
Superantigens and SARS-CoV-2


{kind=link}
Abstract
1. What Is a Superantigen?
2. Lessons from Dengue
3. Superantigens and T-Cell Dysfunction
4. Superantigens and Autoimmunity
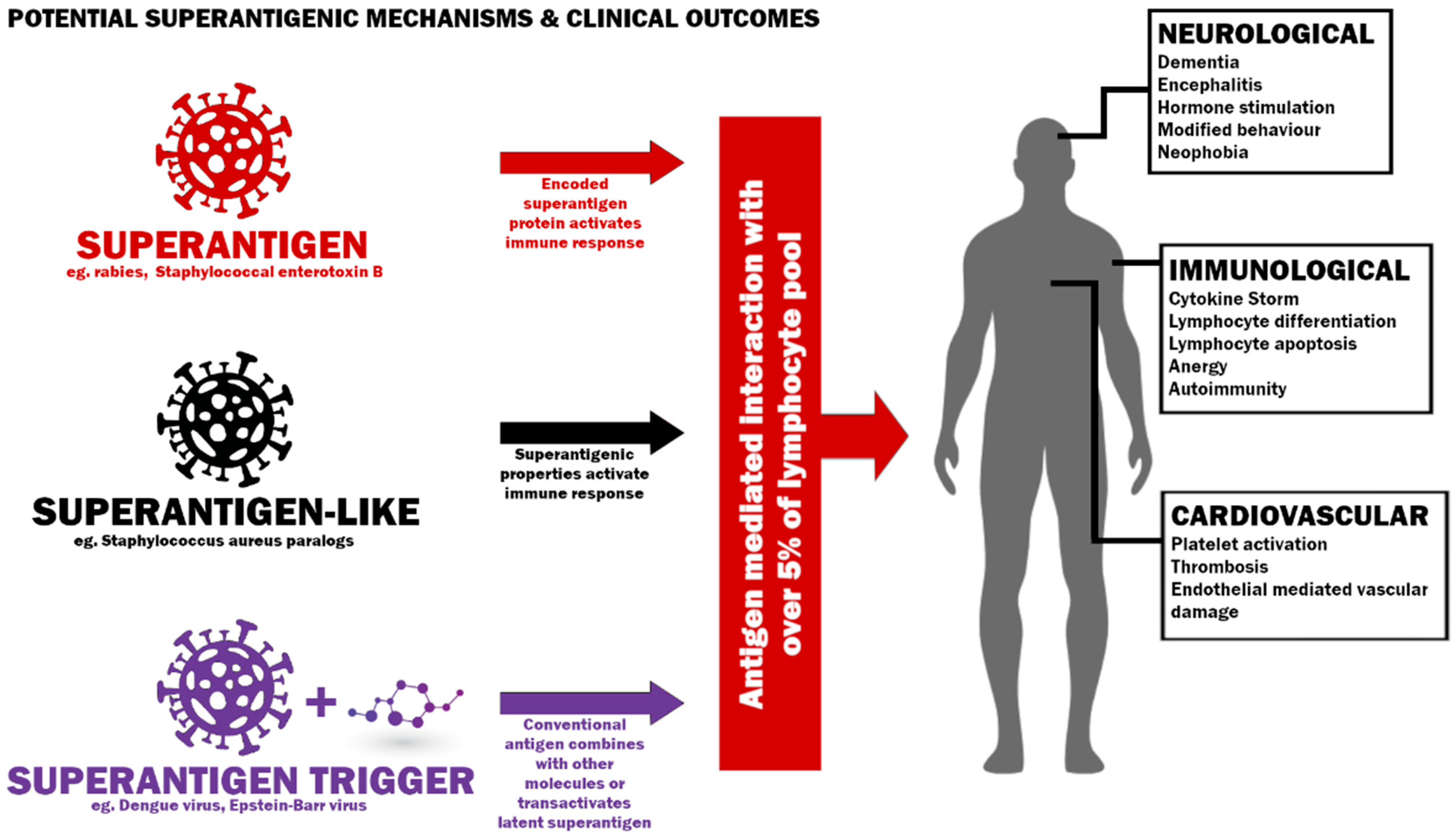
5. SARS-CoV-2 as a Superantigenic, Superantigen-like Pathogen or Superantigen Trigger
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- White, J.; Herman, A.; Pullen, A.M.; Kubo, R.; Kappler, J.W.; Marrack, P. The V Beta-Specific Superantigen Staphylococcal Enterotoxin B: Stimulation of Mature T Cells and Clonal Deletion in Neonatal Mice. Cell 1989, 56, 27–35. [Google Scholar] [CrossRef]
- Pascual, V.; Capra, J.D. B-Cell Superantigens? Curr. Biol. 1991, 1, 315–317. [Google Scholar] [CrossRef]
- Silverman, G.J.; Goodyear, C.S. Confounding B-Cell Defences: Lessons From a Staphylococcal Superantigen. Nat. Rev. Immunol. 2006, 6, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Lafferty, J.A.; Clements, J.R.; Todd, J.K.; Gelfand, E.W.; Kappler, J.; Marrack, P.; Kotzin, B.L. Selective Expansion of T Cells Expressing V Beta 2 in Toxic Shock Syndrome. J. Exp. Med. 1990, 172, 981–984. [Google Scholar] [CrossRef]
- Kotzin, B.L.; Leung, D.Y.M.; Kappler, J.; Marrack, P. Superantigens and Their Potential Role in Human Disease. Adv. Immunol. 1993, 54, 99–166. [Google Scholar] [CrossRef]
- Kotb, M. Bacterial pyrogenic exotoxins as superantigens. Clin. Microbiol. Rev. 1995, 8, 411–426. [Google Scholar] [CrossRef]
- Ericsson, P.O.; Hansson, J.; Widegren, B.; Dohlsten, M.; Sjogren, H.O.; Hedlund, G. In vivo induction of gamma/delta T cells with highly potent and selective anti-tumor cytotoxicity. Eur. J. Immunol. 1996, 21, 2797–2802. [Google Scholar] [CrossRef]
- McCormack, J.E.; Callahan, J.E.; Kappler, J.; Marrack, P.C. Profound deletion of mature T-cells in vivo by chronic exposure to exogenous superantigen. J. Immunol. 1993, 150, 3785–3792. [Google Scholar]
- Janik, D.K.; Lee, W.T. Staphylococcal Enterotoxin B (SEB) Induces Memory CD4 T Cell Anergy in vivo and Impairs Recall Immunity to Unrelated Antigens. J. Clin. Cell Immunol. 2015, 6, 1–8. [Google Scholar] [CrossRef]
- Guedez, Y.; Norrby-Teglund, A.; Low, D.; McGeer, A.; Kotb, M. HLA class II alleles associated with outcome of invasive group A streptococcal infections. In Proceedings of the 37th Annual Meeting of the Interscience Conference on Antimicrobial Agents and Chemotherapy, Toronto, ON, Canada, 29 September–1 October 1997. [Google Scholar]
- Nadal, D.; Lauener, R.P.; Braegger, C.P.; Kaufhold, A.; Simma, B.; Lutticken, R.; Seger, R.A. T-cell activation and cytokine release in streptococcal toxic shock-like syndrome. J. Pediatr. 1993, 122, 727–729. [Google Scholar] [CrossRef]
- Emmer, A.; Gerlach, K.; Staege, M.S.; Kornhuber, M.E. Superantigen-Mediated Encephalitis; Hayasaka, D., Ed.; InTech: Rijeka, Croatia, 2011. [Google Scholar] [CrossRef]
- Kornhuber, M.E.; Emmer, A.; Gerlach, K.; Staege, M.S. Experimental Models of Superantigen-Mediated Neuropathology. In Superantigens; Kotb, M., Fraser, J.D., Eds.; ASM Press: Washington, DC, USA, 2007. [Google Scholar] [CrossRef]
