Abstract
Microplastics (MPs) and nanoplastics (NPs) are hotspots for the exchange of antimicrobial resistance genes (ARGs) between different bacterial taxa in the environment. Propagation of antimicrobial resistance (AMR) is a global public health issue that needs special attention concerning horizontal gene transfer (HGT) under micro-nano plastics (MNPs) pressure. Interactions between MNPs and microbes, or mere persistence of MNPs in the environment (either water or soil), influence microbial gene expressions, affecting autochthonous microbiomes, their resistomes, and the overall ecosystem. The adsorption of a range of co-contaminants on MNPs leads to the increased interaction of pollutants with microbes resulting in changes in AMR, virulence, toxin production, etc. However, accurately estimating the extent of MNP infestation in agroecosystems remains challenging. The main limitation in estimating the level of MNPs contamination in agroecosystems, surface and subsurface waters, or sediments is the lack of standardized protocols for extraction of MPs and analytical detection methods from complex high organic content matrices. Nonetheless, recent advances in MPs detection from complex matrices with high organic matter content are highly promising. This review aims to provide an overview of relevant information available to date and summarize the already existing knowledge about the mechanisms of MNP-microbe interactions including the different factors with influence on HGT and AMR. In-depth knowledge of the enhanced ARGs propagation in the environment under the influence of MNPs could raise the needed awareness, about future consequences and emergence of multidrug-resistant bacteria.
1. Introduction
The term “microplastics” (MPs) was first coined by Thompson in 2004 to describe the small (less than 5 mm in size) plastic fragments in the oceans []. Since then, MPs from almost all parts of the ecosystem have been reported extensively, indicating possible deleterious consequences on living systems. The early research on MPs focused more on the estimation of the levels of contamination in the ecosystem and understanding the fate of such plastic components of different sizes, shapes, or types in marine and terrestrial animal tissues [,,]. A growing number of recent studies explain the physiological threat posed by MPs [,,]. With the gradual infestation of agricultural soil by MPs, the impact of MPs on agroecosystems [,], groundwater resources [,], bioaccumulation of MPs in plants, and trophic transfer to the food chain via food crops is gaining increased attention []. MPs in aqueous environments or agroecosystems can easily migrate throughout the ecosystem, causing more complex environmental problems. Reports show that MPs in agricultural soil could affect soil pH, respiration, and enzymatic activities depending on MP shape and polymer type []. Some recent articles about MPs in agroecosystems [,,,] shed light on many aspects such as sources of MPs, their transformation, transport, effects to agricultural soils, etc., which have raised concerns about persistence of MNPs in ecosystem and a range of their negative effects such as other anthropogenic pollutants, such as changes in microbial community structure, resistome, and horizontal gene transfer (HGT), which are discussed in this review. Plastic materials have become essential agronomic components for farmers and growers and are used in a wide range of applications as presented in Table 1. Consequently, the route of entry for MPs in agroecosystems are through agricultural fertilizers, agricultural plastic film mulch, sewage/irrigation pipes, etc., and sometimes accidental release of MPs, e.g., through the application of sewage sludge. However, upon long-term use, such plastic materials undergo aging and degradation mostly due to ultraviolet light or other environmental factors such as rain, storm, and wind that accelerate the formation of MPs []. Nanoplastics (NPs) are of sizes less than 100 nm, which typically originate as a result of further degradation of MPs and are more interactive with living systems due to their extremely small sizes and larger surface areas []. Primary sources of micro-nanoplastics (MNPs) are the micro- or nano-sized particles frequently utilized in biomedical and several other consumer application products, while secondary source of MPs results from unregulated disposal of macro-plastic wastes and their subsequent physicochemical and biological degradation in water or soil. Over several years of transformed agricultural practices, plastic components such as protective cultivation films, irrigation/drainage pipes, or protective nets, etc., have helped farmers boost crop productivity while minimizing operational costs, and have become essential agronomic components [].

Table 1.
Major use and application of plastics in agriculture. (adopted from []).
Several recent studies describe the interactions between MPs and microbes in soil and aqueous environments [,,]. Microbes in the environment aggregate on different organic or inorganic surfaces, such as natural environmental components, including leaves or rocks, etc. In addition, as emerging anthropogenic contaminants, MPs are found to act as excellent abiotic surface for attachment of microbial communities. Consequently, MPs in terrestrial and aquatic ecosystems can drive changes in the whole microbiome structure and may modulate different essential nutrient and biogeochemical cycles in the ecosystem [,]. Environmental infestation by MPs has been implicated with another critical public health problem—antimicrobial resistance (AMR). AMR has emerged as a global challenge for human health, agriculture, and animal husbandry. Although resistance development is a multifaceted problem, the environment plays a key role in facilitating resistance evolution and transmission between bacterial species. One major concern for the environmental contamination with MPs is its potential to accelerate microbial interactions and activities by providing suitable surface for attachment of microbes, especially pathogens. Such a process can be driven by a number of mechanisms acting alone or in combination []. One such mechanism is attributed to the high surface area to volume ratios provided by MPs, allowing multiple bacterial species to attach and grow, eventually leading to biofilm formation []; compared with the surrounding environment, bacterial communities (BC) in these “plastisphere” cohabit in close proximity and at higher density, enabling a more efficient exchange of genetic materials. In certain situations, the enrichment of antimicrobial resistance genes (ARGs) and mobile genetic elements (MGEs) in MP biofilms may facilitate the HGT mechanisms []. Moreover, the adsorption of antibiotics and heavy metals on MP surfaces can expose BCs to higher concentrations of these factors, leading to cross-resistance (resistance to antibiotics and metals) or co-resistance (resistance to multiple antibiotics) []. Thus, antimicrobial resistance genes (ARGs) and MNPs both are emerging contaminants in the environment, which are of great public concern due to their potential risk to human health. Figure 1 depicts the route of entry of MPs into the agricultural soil and how it ultimately becomes a health hazard. The aim of this review is to synthesize available information relevant to understanding the interaction of MNPs with the microbiome in agroecosystems affecting microbial resistome and their role in the enhancement of AMR propagation. This is an important issue that came into focus recently and deserved immediate attention to prevent future complications. This review aims to fill in the knowledge gaps and highlight key future research areas in this field with special attention to the factors enhancing the interactions between MNPs, environmental ARGs, and microbes in the soil.
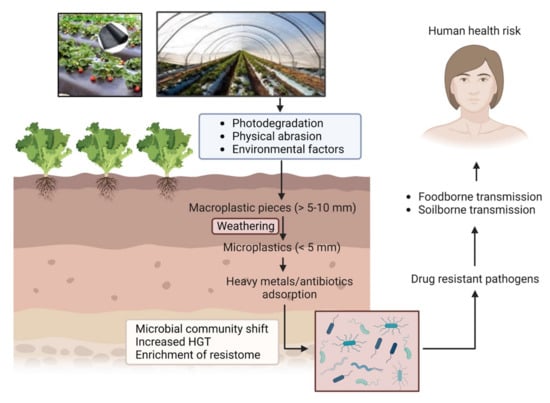
Figure 1.
Pathway of MP/NP mediated antimicrobial resistance through agroecosystem.
2. The Physiologic Implications of Microbe–MNP Interactions
2.1. MNP-Induced Gene Expressions and Gene Transfers
Microorganisms stimulate multiple physiological responses in the presence of MNPs, such as enhanced production of intracellular reactive oxygen species (ROS), increased membrane permeability, and enhanced expression of certain genes. Physical and environmental factors such as size of MNPs, UV-aging, and leachates from MNP degradation are reported for stimulations of such reactions in microbes [,,]. MNPs, especially ~100 nm or smaller, can interact with the microbial surface receptors causing structural changes in the membrane, which allows the internalization of MNPs through endocytosis and ROS generation by NADPH oxidase [,]. A stronger interaction may lead to higher ROS production; however, the viability of both recipient bacteria and donor bacteria decreases significantly with the overproduction of ROS. Higher gene transfer efficiency due to increased ROS response are reported unless the ROS production was so high that it caused excessive damage to bacteria [,]. MNPs have been reported to increase the expressions of genes related to conjugation (trbBp and traF), global regulatory genes (korA, korB, and trbA), outer membrane genes (ompA and ompC), etc. Recent transcriptomics studies showed up-regulation of genes controlling ARG conjugation (kilA and traI), cell permeability (ompR), and intracellular ROS production (recA) upon exposure to aged MPs relative to pristine MP exposures, contributing to enhanced HGT [,,]. All these studies indicated conjugation and transduction as the dominant mechanism for HGT [].
