298–299. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar] [CrossRef]
- Churcher, T.S.; Schwab, A.E.; Prichard, R.K.; Basanez, M.G. An analysis of genetic diversity and inbreeding in Wuchereria bancrofti: Implications for the spread and detection of drug resistance. PLoS Negl. Trop. Dis. 2008, 2, e211. [Google Scholar] [CrossRef]
- Hedtke, S.M.; Zendejas-Heredia, P.A.; Graves, P.M.; Sheridan, S.; Sheel, M.; Fuimaono, S.; Lau, C.L.; Grant, W.N. Genetic epidemiology of lymphatic filariasis in American Samoa after mass drug administration. Int. J. Parasitol. 2021, 15, 137–147. [Google Scholar] [CrossRef]
- World Health Organization; Onchocerciasis Control Programme in the Volta River Basin Area; Rolland, A.; Balay, G. Onchocerciasis Focus in the Bissa Country; Onchocercisis Control Programme in the Volta River Basin Area: Ouagadougou, Burkino Faso, 1985. [Google Scholar]
- Ngoumou, P.; Walsh, J.F. ; WHO Programme for the Prevention of Blindness. UNDP/World Bank/WHO Special Programme for Research and Training in Tropical Diseases. A Manual for Rapid Epidemiological Mapping of Onchocerciasis; Ngoumou, P., Walsh, J.F., Eds.; World Health Organization: Geneva, Switzerland, 1993. [Google Scholar]
- Remme, J.H.F.; Boatin, B.; Boussinesq, M. Helminthic diseases: Onchocerciasis and loiasis. In The International Encyclopedia of Public Health, 2nd ed.; Quah, S.R., Cockerham, W.C., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 3, pp. 576–587. [Google Scholar]
- Duke, B.O. Observations and reflections on the immature stages of Onchocerca volvulus in the human host. Ann. Trop. Med. Parasitol. 1991, 85, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Nelson, G.S. Human onchocerciasis: Notes on the history, the parasite and the life cycle. Ann. Trop. Med. Parasitol. 1991, 85, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Prost, A. Latence parasitaire dans l’onchocercose. Bull. World Health Organ. 1980, 58, 923–925. [Google Scholar]
- Hedtke, S.M.; Kuesel, A.C.; Crawford, K.E.; Graves, P.M.; Boussinesq, M.; Lau, C.L.; Boakye, D.A.; Grant, W.N. Genomic epidemiology in filarial nematodes: Transforming the basis for elimination program decisions. Front. Genet. 2020, 10, 1282. [Google Scholar] [CrossRef]
- Koala, L.; Nikiema, A.; Post, R.J.; Paré, A.B.; Kafando, C.M.; Drabo, F.; Traoré, S. Recrudescence of onchocerciasis in the Comoé valley in Southwest Burkina Faso. Acta Trop. 2017, 166, 96–105. [Google Scholar] [CrossRef]
- Koala, L.; Nikiema, A.S.; Pare, A.B.; Drabo, F.; Toe, L.D.; Belem, A.M.G.; Boakye, D.A.; Traore, S.; Dabire, R.K. Entomological assessment of the transmission following recrudescence of onchocerciasis in the Comoé Valley, Burkina Faso. Parasit. Vectors 2019, 12, 34. [Google Scholar] [CrossRef] [PubMed]
- Garms, R. The reinvasion of the onchocerciasis control programme area in the Volta River Basin by Simulium damnosum s.l., the involvement of the different cytospecies and epidemiological implications. Ann. Soc. Belg. Med. Trop. 1981, 61, 193–198. [Google Scholar]
- Garms, R.; Walsh, J.F.; Davies, J.B. Studies on the reinvasion of the Onchocerciasis Control Programme in the Volta River Basin by Simulium damnosum s.I. with emphasis on the south-western areas. Tropenmed Parasitol. 1979, 30, 345–362. [Google Scholar]
- Magor, J.I.; Rosenberg, L.J. Studies of winds and weather during migrations of Simulium damnosum Theobald (Diptera: Simuliidae), the vector of onchocerciasis in West Africa. Bull. Entomol. Res. 1980, 70, 693–716. [Google Scholar] [CrossRef]