- Urbach-Ross, D.; Kusnecov, A.W. Impact of superantigenic molecules on central nervous system function. Front. Biosci. 2009, 14, 4416–4426. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Brogan, P.A.; Shah, V.; Klein, N.; Dillon, M.J. Vbeta-restricted T cell adherence to endothelial cells: A mechanism for superantigen-dependent vascular injury. Arthritis Rheum. 2004, 50, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Kulhankova, K.; Kinney, K.J.; Stach, J.M.; Gourronc, F.A.; Grumbach, I.A.; Klingelhutz, A.J.; Salgado-Pabon, W. The Superantigen Toxic Shock Syndrome Toxin 1 Alters Human Aortic Endothelial Cell Function. Infect. Immun. 2018, 86, e00848-17. [Google Scholar] [CrossRef]
- Sundstrom, M.; Abrahmsen, L.; Antonsson, P.; Mehindate, K.; Mourad, W.; Dohlsten, M. The crystal structure of staphylococcal enterotoxin type D reveals Zn2+-mediated homodimerization. EMBO J. 1996, 15, 6832–6840. [Google Scholar] [CrossRef]
- Jardetzky, T.; Brown, J.; Gorga, J.; Stern, L.; Urban, R.; Chi, Y.; Stauffacher, C.; Strominger, J.; Wiley, D. Three-dimensional structure of a human class II histocompatibility molecule complexed with superantigen. Nature 1994, 368, 711–718. [Google Scholar] [CrossRef]
- Kim, J.; Urban, R.; Strominger, J.; Wiley, D. Toxic shock syndrome toxin-1 complexed with a class II major histocompatibility molecule HLA-DR1. Science 1994, 266, 1870–1874. [Google Scholar] [CrossRef]
- Al-Shangiti, A.M.; Naylor, C.E.; Nair, S.P.; Briggs, D.C.; Henderson, B.; Chain, B.M. Structural relationships and cellular tropism of staphylococcal superantigen-like proteins. Infect. Immun. 2004, 72, 4261–4270. [Google Scholar] [CrossRef]
- Armstrong, P.C.; Hu, H.; Rivera, J.; Rigby, S.; Chen, Y.C.; Howden, B.P.; Gardiner, E.; Peter, K. Staphylococcal superantigen-like protein 5 induces thrombotic and bleeding complications in vivo: Inhibition by an anti-SSL5 antibody and the glycan Bimosiamose. J. Thromb. Haemost. JTH 2012, 10, 2607–2609. [Google Scholar] [CrossRef]
- Hu, H.; Armstrong, P.C.; Khalil, E.; Chen, Y.C.; Straub, A.; Li, M.; Soosairajah, J.; Hagemeyer, C.E.; Bassler, N.; Huang, D.; et al. GPVI and GPIbα mediate staphylococcal superantigen-like protein 5 (SSL5) induced platelet activation and direct toward glycans as potential inhibitors. PLoS ONE 2011, 6, e19190. [Google Scholar] [CrossRef]
- Gagnon, S.J.; Leporati, A.; Green, S.; Kalayanarooj, S.; Vaughn, D.W.; Stephens, H.A.; Suntayakorn, S.; Kurane, I.; Ennis, F.A.; Rothman, A.L. T cell receptor Vbeta gene usage in Thai children with dengue virus infection. Am. J. Trop. Med. Hyg. 2001, 64, 41–48. [Google Scholar] [CrossRef][Green Version]
- Carod-Artal, F.J.; Wichmann, O.; Farrar, J.; Gascón, J. Neurological complications of dengue virus infection. Lancet Neurol. 2013, 12, 906–919. [Google Scholar] [CrossRef]