2.2. MP-Associated Biofilms and Cluster Formation
MPs existing in several different forms, such as fibers, films, foams, or pellets, may serve as effective abiotic surfaces for microbial colonization due to their small size but bigger surface area, surface texture, hydrophobic composition, and long persistence. Recent studies have detected different forms of plastics and measured more biomass formation on MP-biofilm than on rock surfaces in river/lake environments [,,]. Plastics-associated microbial communities were found to be distinctly different from those on the non-plastic substrates []. Biofilm formation is known to be a dynamic process, which generally involves microbial adhesion, followed by the secretion of extracellular polymeric substances (EPS) and then microbial proliferation. Microbiota associated with MPs, over time, become the platform for association of other microbes and pathogens, which many times facilitates interaction and transfer of antimicrobial resistance genes (ARGs) among microbes. The expression of quorum-sensing genes by the bacteria accumulated on a surface is also reported to cause further cell-to-cell adhesion, maturation, and dispersion of biofilms []. Quorum sensing also plays a crucial role in microbial communication that helps in the initial attachment of microbes to the MP surface and further proliferation of biofilm formation, leading to increased bacterial resistance. The EPS secreted by adhering microbes on MPs is a viscous layer that enhances the adsorption of many environmental pollutants, such as toxic organic compounds, xenobiotic compounds, heavy metals, etc. []. The formation of biofilms on MPs depends on several physical and chemical properties of plastic substrates, such as polymer type, surface properties, and particle sizes. For example, polyvinylchloride (PVC) and polyethylene (PE) MPs favored enhanced bacterial adhesion more than polypropylene (PP) and polyethylene terephthalate (PET) MPs due to their larger specific surface area and roughness []. The surface properties of MPs significantly influence the colonization by bacterial cells on them. Some studies indicate that differences in the attachment and compositions of bacterial communities were surface-hydrophobicity-dependent on MPs [,]. Surface charges and sizes of MPs and MNPs also influence microbial attachment and biofilm formations, which are discussed in the following sections of this paper. The growth of biofilm on MPs changes their density, influencing their mobility in the environment (water or soil), or often leads to partial degradation, thereby enhance the release of leachates and toxic substances into the environment. Bacteria associated with biofilms often create defense colonies against antimicrobial agents or harsh environments for their survival, which helps in the spread of AMR []. Certain biofilm-associated microbes, such as Pseudomonas aeruginosa, are reported to show increased expression levels of various efflux pumps leading to the release of resistance genes into the environment and enhanced AMR []. Metagenomic sequencing and meta-transcriptomic sequencing methods reveal the enhanced transcriptional activity of several ARGs in MP-associated biofilms or plastisphere []. The role of plastic types was elucidated, such as PET plastics associated with biofilm were found to influence the ARG composition significantly []. Encapsulation of biofilms or colonizing of microbes on MP surfaces acts as a reservoir of microbial pathogens and becomes the hotspot for HGT among the biofilm-forming microbes or other microbes through various MGEs, ARGs [].
3. Influence of MNPs in the Propagation of AMR in Agroecosystem
Recent studies have highlighted the role of MNPs in enhancing the mobility and transfer of ARGs and metal resistance genes (MRGs) in the ecosystem, causing the enhanced spread of AMR. Any agent or situation promoting AMR is considered a threat to global public health. A growing body of scientific evidence correlates the elevation of AMR indicators (i.e., HGT, the abundance of ARGs, MGEs) in the presence of persistent MNPs in an ecosystem. However, elaborated information on underlying mechanisms and experimental evidence on the interaction of MNPs with microbes causing the propagation of AMR is still naive. Table 2 summarizes some of the recent research papers (with a brief description of the work done and main findings) updating the current state of knowledge in this area. The conjugative transfer or HGT of antibiotic resistance genes in E. coli are reported to be highly dependent on the size of MPs []. UV-aged polystyrene microplastics (PS-MPs) were found to increase the HGT of ARGs in E. coli []. MP/NPs were reported to promote the propagation of ARGs in phosphorus-removing bacteria and induced microbial community shift []. MPs also demonstrated selective inhibition of ammonia-oxidizing bacteria and enrichment of nitrite-oxidizing bacteria, leading to partial nitrification []. These publications reported similar effects in bacterial systems upon exposure to MPs, such as increased ROS generation, increased cell permeability, and upregulation of HGT-associated genes.
Table 2.
A list of representative studies on the interaction of MNPs with microbes leading to the HGT and enhanced AMR.
3.1. Impacts on Resistome
The reports on the interaction of MNPs and microbial communities to date strongly show that MPs could highly influence the microbial resistome or changes in the resistome of environmental niches. Several other reports provide evidence that changes in the resistome profile of different groups of microbes due to the presence of MNPs could further enhance the propagation of AMR genes in the environmental compartments as a whole [,].
3.1.1. In the Wastewater Treatment System
Several studies have reported changes in resistome of wastewater sludge systems (which contain high content of ARGs, MNPs, and other contaminants. Studies reported shifts in microbial community structure and phosphorus removal efficiency of wastewater effluent treatment systems, under MPs pressure []. The sludge community structure, detected by high-throughput sequencing technology, showed that the abundance of dominant phyla was considerably shifted in the presence of MPs, which led to profound changes in resistome. Other groups of microbes replaced the more abundant polyphosphate accumulating organisms in sludge; however, the phosphorus removal efficiency was not significantly affected. Similarly, the effects of MPs on the nitrification of aerobic granular sludge (AGS) and shifts in the microbial communities due to an increased abundance of intracellular and extracellular ARGs are reported []. The relative abundance of Nitrosomonas decreased significantly, while the abundance of Nitrospira and other nitrifying bacteria showed an increase. In another study, microbial community composition and abundance of ARGs and MRGs on the plastisphere borne biofilm and planktonic bacteria in wastewater treatment plant (by qPCR) revealed a higher abundance of potentially pathogenic bacteria in treated wastewater on the plastisphere than in the planktonic bacterial community [].
3.1.2. In the Water Ecosystem
MPs in aquatic bodies provide bacterial niches, which serve as hotspot for dissemination of ARGs, and influence the resistome. A clear variation for ARG profiles was elucidated through metagenomic studies between river water and MPs from the water []. Some special ARGs were detected in MPs-associated biofilm, but not in the surrounding river water, indicating selective enrichment and changes in resistome []. Twelve ARGs/MGEs, including four genes responsible for resistance to tetracycline and ampicillin were detected in sewage water in a study, which provided significant information about the combined disposal of MPs, ARGs, and pharmaceuticals in domestic sewage []. Comparison between bacterial taxa, specially pathogenic bacteria and ARGs, on the plastisphere and water revealed the extensive influence of urbanization on the selectivity of microbes and ARGs for MPs []. Metagenomic insights into the ARGs and their hosts on biodegradable and non-biodegradable PET-based MPs suggested selective preferences for two types of plastics harboring ARGs and influencing the resistome in a coastal lagoon in the northern Gulf of Mexico [].