- Baker, R.H.; Guillet, P.; Seketeli, A.; Poudiougo, P.; Boakye, D.; Wilson, M.D.; Bissan, Y.; Garms, R.; Cheke, R.A.; Sachs, R.; et al. Progress in controlling the reinvasion of windborne vectors into the western area of the Onchocerciasis Control Programme in West Africa [and discussion]. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1990, 328, 731–747. [Google Scholar] [CrossRef]
- McCulloch, K.; Hedtke, S.M.; McCaw, J.; McVernon, J.; Basáñez, M.-G.; Walker, M.; Kuesel, A.C.; Grant, W.N. Impact of human movement between hypo- and hyperendemic areas on sustainability of elimination of Onchocerca volvulus transmission. medRxiv 2023. [Google Scholar] [CrossRef]
- de Vos, A.S.; Stolk, W.A.; de Vlas, S.J.; Coffeng, L.E. The effect of assortative mixing on stability of low helminth transmission levels and on the impact of mass drug administration: Model explorations for onchocerciasis. PLoS Negl. Trop. Dis. 2018, 12, e0006624. [Google Scholar] [CrossRef] [PubMed]
- Specht, S.; Hoerauf, A.; Adjei, O.; Debrah, A.; Buttner, D.W. Newly acquired Onchocerca volvulus filariae after doxycycline treatment. Parasitol. Res. 2009, 106, 23–31. [Google Scholar] [CrossRef] [PubMed]





| Person | Sample Size | Observed No. Haplotypes | Chao 1 (SE) | ACE (SE) |
|---|---|---|---|---|
| DRC: OAE015 | 15 | 4 | 5.0 (2.17) | 16.0 (4.14) |
| DRC: OAE073 | 55 | 13 | 16.0 (4.14) | 15.61 (1.47) |
| DRC: OAE121 | 55 | 15 | 18.3 (4.10) | 18.66 (1.89) |
| DRC: OAE209 | 58 | 4 | 4.0 (0.22) | 5.24 (1.31) |
| DRC: OAE369 | 16 | 7 | 13.0 (7.08) | 11.47 (1.49) |
| South Sudan: K028 | 27 | 16 | 20.7 (4.48) | 22.74 (2.33) |
| South Sudan: K096 | 14 | 13 | 46.0 (26.3) | 91.0 (0.98) |
| South Sudan: M204 | 35 | 14 | 14.1 (0.49) | 14.85 (1.19) |
| South Sudan: M206 | 51 | 26 | 52.3 (18.74) | 45.66 (3.50) |
| South Sudan: M219 | 13 | 7 | 7.2 (0.62) | 8.27 (1.36) |
| South Sudan: M224 | 37 | 21 | 36.6 (11.63) | 40.59 (3.18) |
| South Sudan: M238 | 29 | 18 | 29.0 (8.86) | 31.53 (2.69) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hedtke, S.M.; Choi, Y.-J.; Kode, A.; Chalasani, G.C.; Sirwani, N.; Jada, S.R.; Hotterbeekx, A.; Mandro, M.; Siewe Fodjo, J.N.; Amambo, G.N.; et al. Assessing Onchocerca volvulus Intensity of Infection and Genetic Diversity Using Mitochondrial Genome Sequencing of Single Microfilariae Obtained before and after Ivermectin Treatment. Pathogens 2023, 12, 971. https://doi.org/10.3390/pathogens12070971
Hedtke SM, Choi Y-J, Kode A, Chalasani GC, Sirwani N, Jada SR, Hotterbeekx A, Mandro M, Siewe Fodjo JN, Amambo GN, et al. Assessing Onchocerca volvulus Intensity of Infection and Genetic Diversity Using Mitochondrial Genome Sequencing of Single Microfilariae Obtained before and after Ivermectin Treatment. Pathogens. 2023; 12(7):971. https://doi.org/10.3390/pathogens12070971
Chicago/Turabian StyleHedtke, Shannon M., Young-Jun Choi, Anusha Kode, Gowtam C. Chalasani, Neha Sirwani, Stephen R. Jada, An Hotterbeekx, Michel Mandro, Joseph N. Siewe Fodjo, Glory Ngongeh Amambo, and et al. 2023. "Assessing Onchocerca volvulus Intensity of Infection and Genetic Diversity Using Mitochondrial Genome Sequencing of Single Microfilariae Obtained before and after Ivermectin Treatment" Pathogens 12, no. 7: 971. https://doi.org/10.3390/pathogens12070971
APA StyleHedtke, S. M., Choi, Y.-J., Kode, A., Chalasani, G. C., Sirwani, N., Jada, S. R., Hotterbeekx, A., Mandro, M., Siewe Fodjo, J. N., Amambo, G. N., Abong, R. A., Wanji, S., Kuesel, A. C., Colebunders, R., Mitreva, M., & Grant, W. N. (2023). Assessing Onchocerca volvulus Intensity of Infection and Genetic Diversity Using Mitochondrial Genome Sequencing of Single Microfilariae Obtained before and after Ivermectin Treatment. Pathogens, 12(7), 971. https://doi.org/10.3390/pathogens12070971