- Wan, S.-W.; Lin, C.-F.; Yeh, T.-M.; Liu, C.-C.; Liu, H.-S.; Wang, S.; Ling, P.; Anderson, R.; Lei, H.-Y.; Lin, Y.-S. Autoimmunity in dengue pathogenesis. J. Formos. Med. Assoc. 2013, 112, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Qiu, Y.; Liu, H.; Liang, B.; Fan, B.; Zhou, X.; Liu, D. Transcription profile of human endogenous retroviruses in response to dengue virus serotype 2 infection. Virology 2020, 544, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Katzourakis, A.; Tristem, M. Phylogeny of human endogenous and exogenous retroviruses. Retrovir. Primate Genome Evol. 2005, 186, 203. [Google Scholar]
- Nelson, P.N.; Carnegie, P.R.; Martin, J.; Davari Ejtehadi, H.; Hooley, P.; Roden, D.; Rowland-Jones, S.; Warren, P.; Astley, J. Demystified. Human endogenous retroviruses. Mol Pathol. 2003, 56, 11–18. [Google Scholar] [CrossRef]
- Posso-Osorio, I.; Tobón, G.J.; Cañas, C.A. Human endogenous retroviruses (HERV) and non-HERV viruses incorporated into the human genome and their role in the development of autoimmune diseases. J. Transl. Autoimmun. 2021, 4, 100137. [Google Scholar] [CrossRef]
- Posnett, D.N.; Yarilina, A.A. Sleeping with the enemy—Endogenous superantigens in humans. Immunity 2001, 15, 503–506. [Google Scholar] [CrossRef]
- Sutkowski, N.; Conrad, B.; Thorley-Lawson, D.; Huber, B.T. Epstein-Barr virus transactivates the human endogenous retrovirus HERV-K18 that encodes a superantigen. Immunity 2001, 15, 579–589. [Google Scholar] [CrossRef]
- Sicat, J.; Sutkowski, N.; Huber, B.T. Expression of human endogenous retrovirus HERV-K18 superantigen is elevated in juvenile rheumatoid arthritis. J. Rheumatol. 2005, 32, 1821–1831. [Google Scholar]
- Wang, M.; Wang, L.; Liu, H.; Chen, J.; Liu, D. Transcriptome Analyses Implicate Endogenous Retroviruses Involved in the Host Antiviral Immune System through the Interferon Pathway. Virol. Sin. 2021, 36, 1315–1326. [Google Scholar] [CrossRef]
- Temerozo, J.; Fintelman-Rodrigues, N.; Santos, M.C.; Hottz, E.; Sacramento, C.; Silva, A.; Mandacaru, S.; Moraes, E.C.; Trugilho, M.; Gesto, J.; et al. Human Endogenous Retrovirus K Activation in the Lower Respiratory Tract of Severe COVID-19 Patients Associates with Early Mortality. Res. Sq. 2021, 21, 514541. [Google Scholar] [CrossRef]
- Balestrieri, E.; Minutolo, A.; Petrone, V.; Fanelli, M.; Iannetta, M.; Malagnino, V.; Zordan, M.; Vitale, P.; Charvet, B.; Horvat, B.; et al. Evidence of the pathogenic HERV-W envelope expression in T lymphocytes in association with the respiratory outcome of COVID-19 patients. EBioMedicine 2021, 66, 103341. [Google Scholar] [CrossRef] [PubMed]
- Charvet, B.; Brunel, J.; Pierquin, J.; Iampietro, M.; Decimo, D.; Queruel, N.; Lucas, A.; Encabo-Berzosa, M.; Arenaz, I.; Marmolejo, T.P.; et al. SARS-CoV-2 induces human endogenous retrovirus type W envelope protein expression in blood lymphocytes and in tissues of COVID-19 patients. medRxiv 2022. Available online: https://www.medrxiv.org/content/10.1101/2022.01.18.21266111v2.full.pdf (accessed on 7 February 2022).