3.1.3. In Soil
Soil is the largest reservoir for several environmental contaminants, which include antibiotic-resistant genes or other mobile genetic elements. Selective enrichment of ARGs or specific groups of bacteria, particularly pathogenic microbes on plastisphere, may increase or contribute more to the pathogenicity by antibiotic resistance gene transfer, making it difficult to kill pathogens that have acquired the ARGs. Overall higher relative abundances of 102 ARGs and 3 MGEs are observed in the plastisphere present in soil compared to the soil environment itself through DNA extraction from soil and sequencing []. Similarly, soil DNA extraction followed by high-throughput sequencing showed a significantly higher number of total ARGs in the plastisphere compared to the surrounding soil for all MP types and sizes considered in the study []. Higher ARGs abundance on larger and aged MPs in the soil after ten years of crop planting is detected using a high-throughput fluorescence quantitative reaction platform []. The changing patterns of ARGs and the bacterial communities on different types and sizes of MPs in soil environments through metagenomic approaches are reported [].
3.2. Impacts on the HGT
Factors influencing the dissemination and proliferation of ARGs in the presence of MPs have come into focus recently. The surrounding environment has been considered an important factor influencing the attached communities compared to the polymer type []. Specific bacterial assemblages are also now known to play a crucial role in ARGs’ persistence and proliferation on the plastisphere, since close and stable relations between ARGs and bacterial taxa have been identified []. The abundance of antibiotics in the environment due to misuse of antibiotics is another important factor for the selection and persistence of ARGs in the environment []. The leachates from MP degradation in soil or due to natural aging may exert additional pressure on enhancement of AMR in opportunistic pathogens over a long time span. As also reported by [], the presence of a range of potential pathogens and ARGs/MGEs in the plastisphere enhances a high risk of ARGs entering potential pathogens. Although it is not clear whether the residual antibiotic drives the selection of the associated resistant gene, it is more likely that antibiotic pressure contributes to the emergence of co -resistance since some types of ARGs may be co-selected []. Some specific factors regarding the interaction of MPs and microbes that have been reported to the date to be responsible for proliferation of AMR are discussed in more detail below. Figure 2 shows a pictorial representation of the factors leading to enhanced ARGs transfer into pathogenic bacteria.
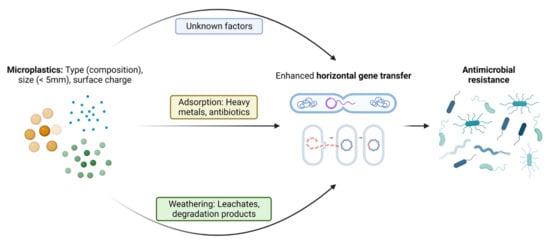
Figure 2.
Pictorial representation of the crucial factors in MNPs-mediated enhancement of the horizontal transfer of intracellular and extracellular antibiotic-resistant genes among microbes.
3.2.1. Size of MPs
As listed in Table 2, the influence of MPs on gene transfer efficiency is reported to be dependent on their sizes []. The relationship between the size distribution of MPs and its impact on gene transfer is an important factor and must be understood in detail. In previous study, smaller MPs (<50 nm) were reported to increase the gene transfer; however, at very high concentrations (>100 mg/L), they exert a negative effect on cell viability and the mechanism of gene transfer []. Moderate-sized MPs (~100 nm) increased the gene transfer efficiency significantly with an increased concentration of MPs. Large MPs (>1 µm) caused no significant increase in gene transfer efficiency []. It is to be noted that the large plastic particles (hundreds of micrometers or at least >5 µm) provide a broad aggregation surface for bacteria to grow on, which in many publications has been referred to as plastisphere. When different types of bacteria attach to the plastisphere surface, the contact between those bacteria becomes more stable, which ultimately results in increased gene transfer efficiency. Several researchers have demonstrated that ARGs can also adsorb directly on the surface of extremely large MPs (hundreds of micrometer scales), accelerating the dissemination of antibiotic-resistance genes in the environment [,]. However, for MPs of sizes smaller than bacteria or near about similar, it is impossible for bacteria to grow on the MPs surface, and such MPs could enhance the gene transfer by inducing more ROS production and increasing cell membrane permeability. The large MPs do not influence the gene transfer because they fail to change these factors and just support stable contact between microbes with MPs and many other contaminants such as heavy metals. Increase of tetracycline resistance gene abundances under MP pressure was found higher than under NP pressure; this difference is potentially due to a stronger selective pressure from the inhibition of growth and metabolism of resistant microorganisms by NPs, resulting from more leaching of chemical additives and increased ease to pass-through biological barriers [].
3.2.2. Type and Concentration of MPs
The nitrification performance of the aerobic granular sludge (AGS) was inhibited by different concentrations and different types of MPs []. Different types of MPs, such as PVC, PE, polyamide (PA), and polystyrene (PS), differentially affected the ammonia removal efficiency and the nitrification function of AGS. Furthermore, the concentration of MPs influenced the nitrification function of AGS, with the nitrification inhibited upon increasing the concentration from 1.0 mg/L to 10 mg/L. Accordingly, nitrification-function-related genes were found to be affected differently by different concentrations of MPs. Although bio-adhesion was observed on the surface of all MPs, it was more pronounced on PE []. The lowest MP concentration at which an effect on gene transfer efficiency is observed is ~0.1 mg/L; however, the maximum concentration is limited by the low solubility of MPs as very high concentration of MPs may cause technical problems, such as causing flocculent precipitates and may interfere with results []. Therefore, 100 mg/L was usually set as the upper limit for the concentration of MPs in most reported studies.
3.2.3. Aging of MPs
As listed in Table 2, the UV-aged MPs, obtained by exposing MPs to UV for up to 20 days, were found to increase the gene transfer frequency relative to the pristine MP without any treatment []. UV irradiation was found to substantially alter the surface chemistry and morphology of MPs, like the natural aging process. For pristine non-rinsed MPs, HGT enhancement was mainly caused by proximal adsorption of ARG donors and recipients, while the contribution from the leachates from MPs was marginal. For aged non-rinsed MPs, both adsorption and leachate contributed to the enhanced ARG transfer frequency, and the latter became more predominant with increased aging. Leachates from aged MPs exert higher oxidative stress on bacteria, thus inducing more intracellular ROS and an increase in cell permeability than those from pristine MPs. While pristine MPs interact with biomolecules mainly via hydrophobic attraction and π-π stacking, the aged MPs with more oxygen-containing functional groups (e.g., C–O-OH) on the surface may form more stable hydrogen bonds with the ARG vectors. A stronger interactive force contributed to the enhanced adsorption capacity of non-rinsed UV-aged MPs towards both ARG donors and recipients. Along with increased retention of ARG donors and recipients, they also increased cell permeability relative to unexposed controls. All these factors contribute to the enhancement of HGT.
4. Impacts of Long-Term Persistence of MPs on Agroecosystem
The persistence of MNPs in an ecosystem can disrupt the delicate balance that exists between its various components. In agroecosystems, MNPs can significantly affect the soil microbial biomass, microbial activity, and functional diversity []. MNPs can also alter the essential nutrient cycles (plant nutrient elements distribution), which are reported to affect plant seed germination or seedling growth indirectly []. Additionally, MPs can block some of the important initial plant life-cycle functions, such as gas exchange and water absorption, along with a potential impact on soil functions by changing the interactions between soil organisms and the surrounding environment. MPs that persist in agroecosystems for a long period gradually undergo aging, which involves leaching out of additives/plasticizers and eventually breaking down into NPs. Various factors, including environmental effects, photodegradation, and/or soil microbial enzyme-mediated degradation, are identified to promote the aging of MPs []. In a study conducted on agricultural farmland soil, ARG abundance on MPs was found to be dependent on MP size, weathering, and years of cultivation [].
Adsorption of Co-Contaminants
Though MPs are classified as emerging pollutants, they may not be hazardous per se to living beings in their native chemical compositions in a pristine microenvironment. However, their omnipresence in the environment is a matter of concern owing to their role as a vector for several hazardous agents, including heavy metals [], rare earth elements [], pharmaceuticals (antibiotics) [,], personal care products [,], and a variety of other emerging contaminants of concern including microbes (antibiotic-resistant bacteria, viruses, etc.) []. Figure 3 shows a pictorial representation of such a long-term natural process amplifying the toxicity of MPs in agroecosystems. The vector hypothesis has led to forecast MPs contamination in the food chain, which poses a hazard to the living world by increasing the risk of outbreaks of food-borne diseases []. The extent of adsorption and desorption of contaminants on MPs depend on the properties of both MPs and contaminants, such as hydrophobicity, surface area, and surface functional groups [,] and the MPs frequently tend to selectively accumulate certain contaminants, resulting in very high local concentration even though the concentrations in the environment are below detection limits []. The interactions between MPs and other pollutants are driven by weak intermolecular forces such as hydrogen bonding, van der Waals force, electrostatic interaction [], pore filling, and π-π interactions [,]. The pH [,], viscosity, salinity [], ionic strength of the media, presence or absence of dissolved organic matter [], diffusion of intraparticle and films, photodegradation, and mechanical stress, are also known to influence the adsorption of contaminants on MPs []. The mobility of pollutants in water or soil matrix increases to varying degrees depending on the amount of adsorption of contaminants onto MPs []. Therefore, pollutant-adsorbed MPs may cause severe disruptions in the agroecosystems by increasing the risks of ecotoxicity and bioaccumulation of toxic agents, having deleterious effects on living systems in the environment [].
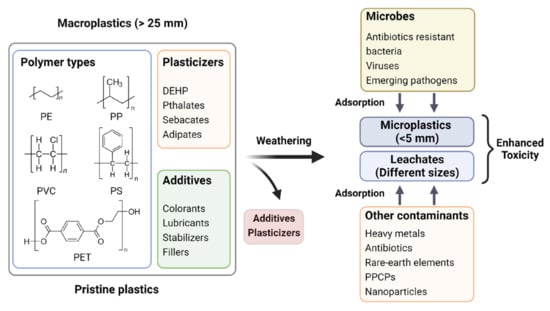
Figure 3.
Increased toxicity of MPs/MP-leachates in agroecosystem upon aging and gradual adsorption/accumulation of co-contaminants.
5. Rapid Detection, Identification, and Quantification of MPs from Environmental Matrices
The currently available methods for separating and detecting MNPs from a complex matrix such as soil have several limitations, restricting them mainly to qualitative enumeration (for MPs) or estimation of total mass (for NPs). Scientists worldwide are actively engaged in developing standardized protocols for separating MPs from the soil matrix and their identification []. Techniques currently adopted by researchers for the isolation of MNPs (of different types/states/nature) from different complex environmental matrices are summarized in Table 3. To detect and quantify MPs, a variety of techniques are currently employed alone or in combination, which includes infra-red spectroscopy (ATR-FTIR, micro-FTIR), Raman spectroscopy, electron microscopy, and mass-spectrometry coupled with gas chromatography.
Table 3.
Frequently used methodologies for detecting MPs/NPs from complex environmental matrices.
5.1. Methodological Challenges
MPs are heterogeneous in nature; they can be formed by different polymers and present with various densities, shapes, and structures. Additionally, they often contain additives and are found in different aging states. These properties of MPs make their extraction, detection and quantification in complex matrices challenging. The smaller size of NPs further prevents their sampling and efficient detection from any complex matrix by techniques commonly employed for MPs detection. Furthermore, the optical detection of NPs is diffraction limited, impeding their analysis below the size of 300 nm. Moreover, when compared with MPs, the smaller size of the NPs poses further restrictions in the availability of reference materials []. Sampling and extraction of MPs from a complex matrix such as agricultural soil is commonly carried out by digestion of biological materials by Fenton oxidation treatment, followed by centrifugation at 3700 rpm for 10 min, and finally visualization of extracted MPs is performed by Nile red staining and fluorescence microscopy []. A number of other techniques are described for MNP extraction, including sieving [], filtration and dialysis [], ultrasonication [], pressurized solvent extraction (PSE) [,], and chemical digestion of organic materials [,]. Each of these extraction techniques has their strengths and limitations, and therefore, the selection of an optimal method for a specific sample should be guided by the sample characteristics. Sample preparation and handling (prevention of contamination) is a huge challenge for reliable detection of MPs. Variation in sample preparation influences the recovery of MPs after extraction from complex matrices. Recent study has compared different sample preparation methods, highlighting their effect on average recoveries of MPs from water samples []. A novel Small Volume Glass Separator and the related standard operating procedures have been developed to overcome the lack of consistency in MPs recovery []. The detection and analysis of extracted MNPs are commonly performed by pyrolysis gas chromatography-mass spectrometry (py-GC-MS), and Fourier transform infrared spectroscopy (FTIR)/Raman spectroscopy [,,,,]. These techniques have greatly improved the detection limits of MPs and NPs; even with these sensitive techniques, residual organic matter/minerals can cause difficulty in the detection; thus, a suitable extraction plays a critical role in the analysis [].
5.2. Recent Developments in Analytical Methods for MNP Detection
In recent years, a number of new techniques, alone or in combination with existing techniques, are increasingly used for MNP analysis. Some of these techniques are specialized ATR-FTIR with hyperspectral imaging, GC-ToF with thermo-gravitimetric analysis [], Laser directed infrared (LDIR) [,], and micro-FTIR [], which are often coupled with existing visualization techniques such as scanning/transmission electron microscopy (SEM/TEM-EDS) [], stereomicroscopy [], and X-ray photoelectron microscopy []. Each of these techniques is limited by a set of advantages and disadvantages. For example, Raman imaging being insensitive to water provides a greater resolution in aquatic sample visualization, but there can be fluorescence interference from background organic–inorganic residues and additives. There is also a risk of overestimation or underestimation when MNPs form aggregates with natural particles. Moreover, setting up parameters such as laser wavelength and magnification can be tricky [,]. Using currently available technologies, reliable detection of each individual MPs sized 4–5 µm are possible; however, only mass concentration can be obtained for NPs of ≤1.0 µm. Some modern techniques utilize UV-Vis method to quantify NPs from different organic matters with non-preferential adsorption of the particles to geothite and resulting in increased specific absorbance []; liquid chromatography-mass spectrometry (LC-MS) is often used for detection of MNPs with limit of detection (LOD) up to 2.0 µg/L and 1.2 µg/L for influents and effluents respectively []. Ref. [] used multi-functional, light-powered, and self-propelled MXene-derived γ-Fe2O3/Pt/TiO2 microrobots for NP detection. They can trap NPs via magnetic attraction with their stacks and enable on-site electrochemical detection and screening. The use of a fluorescent molecular rotor for NP detection in radish sprouts and mussel tissues has been performed, paving the way for new perspectives to overcome challenges in this field []. Researchers around the world are focusing on developing more automated detection techniques and standardization of already-developed protocols as more sensitive and higher-resolution technologies are needed to meet the current challenges in the detection and analysis of MNPs [].
6. Conclusions
MPs provide a suitable surface for attachment of microbes and the formation of biofilms. The slimy nature of biofilm plays a major role in the subsequent attachment and adsorption of many other pollutants and pathogenic microbes, thereby becoming a hub or hotspot for gene transfer. Such close interactions among microbes, and between microbes and pollutants, greatly influence the resistome, thereby enhancing the spread of AMR. Several other properties of MPs, particularly their chemical composition, surface charge, or size, play an important role in gene transfer efficiency between microbes. Moreover, strongly aged MPs had higher ARG abundance compared to weakly aged ones. In agricultural soil, the persistence of MPs and their interaction with soil microbes also influence soil nutrient balance and crop productivity. This review highlighted several factors that have been studied and reported, specifically focusing on the mechanism of horizontal dissemination of ARGs, changes in resistome profile, and the resultant impact on the agroecosystem. However, this area of research is only in its initial phase, and we strongly believe that several other important factors may be yet to be identified. The existing body of research on the impact of MPs in the ecosystem shows that MPs may enter the food chain in several ways and affect human health. However, current research is still in the stage of exploration. Furthermore, accurate quantification of MPs in complex matrices from environmental samples is impacted by the lack of appropriate techniques and standard protocols for extraction and detection of MNPs. In this review, we summarized the promising recent advancements in the extraction and detection of MNPs. Overall, a deeper insight into MNP contamination in agroecosystems and its impact on different related constituents needs continued and extensive research. It is also crucial that knowledge from these should be utilized to benefit humankind and protect human health by implementing proper control measures for plastic usage in agriculture.