- Perron, H.; Jouvin-Marche, E.; Michel, M.; Ounanian-Paraz, A.; Camelo, S.; Dumon, A.; Jolivet-Reynaud, C.; Marcel, F.; Souillet, Y.; Borel, E.; et al. Multiple sclerosis retrovirus particles and recombinant envelope trigger an abnormal immune response in vitro, by inducing polyclonal Vβ16 T-lymphocyte activation. Virology 2001, 287, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Deacy, A.M.; Gan, S.K.-E.; Derrick, J.P. Superantigen Recognition and Interactions: Functions, Mechanisms and Applications. Front. Immunol. 2021, 12, 1845. [Google Scholar] [CrossRef]
- Miethke, T.; Wahl, C.; Heeg, K.; Wagner, H. Acquired resistance to superantigen-induced T cell shock. V beta selective T cell unresponsiveness unfolds directly from a transient state of hyperreactivity. J. Immunol. 1993, 150, 3776–3784. [Google Scholar]
- Kuroda, K.; Yagi, J.; Imanishi, K.; Yan, X.J.; Li, X.Y.; Fujimaki, W.; Kato, H.; Miyoshi-Akiyama, T.; Kumazawa, Y.; Abe, H.; et al. Implantation of IL-2-containing osmotic pump prolongs the survival of superantigen-reactive T cells expanded in mice injected with bacterial superantigen. J. Immunol. 1996, 157, 1422–1431. [Google Scholar]
- Meilleur, C.E.; Wardell, C.M.; Mele, T.S.; Dikeakos, J.D.; Bennink, J.R.; Mu, H.; McCormick, J.K.; Mansour Haeryfar, S.M. Bacterial Superantigens Expand and Activate, Rather than Delete or Incapacitate, Preexisting Antigen-Specific Memory CD8+ T Cells. J. Infect. Dis. 2019, 219, 1307–1317. [Google Scholar] [CrossRef]
- Whiteside, S.K.; Snook, J.P.; Williams, M.A.; Weis, J.J. Bystander T Cells: A Balancing Act of Friends and Foes. Trends Immunol. 2018, 39, 1021–1035. [Google Scholar] [CrossRef]
- Tough, D.F.; Borrow, P.; Sprent, J. Induction of bystander T cell proliferation by viruses and type I interferon in vivo. Science 1996, 272, 1947–1950. [Google Scholar] [CrossRef]
- Coppola, M.A.; Blackman, M.A. Bacterial superantigens reactivate antigen-specific CD8+ memory T cells. Int. Immunol. 1997, 9, 1393–1403. [Google Scholar] [CrossRef] [PubMed]
- Biasi, G.; Panozzo, M.; Pertile, P.; Mezzalira, S.; Facchinetti, A. Mechanism underlying superantigen-induced clonal deletion of mature T lymphocytes. Int. Immunol. 1994, 6, 983–989. [Google Scholar] [CrossRef] [PubMed]
- Ehl, S.; Hombach, J.; Aichele, P.; Hengartner, H.; Zinkernagel, R.M. Bystander activation of cytotoxic T cells: Studies on the mechanism and evaluation of in vivo significance in a transgenic mouse model. J. Exp. Med. 1997, 185, 1241–1251. [Google Scholar] [CrossRef] [PubMed]
- Halnon, N.J.; Jamieson, B.; Plunkett, M.; Kitchen, C.M.; Pham, T.; Krogstad, P. Thymic function and impaired maintenance of peripheral T cell populations in children with congenital heart disease and surgical thymectomy. Pediatr. Res. 2005, 57, 42–48. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Palmer, S.; Albergante, L.; Blackburn, C.C.; Newman, T.J. Thymic involution and rising disease incidence with age. Proc. Natl. Acad. Sci. USA 2018, 115, 1883–1888. [Google Scholar] [CrossRef] [PubMed]
- Phetsouphanh, C.; Darley, D.R.; Wilson, D.B.; Howe, A.; Munier, C.M.; Patel, S.K.; Juno, J.A.; Burrell, L.M.; Kent, S.J.; Dore, G.J.; et al. Immunological dysfunction persists for 8 months following initial mild-to-moderate SARS-CoV-2 infection. Nat. Immunol. 2022, 23, 210–216. [Google Scholar] [CrossRef]
- Li, M.; Yao, D.; Zeng, X.; Kasakovski, D.; Zhang, Y.; Chen, S.; Zha, X.; Li, Y.; Xu, L. Age related human T cell subset evolution and senescence. Immun. Ageing 2019, 16, 24. [Google Scholar] [CrossRef]
- Kaech, S.; Wherry, E.; Ahmed, R. Effector and memory T-cell differentiation: Implications for vaccine development. Nat. Rev. Immunol. 2002, 2, 251–262. [Google Scholar] [CrossRef]
- Cicin-Sain, L.; Smyk-Pearson, S.; Currier, N.; Byrd, L.; Koudelka, C.; Robinson, T.; Swarbrick, G.; Tackitt, S.; Legasse, A.; Fischer, M.; et al. Loss of naive T cells and repertoire constriction predict poor response to vaccination in old primates. J. Immunol. 2010, 184, 6739–6745. [Google Scholar] [CrossRef]
- Yarwood, J.M.; Leung, D.Y.; Schlievert, P.M. Evidence for the involvement of bacterial superantigens in psoriasis, atopic dermatitis, and Kawasaki syndrome. FEMS Microbiol. Lett. 2000, 192, 1–7. [Google Scholar] [CrossRef]
- Conrad, B.; Weissmahr, R.N.; Böni, J.; Arcari, R.; Schüpbach, J.; Mach, B. A Human Endogenous Retroviral Superantigen as Candidate Autoimmune Gene in Type I Diabetes. Cell 1997, 90, 303–313, ISSN 0092-8674. [Google Scholar] [CrossRef]
- Friedman, S.M.; Posnett, D.N.; Tumang, J.R.; Cole, B.C.; Crow, M.K. A potential role for microbial superantigens in the pathogenesis of systemic autoimmune disease. Arthritis Rheum. 1991, 34, 468–480. [Google Scholar] [CrossRef] [PubMed]
- Acha-Orbea, H. Bacterial and viral superantigens: Roles in autoimmunity? Ann. Rheum. Dis. 1993, 52, S6–S16. [Google Scholar] [CrossRef] [PubMed]