Funding
Partial supports for J.N. and J.D. were from the National Institute of Food and Agriculture (NIFA) of the United States Department of Agriculture (USDA) grants (2020-70020-33033 and 2022-70020-37590) awarded to P.B.
Data Availability Statement
This review used information from published peer-reviewed articles. All data and findings reported in this paper can be found in the respective references.
Acknowledgments
Conflicts of Interest
The authors declare no conflict of interest.
References
- Thompson, R.C.; Olsen, Y.; Mitchell, R.P.; Davis, A.; Rowland, S.J.; John, A.W.; McGonigle, D.; Russell, A.E. Lost at sea: Where is all the plastic? Science 2004, 304, 838. [Google Scholar] [CrossRef]
- Bråte, I.L.N.; Hurley, R.; Iversen, K.; Beyer, J.; Thomas, K.V.; Steindal, C.C.; Green, N.W.; Olsen, M.; Lusher, A. Mytilus spp. as sentinels for monitoring microplastic pollution in Norwegian coastal waters: A qualitative and quantitative study. Environ. Pollut. 2018, 243, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Digka, N.; Tsangaris, C.; Torre, M.; Anastasopoulou, A.; Zeri, C. Microplastics in mussels and fish from the Northern Ionian Sea. Mar. Pollut. Bull. 2018, 135, 30–40. [Google Scholar] [CrossRef]
- Gedik, K.; Eryaşar, A.R. Microplastic pollution profile of Mediterranean mussels (Mytilus galloprovincialis) collected along the Turkish coasts. Chemosphere 2020, 260, 127570. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, T.; Liu, P.; Li, H.; Hu, F. The formation of specific bacterial communities contributes to the enrichment of antibiotic resistance genes in the soil plastisphere. J. Hazard. Mater. 2022, 436, 129247. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Mi, J.; Yan, Q.; Wen, X.; Zhou, S.; Wang, Y.; Ma, B.; Zou, Y.; Liao, X.; Wu, Y. Animal manures application increases the abundances of antibiotic resistance genes in soil-lettuce system associated with shared bacterial distributions. Sci. Total Environ. 2021, 787, 147667. [Google Scholar] [CrossRef] [PubMed]
- Lai, K.P.; Tsang, C.F.; Li, L.; Yu, R.M.K.; Kong, R.Y.C. Microplastics act as a carrier for wastewater-borne pathogenic bacteria in sewage. Chemosphere 2022, 301, 134692. [Google Scholar] [CrossRef]
- Dai, Y.; Shi, J.; Zhang, N.; Pan, Z.; Xing, C.; Chen, X. Current research trends on microplastics pollution and impacts on agro-ecosystems: A short review. Sep. Sci. Technol. 2022, 57, 656–669. [Google Scholar] [CrossRef]
- Isari, E.A.; Papaioannou, D.; Kalavrouziotis, I.K.; Karapanagioti, H.K. Microplastics in Agricultural Soils: A Case Study in Cultivation of Watermelons and Canning Tomatoes. Water 2021, 13, 2168. [Google Scholar] [CrossRef]
- Lang, M.; Wang, G.; Yang, Y.; Zhu, W.; Zhang, Y.; Ouyang, Z.; Guo, X. The occurrence and effect of altitude on microplastics distribution in agricultural soils of Qinghai Province, northwest China. Sci. Total Environ. 2022, 810, 152174. [Google Scholar] [CrossRef]
- Castan, S.; Henkel, C.; Hüffer, T.; Hofmann, T. Microplastics and nanoplastics barely enhance contaminant mobility in agricultural soils. Commun. Earth Environ. 2021, 2, 193. [Google Scholar] [CrossRef]
- Helmberger, M.S.; Tiemann, L.K.; Grieshop, M.J. Towards an ecology of soil microplastics. Funct. Ecol. 2020, 34, 550–560. [Google Scholar] [CrossRef]
- Zhao, T.; Lozano, Y.M.; Rillig, M.C. Microplastics increase soil pH and decrease microbial activities as a function of microplastic shape, polymer type, and exposure time. Front. Environ. Sci. 2021, 9, 675803. [Google Scholar] [CrossRef]
- Ding, L.; Huang, D.; Ouyang, Z.; Guo, X. The effects of microplastics on soil ecosystem: A review. Curr. Opin. Environ. Sci. Health 2022, 26, 100344. [Google Scholar] [CrossRef]
- Joos, L.; De Tender, C. Soil under stress: The importance of soil life and how it is influenced by (micro) plastic pollution. Comput. Struct. Biotechnol. J. 2022, 20, 1554–1566. [Google Scholar] [CrossRef]
- Tian, L.; Jinjin, C.; Ji, R.; Ma, Y.; Yu, X. Microplastics in agricultural soils: Sources, effects, and their fate. Curr. Opin. Environ. Sci. Health 2022, 25, 100311. [Google Scholar] [CrossRef]
- Maity, S.; Guchhait, R.; Sarkar, M.B.; Pramanick, K. Occurrence and distribution of micro/nanoplastics in soils and their phytotoxic effects: A review. Plant Cell Environ. 2022, 45, 1011–1028. [Google Scholar] [CrossRef]
- He, F.; Shi, H.; Liu, R.; Tian, G.; Qi, Y.; Wang, T. Randomly-shaped nanoplastics induced stronger biotoxicity targeted to earthworm Eisenia fetida species: Differential effects and the underlying mechanisms of realistic and commercial polystyrene nanoplastics. Sci. Total Environ. 2023, 877, 162854. [Google Scholar] [CrossRef]
- Pazienza, P.; De Lucia, C. For a new plastics economy in agriculture: Policy reflections on the EU strategy from a local perspective. J. Clean. Prod. 2020, 253, 119844. [Google Scholar] [CrossRef]
- Scarascia-Mugnozza, G.; Sica, C.; Russo, G. Plastic materials in European agriculture: Actual use and perspectives. J. Agric. Eng. 2011, 42, 15–28. [Google Scholar] [CrossRef]
- Battulga, B.; Kawahigashi, M.; Oyuntsetseg, B. Characterization of biofilms formed on polystyrene microplastics (PS-MPs) on the shore of the Tuul River, Mongolia. Environ. Res. 2022, 212, 113329. [Google Scholar] [CrossRef] [PubMed]
- Sagawa, N.; Kawaai, K.; Hinata, H. Abundance and size of microplastics in a coastal sea: Comparison among bottom sediment, beach sediment, and surface water. Mar. Pollut. Bull. 2018, 133, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Ma, J.; Li, G.; Rillig, M.C.; Zhu, Y.-G. Soil plastispheres as hotspots of antibiotic resistance genes and potential pathogens. ISME J. 2022, 16, 521–532. [Google Scholar] [CrossRef]
- Xu, B.; Liu, F.; Cryder, Z.; Huang, D.; Lu, Z.; He, Y.; Wang, H.; Lu, Z.; Brookes, P.C.; Tang, C.; et al. Microplastics in the soil environment: Occurrence, risks, interactions and fate–a review. Crit. Rev. Environ. Sci. Technol. 2020, 50, 2175–2222. [Google Scholar] [CrossRef]
- Rillig, M.C.; Leifheit, E.; Lehmann, J. Microplastic effects on carbon cycling processes in soils. PLoS Biol. 2021, 19, e3001130. [Google Scholar] [CrossRef] [PubMed]
- Junaid, M.; Liu, X.; Wu, Y.; Wang, J. Selective enrichment of antibiotic resistome and bacterial pathogens by aquatic microplastics. J. Hazard. Mater. Adv. 2022, 7, 100106. [Google Scholar] [CrossRef]
- Wu, X.; Pan, J.; Li, M.; Li, Y.; Bartlam, M.; Wang, Y. Selective enrichment of bacterial pathogens by microplastic biofilm. Water Res. 2019, 165, 114979. [Google Scholar] [CrossRef]
- Zhang, G.; Chen, J.; Li, W. Conjugative antibiotic-resistant plasmids promote bacterial colonization of microplastics in water environments. J. Hazard. Mater. 2022, 430, 128443. [Google Scholar] [CrossRef]
- Imran, M.; Das, K.R.; Naik, M.M. Co-selection of multi-antibiotic resistance in bacterial pathogens in metal and microplastic contaminated environments: An emerging health threat. Chemosphere 2019, 215, 846–857. [Google Scholar] [CrossRef]
- Yuan, Q.; Sun, R.; Yu, P.; Cheng, Y.; Wu, W.; Bao, J.; Alvarez, P.J. UV-aging of microplastics increases proximal ARG donor-recipient adsorption and leaching of chemicals that synergistically enhance antibiotic resistance propagation. J. Hazard. Mater. 2022, 427, 127895. [Google Scholar] [CrossRef]
- Zhao, T.; Tan, L.; Han, X.; Wang, X.; Zhang, Y.; Ma, X.; Lin, K.; Wang, R.; Ni, Z.; Wang, J.; et al. Microplastic-induced apoptosis and metabolism responses in marine Dinoflagellate, Karenia mikimotoi. Sci. Total Environ. 2022, 804, 150252. [Google Scholar] [CrossRef]
- Feng, L.-J.; Sun, X.-D.; Zhu, F.-P.; Feng, Y.; Duan, J.-L.; Xiao, F.; Li, X.-Y.; Shi, Y.; Wang, Q.; Sun, J.-W.; et al. Nanoplastics promote microcystin synthesis and release from cyanobacterial Microcystis aeruginosa. Environ. Sci. Technol. 2020, 54, 3386–3394. [Google Scholar] [CrossRef] [PubMed]
- Zha, Y.; Li, Z.; Zhong, Z.; Ruan, Y.; Sun, L.; Zuo, F.; Li, L.; Hou, S. Size-dependent enhancement on conjugative transfer of antibiotic resistance genes by micro/nanoplastics. J. Hazard. Mater. 2022, 431, 128561. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.-J.; Zeng, Y.-H.; Zhu, J.-M.; Cai, Z.-H.; Zhou, J. The structure and assembly mechanisms of plastisphere microbial community in natural marine environment. J. Hazard. Mater. 2022, 421, 126780. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gao, J.; Dai, H.; Zhao, Y.; Li, D.; Duan, W.; Guo, Y. Microplastics affect the ammonia oxidation performance of aerobic granular sludge and enrich the intracellular and extracellular antibiotic resistance genes. J. Hazard. Mater. 2021, 409, 124981. [Google Scholar] [CrossRef]
- Huang, H.; Feng, G.; Wang, M.; Liu, C.; Wu, Y.; Dong, L.; Feng, L.; Zheng, X.; Chen, Y. Nitric oxide: A neglected driver for the conjugative transfer of antibiotic resistance genes among wastewater microbiota. Environ. Sci. Technol. 2022, 56, 6466–6478. [Google Scholar] [CrossRef]
- Lenaker, P.L.; Corsi, S.R.; Mason, S.A. Spatial distribution of microplastics in surficial benthic sediment of Lake Michigan and Lake Erie. Environ. Sci. Technol. 2020, 55, 373–384. [Google Scholar] [CrossRef]
- Steinman, A.D.; Scott, J.; Green, L.; Partridge, C.; Oudsema, M.; Hassett, M.; Kindervater, E.; Rediske, R.R. Persistent organic pollutants, metals, and the bacterial community composition associated with microplastics in Muskegon Lake (MI). J. Great Lakes Res. 2020, 46, 1444–1458. [Google Scholar] [CrossRef]
- Ogonowski, M.; Motiei, A.; Ininbergs, K.; Hell, E.; Gerdes, Z.; Udekwu, K.I.; Bacsik, Z.; Gorokhova, E. Evidence for selective bacterial community structuring on microplastics. Environ. Microbiol. 2018, 20, 2796–2808. [Google Scholar] [CrossRef]
- Lami, R. Quorum sensing in marine biofilms and environments. In Quorum Sensing; Academic Press: Cambridge, MA, USA, 2019; pp. 55–96. [Google Scholar] [CrossRef]
- Liu, S.; Shi, J.; Wang, J.; Dai, Y.; Li, H.; Li, J.; Liu, X.; Chen, X.; Wang, Z.; Zhang, P. Interactions Between Microplastics and Heavy Metals in Aquatic Environments: A Review. Front. Microbiol. 2021, 12, 652520. [Google Scholar] [CrossRef]
- Cai, L.; Wu, D.; Xia, J.; Shi, H.; Kim, H. Influence of physicochemical surface properties on the adhesion of bacteria onto four types of plastics. Sci. Total Environ. 2019, 671, 1101–1107. [Google Scholar] [CrossRef]
- Xu, X.; Wang, S.; Gao, F.; Li, J.; Zheng, L.; Sun, C.; He, C.; Wang, Z.; Qu, L. Marine microplastic-associated bacterial community succession in response to geography, exposure time, and plastic type in China’s coastal seawaters. Mar. Pollut. Bull. 2019, 145, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Kaur, K.; Reddy, S.; Barathe, P.; Oak, U.; Shriram, V.; Kharat, S.S.; Govarthanan, M.; Kumar, V. Microplastic-associated pathogens and antimicrobial resistance in environment. Chemosphere 2021, 291, 133005. [Google Scholar] [CrossRef]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.-J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Liu, Z.; Li, M.; Bartlam, M.; Wang, Y. Integrated metagenomic and metatranscriptomic analysis reveals actively expressed antibiotic resistomes in the plastisphere. J. Hazard. Mater. 2022, 430, 128418. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Cao, N.; Duan, C.; Wang, Q.; Ding, C.; Wang, J. Selection of antibiotic resistance genes on biodegradable and non-biodegradable microplastics. J. Hazard. Mater. 2021, 409, 124979. [Google Scholar] [CrossRef]
- Abe, K.; Nomura, N.; Suzuki, S. Biofilms: Hot spots of horizontal gene transfer (HGT) in aquatic environments, with a focus on a new HGT mechanism. FEMS Microbiol. Ecol. 2020, 96, fiaa031. [Google Scholar] [CrossRef]
- Zhou, C.-S.; Wu, J.-W.; Liu, B.-F.; Ma, W.-L.; Yang, S.-S.; Cao, G.-L. (Micro) nanoplastics promote the risk of antibiotic resistance gene propagation in biological phosphorus removal system. J. Hazard. Mater. 2022, 431, 128547. [Google Scholar] [CrossRef]
- Wang, X.; Li, H.; Chen, Y.; Meng, X.; Dieketseng, M.Y.; Wang, X.; Yan, S.; Wang, B.; Zhou, L.; Zheng, G. A neglected risk of nanoplastics as revealed by the promoted transformation of plasmid-borne ampicillin resistance gene by Escherichia coli. Environ. Microbiol. 2022, 24, 4946–4959. [Google Scholar] [CrossRef]
- Li, R.; Zhu, L.; Yang, K.; Li, H.; Zhu, Y.-G.; Cui, L. Impact of Urbanization on Antibiotic Resistome in Different Microplastics: Evidence from a Large-Scale Whole River Analysis. Environ. Sci. Technol. 2021, 55, 8760–8770. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, J.; Wu, J.; Wang, J.; Luo, Y. Potential risks of microplastics combined with superbugs: Enrichment of antibiotic resistant bacteria on the surface of microplastics in mariculture system. Ecotoxicol. Environ. Saf. 2020, 187, 109852. [Google Scholar] [CrossRef] [PubMed]
- Galafassi, S.; Sabatino, R.; Sathicq, M.B.; Eckert, E.M.; Fontaneto, D.; Fontana, G.D.; Mossotti, R.; Corno, G.; Volta, P.; Di Cesare, A. Contribution of microplastic particles to the spread of resistances and pathogenic bacteria in treated wastewaters. Water Res. 2021, 201, 117368. [Google Scholar] [CrossRef]