- Silverman, G.J. B cell superantigens: Possible roles in immunodeficiency and autoimmunity. Semin. Immunol. 1998, 10, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Soos, J.M.; Schiffenbauer, J.; Torres, B.A.; Johnson, H.M. Superantigens as virulence factors in autoimmunity and immunodeficiency diseases. Med. Hypotheses 1997, 48, 253–259. [Google Scholar] [CrossRef]
- Li, H.; Llera, A.; Malchiodi, E.L.; Mariuzza, R.A. The structural basis of T-cell activation by superantigens. Ann. Rev. Immunol. 1999, 17, 435–466. [Google Scholar] [CrossRef] [PubMed]
- Stollerman, G.H. Rheumatogenic group A streptococci and the return of rheumatic fever. Adv. Intern. Med. 1990, 35, 1–25. [Google Scholar]
- Domiati-Saad, R.; Attrep, J.F.; Brezinschek, H.P.; Cherrie, A.H.; Karp, D.R.; Lipsky, P.E. Staphylococcal enterotoxin D functions as a human B-cell superantigen by rescuing VH4-expressing B-cells from apoptosis. J. Immunol. 1996, 156, 3608–3620. [Google Scholar]
- Hofer, M.F.; Newell, K.; Duke, R.C.; Schlievert, P.M.; Freed, J.H.; Leung, D.Y.M. Differential effects of staphylococcal toxic shock syndrome toxin 1 on B cell apoptosis. Proc. Natl. Acad. Sci. USA 1996, 93, 5425–5430. [Google Scholar] [CrossRef]
- Chowdhary, V.R.; Tilahun, A.Y.; Clark, C.R.; Grande, J.P.; Rajagopalan, G. Chronic exposure to staphylococcal superantigen elicits a systemic inflammatory disease mimicking lupus. J. Immunol. 2012, 189, 2054–2062. [Google Scholar] [CrossRef]
- Dar, S.A.; Janahi, E.M.; Haque, S.; Akhter, N.; Jawed, A.; Wahid, M.; Ramachandran, V.G.; Bhattacharya, S.N.; Banerjee, B.D.; Das, S. Superantigen influence in conjunction with cytokine polymorphism potentiates autoimmunity in systemic lupus erythematosus patients. Immunol. Res. 2016, 64, 1001–1012. [Google Scholar] [CrossRef] [PubMed]
- Sekigawa, I.; Ogasawara, H.; Kaneko, H.; Hishikawa, T.; Hashimoto, H. Retroviruses and autoimmunity. Intern. Med. 2001, 40, 80–86. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ogasawara, H.; Naito, T.; Kaneko, H.; Hishikawa, T.; Sekigawa, I.; Hashimoto, H.; Kaneko, Y.; Yamamoto, N.; Maruyama, N.; Yamamoto, N. Quantitative analyses of messenger RNA of human endogenous retrovirus in patients with systemic lupus erythematosus. J. Rheumatol. 2001, 28, 533–538. [Google Scholar] [PubMed]
- Kitsou, K.; Kotanidou, A.; Paraskevis, D.; Karamitros, T.; Katzourakis, A.; Tedder, R.; Hurst, T.; Sapounas, S.; Kotsinas, A.; Gorgoulis, V.; et al. Upregulation of Human Endogenous Retroviruses in Bronchoalveolar Lavage Fluid of COVID-19 Patients. Microbiol. Spectr. 2021, 9, e01260-21. [Google Scholar] [CrossRef]
- Levet, S.; Medina, J.; Joanou, J.; Demolder, A.; Queruel, N.; Reant, K.; Normand, M.; Seffals, M.; Dimier, J.; Germi, R.; et al. An ancestral retroviral protein identified as a therapeutic target in type-1 diabetes. JCI Insight 2017, 2, 17. [Google Scholar] [CrossRef] [PubMed]
- Niegowska, M.; Wajda-Cuszlag, M.; Stepien-Ptak, G.; Trojanek, J.; Michałkiewicz, J.; Szalecki, M.; Sechi, L. Anti-HERV-WEnv antibodies are correlated with seroreactivity against Mycobacterium avium subsp. paratuberculosis in children and youths at T1D risk. Sci. Rep. 2019, 9, 6282. [Google Scholar] [CrossRef]
- Pan, T.; Cao, G.; Tang, E.; Zhao, Y.; Penaloza-MacMaster, P.; Fang, Y.; Huang, J. A single-cell atlas reveals shared and distinct immune responses and metabolism during SARS-CoV-2 and HIV-1 infections. bioRxiv 2022. Available online: https://www.biorxiv.org/content/10.1101/2022.01.10.475725v1.full.pdf (accessed on 7 February 2022).
- Wang, E.Y.; Mao, T.; Klein, J.; Dai, Y.; Huck, J.D.; Jaycox, J.R.; Liu, F.; Zhou, T.; Israelow, B.; Wong, P.; et al. Diverse functional autoantibodies in patients with COVID-19. Nature 2021, 595, 283–288. [Google Scholar] [CrossRef]
- Muratori, P.; Lenzi, M.; Muratori, L.; Granito, A. Antinuclear antibodies in COVID 19. Clin. Transl. Sci. 2021, 14, 1627–1628. [Google Scholar] [CrossRef]
- Kouo, T.; Chaisawangwong, W. SARS-CoV-2 as a superantigen in multisystem inflammatory syndrome in children. J. Clin. Investig. 2021, 131, e149327. [Google Scholar] [CrossRef]
- Rha, M.S.; Shin, E.C. Activation or exhaustion of CD8+ T cells in patients with COVID-19. Cell Mol. Immunol. 2021, 18, 2325–2333. [Google Scholar] [CrossRef] [PubMed]
- Gottesman, B.L.; Yu, J.; Tanaka, C.; Longhurst, C.A.; Kim, J.J. Incidence of New-Onset Type 1 Diabetes Among US Children During the COVID-19 Global Pandemic. JAMA Pediatr. 2022, 24, 5801. [Google Scholar] [CrossRef] [PubMed]
- Chertow, D.; Stein, S.; Ramelli, S.; Grazioli, A.; Chung, J.; Singh, M.; Yinda, C.K.; Winkler, C.; Dickey, J.; Ylaya, K.; et al. SARS-CoV-2 infection and persistence throughout the human body and brain. Available online: https://www.researchsquare.com/article/rs-1139035/v1. (accessed on 7 February 2022).
- de Melo, G.D.; Lazarini, F.; Levallois, S.; Hautefort, C.; Michel, V.; Larrous, F.; Verillaud, B.; Aparicio, C.; Wagner, S.; Gheusi, G.; et al. COVID-19-related anosmia is associated with viral persistence and inflammation in human olfactory epithelium and brain infection in hamsters. Sci. Transl. Med. 2021, 13, eabf8396. [Google Scholar] [CrossRef] [PubMed]
- Vibholm, L.K.; Nielsen, S.S.F.; Pahus, M.H.; Frattari, G.S.; Olesen, R.; Andersen, R.; Monrad, I.; Andersen, A.H.F.; Thomsen, M.M.; Konrad, C.V.; et al. SARS-CoV-2 persistence is associated with antigen-specific CD8 T-cell responses. EBioMedicine 2021, 64, 103230. [Google Scholar] [CrossRef] [PubMed]
- Gaebler, C.; Wang, Z.; Lorenzi, J.C.C.; Muecksch, F.; Finkin, S.; Tokuyama, M.; Cho, A.; Jankovic, M.; Schaefer-Babajew, D.; Oliveira, T.Y.; et al. Evolution of antibody immunity to SARS-CoV-2. Nature 2021, 591, 639–644. [Google Scholar] [CrossRef]
- Reddy, R.; Farber, N.; Kresch, E.; Seetharam, D.; Diaz, P.; Ramasamy, R. SARS-CoV-2 in the Prostate: Immunohistochemical and Ultrastructural Studies. World J. Men’s Health 2022, 40, e12. [Google Scholar] [CrossRef]
- Martin-Cardona, A.; Lloreta Trull, J.; Albero-González, R.; Beser, M.P.; Andújar, X.; Ruiz-Ramirez, P.; Tur-Martínez, J.; Ferrer, C.; De Marcos Izquierdo, J.A.; Madrigal, A.P.; et al. SARS-CoV-2 identified by transmission electron microscopy in lymphoproliferative and ischaemic intestinal lesions of COVID-19 patients with acute abdominal pain: Two case reports. BMC Gastroenterol. 2021, 21, 334. [Google Scholar] [CrossRef]
- Arostegui, D.; Castro, K.; Schwarz, S.; Vaidy, K.; Rabinowitz, S.; Wallach, T. Persistent SARS-CoV-2 Nucleocapsid Protein Presence in the Intestinal Epithelium of a Pediatric Patient 3 Months After Acute Infection. PGN Rep. 2021, 3, e152. [Google Scholar] [CrossRef]
- Cheng, M.H.; Zhang, S.; Porritt, R.A.; Rivas, M.N.; Paschold, L.; Willscher, E.; Binder, M.; Arditi, M.; Bahar, I. Superantigenic character of an insert unique to SARS-CoV-2 spike supported by skewed TCR repertoire in patients with hyperinflammation. Proc. Natl. Acad. Sci. USA 2020, 117, 25254–25262. [Google Scholar] [CrossRef]
- Hoste, L.; Roels, L.; Naesens, L.; Bosteels, V.; Vanhee, S.; Dupont, S.; Bosteels, C.; Browaeys, R.; Vandamme, N.; Verstaen, K.; et al. TIM3+ TRBV11-2 T cells and IFNγ signature in patrolling monocytes and CD16+ NK cells delineate MIS–C. J. Exp. Med. 2022, 219, e20211381. [Google Scholar] [CrossRef]
- Moreews, M.; Le Gouge, K.; Khaldi-Plassart, S.; Pescarmona, R.; Mathieu, A.L.; Malcus, C.; Djebali, S.; Bellomo, A.; Dauwalder, O.; Perret, M.; et al. Polyclonal expansion of TCR Vb 21.3+ CD4+ and CD8+ T cells is a hallmark of multisystem inflammatory syndrome in children. Sci. Immunol. 2021, 6, eabh1516. [Google Scholar] [CrossRef] [PubMed]
- Brodin, P. SARS-CoV-2 infections in children: Understanding diverse outcomes. Immunity 2022, 55, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.B.; Schwartz, N.G.; Patel, P.; Abbo, L.; Beauchamps, L.; Balan, S.; Lee, E.H.; Paneth-Pollak, R.; Geevarughese, A.; Lash, M.K.; et al. Case Series of Multisystem Inflammatory Syndrome in Adults Associated with SARS-CoV-2 Infection—United Kingdom and United States, March-August 2020. MMWR Morb. Mortal. Wkly. Rep. 2020, 69, 1450–1456. [Google Scholar] [CrossRef] [PubMed]
- Gomard-Mennesson, E.; Landron, C.; Dauphin, C.; Epaulard, O.; Petit, C.; Green, L.; Roblot, P.; Lusson, J.-R.; Broussolle, C.; Sève, P. Kawasaki Disease in Adults. Medicine 2010, 89, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Kontopoulou, T.; Kontopoulos, D.G.; Vaidakis, E.; Mousoulis, G.P. Adult Kawasaki disease in a European patient: A case report and review of the literature. J. Med. Case Rep. 2015, 9, 75. [Google Scholar] [CrossRef] [PubMed]
- Scaglioni, V.; Soriano, E.R. Are superantigens the cause of cytokine storm and viral sepsis in severe COVID-19? Observations and hypothesis. Scand. J. Immunol. 2020, 92, e12944. [Google Scholar] [CrossRef]
- Porritt, R.A.; Paschold, L.; Rivas, M.N.; Cheng, M.H.; Yonker, L.M.; Chandnani, H.; Lopez, M.; Simnica, D.; Schultheiß, C.; Santiskulvong, C.; et al. HLA class I-associated expansion of TRBV11-2 T cells in multisystem inflammatory syndrome in children. J. Clin. Investig. 2021, 131, e146614. [Google Scholar] [CrossRef]
- Gómez-Icazbalceta, G.; Hussain, Z.; Vélez-Alavez, M. In silico evidence of superantigenic features in ORF8 protein from COVID-19. bioRxiv 2021. Available online: https://www.biorxiv.org/content/10.1101/2021.12.14.472240v1.full.pdf (accessed on 7 February 2022).