- Xu, C.; Lu, J.; Shen, C.; Wang, J.; Li, F. Deciphering the mechanisms shaping the plastisphere antibiotic resistome on riverine microplastics. Water Res. 2022, 225, 119192. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gao, J.; Zhao, Y.; Dai, H.; Jia, J.; Zhang, D. Plastisphere enrich antibiotic resistance genes and potential pathogenic bacteria in sewage with pharmaceuticals. Sci. Total Environ. 2021, 768, 144663. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Luo, Q.; Zhao, S.; Zhao, P.; Yang, X.; Huang, Q.; Su, J. Watershed urbanization enhances the enrichment of pathogenic bacteria and antibiotic resistance genes on microplastics in the water environment. Environ. Pollut. 2022, 313, 120185. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.-M.; Lu, P.-Z.; Liu, X.-P. Fate and abundance of antibiotic resistance genes on microplastics in facility vegetable soil. Sci. Total Environ. 2020, 709, 136276. [Google Scholar] [CrossRef] [PubMed]
- Basili, M.; Quero, G.M.; Giovannelli, D.; Manini, E.; Vignaroli, C.; Avio, C.G.; De Marco, R.; Luna, G.M. Major role of surrounding environment in shaping biofilm community composition on marine plastic debris. Front. Mar. Sci. 2020, 7, 262. [Google Scholar] [CrossRef]
- Su, Y.; Zhang, Z.; Zhu, J.; Shi, J.; Wei, H.; Xie, B.; Shi, H. Microplastics act as vectors for antibiotic resistance genes in landfill leachate: The enhanced roles of the long-term aging process. Environ. Pollut. 2021, 270, 116278. [Google Scholar] [CrossRef]
- Zhao, R.; Yu, K.; Zhang, J.; Zhang, G.; Huang, J.; Ma, L.; Deng, C.; Li, X.; Li, B. Deciphering the mobility and bacterial hosts of antibiotic resistance genes under antibiotic selection pressure by metagenomic assembly and binning approaches. Water Res. 2020, 186, 116318. [Google Scholar] [CrossRef]
- Wiedner, K.; Polifka, S. Effects of microplastic and microglass particles on soil microbial community structure in an arable soil (Chernozem). Soil 2020, 6, 315–324. [Google Scholar] [CrossRef]
- Bosker, T.; Bouwman, L.J.; Brun, N.R.; Behrens, P.; Vijver, M.G. Microplastics accumulate on pores in seed capsule and delay germination and root growth of the terrestrial vascular plant Lepidium sativum. Chemosphere 2019, 226, 774–781. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Hu, G.; Fan, X.; Jia, H. Sorption properties of cadmium on microplastics: The common practice experiment and A two-dimensional correlation spectroscopic study. Ecotoxicol. Environ. Saf. 2020, 190, 110118. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Yang, C.; Zhu, Z.; Bai, X.; Ma, J. Adsorption behavior of organic pollutants and metals on micro/nanoplastics in the aquatic environment. Sci. Total Environ. 2019, 694, 133643. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, J.; Zhou, B.; Zhou, Y.; Dai, Z.; Zhou, Q.; Christie, P.; Luo, Y. Enhanced adsorption of oxytetracycline to weathered microplastic polystyrene: Kinetics, isotherms and influencing factors. Environ. Pollut. 2018, 243 Pt B, 1550–1557. [Google Scholar] [CrossRef]
- Fan, X.; Zou, Y.; Geng, N.; Liu, J.; Hou, J.; Li, D.; Yang, C.; Li, Y. Investigation on the adsorption and desorption behaviors of antibiotics by degradable MPs with or without UV ageing process. J. Hazard. Mater. 2021, 401, 123363. [Google Scholar] [CrossRef]
- Ho, W.-K.; Law, J.C.-F.; Lo, J.C.-W.; Chng, I.K.-X.; Hor, C.H.-H.; Leung, K.S.-Y. Sorption Behavior, Speciation, and Toxicity of Microplastic-Bound Chromium in Multisolute Systems. Environ. Sci. Technol. Lett. 2023, 10, 27–32. [Google Scholar] [CrossRef]
- Pompei, C.M.E.; Campos, L.C.; da Silva, B.F.; Fogo, J.C.; Vieira, E.M. Occurrence of PPCPs in a Brazilian water reservoir and their removal efficiency by ecological filtration. Chemosphere 2019, 226, 210–219. [Google Scholar] [CrossRef]
- Fang, S.; Yu, W.; Li, C.; Liu, Y.; Qiu, J.; Kong, F. Adsorption behavior of three triazole fungicides on polystyrene microplastics. Sci. Total Environ. 2019, 691, 1119–1126. [Google Scholar] [CrossRef]
- Zarfl, C.; Matthies, M. Are marine plastic particles transport vectors for organic pollutants to the Arctic? Mar. Pollut. Bull. 2010, 60, 1810–1814. [Google Scholar] [CrossRef]
- Elizalde-Velázquez, A.; Subbiah, S.; Anderson, T.A.; Green, M.J.; Zhao, X.; Cañas-Carrell, J.E. Sorption of three common nonsteroidal anti-inflammatory drugs (NSAIDs) to microplastics. Sci. Total Environ. 2020, 715, 136974. [Google Scholar] [CrossRef]
- Yamashita, R.; Tanaka, K.; Yeo, B.G.; Takada, H.; van Franeker, J.A.; Dalton, M.; Dale, E. Hazardous Chemicals in Plastics in Marine Environments: International Pellet Watch; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Puckowski, A.; Cwięk, W.; Mioduszewska, K.; Stepnowski, P.; Białk-Bielińska, A. Sorption of pharmaceuticals on the surface of microplastics. Chemosphere 2021, 263, 127976. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhang, M.; Sha, W.; Wang, Y.; Hao, H.; Dou, Y.; Li, Y. Sorption Behavior and Mechanisms of Organic Contaminants to Nano and Microplastics. Molecules 2020, 25, 1827. [Google Scholar] [CrossRef]
- Xiao, F.; Zhang, X.; Penn, L.; Gulliver, J.S.; Simcik, M.F. Effects of Monovalent Cations on the Competitive Adsorption of Perfluoroalkyl Acids by Kaolinite: Experimental Studies and Modeling. Environ. Sci. Technol. 2011, 45, 10028–10035. [Google Scholar] [CrossRef]
- Bundschuh, M.; Weyers, A.; Ebeling, M.; Elsaesser, D.; Schulz, R. Narrow pH Range of Surface Water Bodies Receiving Pesticide Input in Europe. Bull. Environ. Contam. Toxicol. 2016, 96, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, M.V.; Carter, L.J.; Agatz, A.; Boxall, A.B. Novel Approach for Characterizing pH-Dependent Uptake of Ionizable Chemicals in Aquatic Organisms. Environ. Sci. Technol. 2017, 51, 6965–6971. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Zheng, M.; Wang, L.; Zhao, Q.; Lou, Y.; Shi, L.; Qu, L. Sorption of polyhalogenated carbazoles (PHCs) to microplastics. Mar. Pollut. Bull. 2019, 146, 718–728. [Google Scholar] [CrossRef]
- Xu, B.; Liu, F.; Brookes, P.C.; Xu, J. The sorption kinetics and isotherms of sulfamethoxazole with polyethylene microplastics. Mar. Pollut. Bull. 2018, 131, 191–196. [Google Scholar] [CrossRef]
- Joo, S.H.; Liang, Y.; Kim, M.; Byun, J.; Choi, H. Microplastics with adsorbed contaminants: Mechanisms and Treatment. Environ. Chall. 2021, 3, 100042. [Google Scholar] [CrossRef]
- Zhao, L.; Rong, L.; Xu, J.; Lian, J.; Wang, L.; Sun, H. Sorption of five organic compounds by polar and nonpolar microplastics. Chemosphere 2020, 257, 127206. [Google Scholar] [CrossRef]