- Chiappelli, F. Comments on “An insertion unique to SARS-CoV-2 exhibits super antigenic character strengthened by recent mutations” by Cheng MH et al. 2020. Bioinformation 2020, 16, 474–476. [Google Scholar] [CrossRef]
- Cheng, M.H.; Porritt, R.A.; Rivas, M.N. A monoclonal antibody against staphylococcal enterotoxin B superantigen inhibits SARS-CoV-2 entry in vitro. Structure 2021, 29, 951.e3–962.e3. [Google Scholar] [CrossRef]
- Bittmann, S.; Weissenstein, A.; Luchter, E.; Moschüring-Alieva Villalon, G. Multisystem inflammatory syndrome in children (MIS–C): The role of viral superantigens in COVID-19 disease. J. Allergy Infect. Dis. 2020, 1, 18–20. [Google Scholar]
- Tang, Y.; Liu, J.; Zhang, D.; Xu, Z.; Ji, J.; Wen, C. Cytokine Storm in COVID-19: The Current Evidence and Treatment Strategies. Front. Immunol. 2020, 11, 1708. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, D.; Pissarra, R.; Reis-Melo, A.; Guimarães, H. Multisystem inflammatory syndrome in children (MISC): A systematic review. Int. J. Clin. Pract. 2021, 75, e14450. [Google Scholar] [CrossRef] [PubMed]
- Buonsenso, D.; Riitano, F.; Valentini, P. Pediatric Inflammatory Multisystem Syndrome Temporally Related With SARS-CoV-2: Immunological Similarities With Acute Rheumatic Fever and Toxic Shock Syndrome. Front. Pediatr. 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Yeung, R.S.M. The etiology of Kawasaki disease: A superantigen-mediated process. Prog. Pediatr. Cardiol. 2004, 19, 115–122, ISSN 1058-9813. [Google Scholar] [CrossRef]
- Aiyegbusi, O.L.; Hughes, S.E.; Turner, G.; Rivera, S.C.; McMullan, C.; Chandan, J.S.; Haroon, S.; Price, G.; Davies, E.H.; Nirantharakumar, K.; et al. Symptoms, complications and management of long COVID: A review. J. R. Soc. Med. 2021, 114, 428–442. [Google Scholar] [CrossRef]
- Lynall, M. Neuropsychiatric symptoms in lupus. Lupus 2018, 27 (Suppl. 1), 18–20. [Google Scholar] [CrossRef]
- Leuchten, N.; Milke, B.; Winkler-Rohlfing, B.; Daikh, D.; Dörner, T.; Johnson, S.R.; Aringer, M. Early symptoms of systemic lupus erythematosus (SLE) recalled by 339 SLE patients. Lupus 2018, 27, 1431–1436. [Google Scholar] [CrossRef]
- Pettersson, S.; Lövgren, M.; Eriksson, L.E.; Moberg, C.; Svenungsson, E.; Gunnarsson, I.; Henriksson, E.W. An exploration of patient-reported symptoms in systemic lupus erythematosus and the relationship to health-related quality of life. Scand. J. Rheumatol. 2012, 41, 383–390. [Google Scholar] [CrossRef]
- Seeßle, J.; Waterboer, T.; Hippchen, T.; Simon, J.; Kirchner, M.; Lim, A.; Müller, B.; Merle, U. Persistent Symptoms in Adult Patients 1 Year After Coronavirus Disease 2019 (COVID-19): A Prospective Cohort Study. Clin. Infect. Dis. 2021. [Google Scholar] [CrossRef]
- Zong, F.; Gan, C.; Wang, Y.; Su, D.; Deng, M.; Xiao, N.; Zhang, Z.; Zhou, D.; Gao, B.; Yang, H. Exposure to aerosolized staphylococcal enterotoxin B potentiated by lipopolysaccharide modifies lung transcriptomes and results in lung injury in the mouse model. J. Appl. Toxicol. 2022, 11, 4289. [Google Scholar] [CrossRef] [PubMed]
- Sharma, L.; Riva, A. Intestinal Barrier Function in Health and Disease-Any role of SARS-CoV-2? Microorganisms 2020, 8, 1744. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.S.; Wang, J.; Yannie, P.J.; Ghosh, S. Intestinal Barrier Dysfunction, LPS Translocation, and Disease Development. J. Endocr. Soc. 2020, 4, bvz039. [Google Scholar] [CrossRef] [PubMed]
- Devaux, C.A.; Lagier, J.C.; Raoult, D. New Insights Into the Physiopathology of COVID-19: SARS-CoV-2-Associated Gastrointestinal Illness. Front. Med. 2021, 8, 640073. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhou, Y.; Jiang, N.; Zhou, Q.; Ma, W.L. Persistence of intestinal SARS-CoV-2 infection in patients with COVID-19 leads to re-admission after pneumonia resolved. Int. J. Infect. Dis. 2020, 95, 433–435. [Google Scholar] [CrossRef]