- Seidensticker, S.; Grathwohl, P.; Lamprecht, J.; Zarfl, C. A combined experimental and modeling study to evaluate pH-dependent sorption of polar and non-polar compounds to polyethylene and polystyrene microplastics. Environ. Sci. Eur. 2018, 30, 30. [Google Scholar] [CrossRef]
- Mári, Á.; Bordós, G.; Gergely, S.; Büki, M.; Háhn, J.; Palotai, Z.; Besenyő, G.; Szabó, É.; Salgó, A.; Kriszt, B.; et al. Validation of microplastic sample preparation method for freshwater samples. Water Res. 2021, 202, 117409. [Google Scholar] [CrossRef]
- Grause, G.; Kuniyasu, Y.; Chien, M.-F.; Inoue, C. Separation of microplastic from soil by centrifugation and its application to agricultural soil. Chemosphere 2022, 288, 132654. [Google Scholar] [CrossRef] [PubMed]
- Bouzid, N.; Anquetil, C.; Dris, R.; Gasperi, J.; Tassin, B.; Derenne, S. Quantification of microplastics by pyrolysis coupled with gas chromatography and mass spectrometry in sediments: Challenges and implications. Microplastics 2022, 1, 229–239. [Google Scholar] [CrossRef]
- De Frond, H.; Hampton, L.T.; Kotar, S.; Gesulga, K.; Matuch, C.; Lao, W.; Weisberg, S.B.; Wong, C.S.; Rochman, C.M. Monitoring microplastics in drinking water: An interlaboratory study to inform effective methods for quantifying and characterizing microplastics. Chemosphere 2022, 298, 134282. [Google Scholar] [CrossRef]
- Lv, L.; He, L.; Jiang, S.; Chen, J.; Zhou, C.; Qu, J.; Lu, Y.; Hong, P.; Sun, S.; Li, C. In situ surface-enhanced Raman spectroscopy for detecting microplastics and nanoplastics in aquatic environments. Sci. Total Environ. 2020, 728, 138449. [Google Scholar] [CrossRef]
- Mowla, M.; Shakiba, S.; Louie, S.M. Selective quantification of nanoplastics in environmental matrices by asymmetric flow field-flow fractionation with total organic carbon detection. Chem. Commun. 2021, 57, 12940–12943. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, R.; Tsuchiya, M.; Lindsay, D.J.; Kitahashi, T.; Fujikura, K.; Fukushima, T. A new small device made of glass for separating microplastics from marine and freshwater sediments. PeerJ 2019, 7, e7915. [Google Scholar] [CrossRef] [PubMed]
- Okoffo, E.D.; Ribeiro, F.; O’Brien, J.W.; O’Brien, S.; Tscharke, B.J.; Gallen, M.; Samanipour, S.; Mueller, J.F.; Thomas, K.V. Identification and quantification of selected plastics in biosolids by pressurized liquid extraction combined with double-shot pyrolysis gas chromatography–mass spectrometry. Sci. Total Environ. 2020, 715, 136924. [Google Scholar] [CrossRef]
- Primpke, S.; Fischer, M.; Lorenz, C.; Gerdts, G.; Scholz-Böttcher, B.M. Comparison of pyrolysis gas chromatography/mass spectrometry and hyperspectral FTIR imaging spectroscopy for the analysis of microplastics. Anal. Bioanal. Chem. 2020, 412, 8283–8298. [Google Scholar] [CrossRef]
- Saliu, F.; Biale, G.; Raguso, C.; La Nasa, J.; Degano, I.; Seveso, D.; Galli, P.; Lasagni, M.; Modugno, F. Detection of plastic particles in marine sponges by a combined infrared micro-spectroscopy and pyrolysis-gas chromatography-mass spectrometry approach. Sci. Total Environ. 2022, 819, 152965. [Google Scholar] [CrossRef]
- Sullivan, G.L.; Gallardo, J.D.; Jones, E.W.; Hollliman, P.J.; Watson, T.M.; Sarp, S. Detection of trace sub-micron (nano) plastics in water samples using pyrolysis-gas chromatography time of flight mass spectrometry (PY-GCToF). Chemosphere 2020, 249, 126179. [Google Scholar] [CrossRef]
- Weckhuysen, B.; ten Have, I.; Meirer, F.; Oord, R.; Zettler, E.; Van Sebille, E.; Amaral-Zettler, L. Nanoscale Infrared Spectroscopy Reveals Nanoplastics at 5000 m Depth in the South Atlantic Ocean. Phys. Sci. 2021; preprint. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Y.; Qiu, Z.; He, Y.; Zhang, Y. Towards a fast and generalized microplastic quantification method in soil using terahertz spectroscopy. Sci. Total Environ. 2022, 841, 156624. [Google Scholar] [CrossRef]
- Urso, M.; Ussia, M.; Novotný, F.; Pumera, M. Trapping and detecting nanoplastics by MXene-derived oxide microrobots. Nat. Commun. 2022, 13, 3573. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Chen, Y.; Ouyang, X.; Weng, L.; Ma, J.; Islam, S.; Li, Y. Resolving natural organic matter and nanoplastics in binary or ternary systems via UV–Vis analysis. J. Colloid Interface Sci. 2023, 632, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Yu, E.-S.; Jeong, E.T.; Lee, S.; Kim, I.S.; Chung, S.; Han, S.; Choi, I.; Ryu, Y.-S. Real-Time Underwater Nanoplastic Detection beyond the Diffusion Limit and Low Raman Scattering Cross-Section via Electro-Photonic Tweezers. ACS Nano 2022, 17, 2114–2123. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Skoczynska, E.; van Putten, R.-J.; Leslie, H.A.; Gruter, G.-J.M. Quantification of polyethylene terephthalate micro-and nanoplastics in domestic wastewater using a simple three-step method. Sci. Total Environ. 2023, 857, 159209. [Google Scholar] [CrossRef]
- Moraz, A.; Breider, F. Detection and Quantification of Nonlabeled Polystyrene Nanoparticles Using a Fluorescent Molecular Rotor. Anal. Chem. 2021, 93, 14976–14984. [Google Scholar] [CrossRef]
- Ivleva, N.P. Chemical analysis of microplastics and nanoplastics: Challenges, advanced methods, and perspectives. Chem. Rev. 2021, 121, 11886–11936. [Google Scholar] [CrossRef]
- Jia, W.; Karapetrova, A.; Zhang, M.; Xu, L.; Li, K.; Huang, M.; Wang, J.; Huang, Y. Automated identification and quantification of invisible microplastics in agricultural soils. Sci. Total Environ. 2022, 844, 156853. [Google Scholar] [CrossRef]
- Prata, J.C.; da Costa, J.P.; Girão, A.V.; Lopes, I.; Duarte, A.C.; Rocha-Santos, T. Identifying a quick and efficient method of removing organic matter without damaging microplastic samples. Sci. Total Environ. 2019, 686, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Amin, A.; Gibson, C.T.; Chuah, C.; Tang, Y.; Naidu, R.; Fang, C. Raman imaging of microplastics and nanoplastics generated by cutting PVC pipe. Environ. Pollut. 2022, 298, 118857. [Google Scholar] [CrossRef] [PubMed]
- Cella, C.; La Spina, R.; Mehn, D.; Fumagalli, F.; Ceccone, G.; Valsesia, A.; Gilliland, D. Detecting micro-and nanoplastics released from food packaging: Challenges and analytical strategies. Polymers 2022, 14, 1238. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Liu, J.; Yu, J.; Zhou, Q.; Wu, W.; Fu, L.; Yin, C.; Fernandez, C.; Karimi-Maleh, H. Current development and future challenges in microplastic detection techniques: A bibliometrics-based analysis and review. Sci. Prog. 2022, 105, 00368504221132151. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).