- O’Donnell, J.S.; Chappell, K.J. Chronic SARS-CoV-2, a Cause of Post-acute COVID-19 Sequelae (Long-COVID)? Front. Microbiol. 2021, 12, 64. [Google Scholar] [CrossRef]
- Robinot, R.; Hubert, M.; de Melo, G.D.; Lazarini, F.; Bruel, T.; Smith, N.; Levallois, S.; Larrous, F.; Fernandes, J.; Gellenoncourt, S.; et al. SARS-CoV-2 infection induces the dedifferentiation of multiciliated cells and impairs mucociliary clearance. Nat. Commun. 2021, 12, 4354. [Google Scholar] [CrossRef]
- Teixeira, P.C.; Dorneles, G.P.; Santana Filho, P.C.; da Silva, I.M.; Schipper, L.L.; Postiga, I.A.L.; Moreira Neves, C.A.; Rodrigues Junior, L.C.; Peres, A.; Trindade de Souto, J.; et al. Increased LPS levels coexist with systemic inflammation and result in monocyte activation in severe COVID-19 patients. Int. Immunopharmacol. 2021, 100, 108125. [Google Scholar] [CrossRef]
- Krakauer, T.; Pradhan, K.; Stiles, B.G. Staphylococcal Superantigens Spark Host-Mediated Danger Signals. Front. Immunol. 2016, 7, 23. [Google Scholar] [CrossRef]
- Gold, J.E.; Okyay, R.A.; Licht, W.E.; Hurley, D.J. Investigation of Long COVID Prevalence and Its Relationship to Epstein-Barr Virus Reactivation. Pathogens 2021, 10, 763. [Google Scholar] [CrossRef]
- Moss, P. “The ancient and the new”: Is there an interaction between cytomegalovirus and SARS-CoV-2 infection? Immun. Ageing 2020, 17, 14. [Google Scholar] [CrossRef] [PubMed]
- Franceschini, E.; Cozzi-Lepri, A.; Santoro, A.; Bacca, E.; Lancellotti, G.; Menozzi, M.; Gennari, W.; Meschiari, M.; Bedini, A.; Orlando, G.; et al. Herpes Simplex Virus Re-Activation in Patients with SARS-CoV-2 Pneumonia: A Prospective, Observational Study. Microorganisms 2021, 9, 1896. [Google Scholar] [CrossRef] [PubMed]
- Sutkowski, N.; Palkama, T.; Ciurli, C.; Sekaly, R.P.; Thorley-Lawson, D.; Huber, B.T. An Epstein-Barr virus-associated superantigen. J. Exp. Med. 1996, 184, 971–980. [Google Scholar] [CrossRef] [PubMed]
- Dobrescu, D.; Ursea, B.; Pope, M.; Ascht, A.S.; Posnett, D.N. Enhanced HIV-1 replication in Vβ12 T cells due to human cytomegalovirus in monocytes: Evidence for a putative herpesvirus superantigen. Cell 1995, 82, 753–763. [Google Scholar] [CrossRef]
- Huber, B.T.; Hsu, P.N.; Sutkowski, N. Virus-encoded superantigens. Microbiol. Rev. 1996, 60, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Greenhalgh, T.; Griffin, S.; Gurdasani, D.; Hamdy, A.; Katzourakis, A.; McKee, M.; Michie, S.; Pagel, C.; Roberts, A.; Yates, K. Covid-19: An urgent call for global “vaccines-plus” action. BMJ 2022, 376, 111. [Google Scholar] [CrossRef]
- LeClaire, R.D.; Bavari, S. Human antibodies to bacterial superantigens and their ability to inhibit T-cell activation and lethality. Antimicrob. Agents Chemother. 2001, 45, 460–463. [Google Scholar] [CrossRef]
- Zambrano, L.D.; Newhams, M.M.; Olson, S.M.; Halasa, N.B.; Price, A.M.; Boom, J.A.; Sahni, L.C.; Kamidani, S.; Tarquinio, K.M.; Maddux, A.B.; et al. Effectiveness of BNT162b2 (Pfizer-BioNTech) mRNA Vaccination Against Multisystem Inflammatory Syndrome in Children Among Persons Aged 12–18 Years—United States, July–December 2021. MMWR Morb. Mortal. Wkly. Rep. 2022, 71, 52–58. [Google Scholar] [CrossRef]
- Levin, E.G.; Lustig, Y.; Cohen, C.; Fluss, R.; Indenbaum, V.; Amit, S.; Doolman, R.; Asraf, K.; Mendelson, E.; Ziv, A.; et al. Waning immune humoral response to BNT162b2 Covid-19 vaccine over 6 months. N. Engl. J. Med. 2021, 385, e84. [Google Scholar] [CrossRef]
- Choe, P.G.; Kang, C.K.; Suh, H.J.; Jung, J.; Song, K.; Bang, J.H.; Kim, E.S.; Kim, H.B.; Park, S.W.; Kim, N.J.; et al. Waning Antibody Responses in Asymptomatic and Symptomatic SARS-CoV-2 Infection. Emerg. Infect. Dis. 2021, 27, 327–329. [Google Scholar] [CrossRef]
- Singh, J.; Pandit, P.; McArthur, A.G.; Banerjee, A.; Mossman, K. Evolutionary trajectory of SARS-CoV-2 and emerging variants. Virol. J. 2021, 18, 166. [Google Scholar] [CrossRef] [PubMed]
- Prevalence of Ongoing Symptoms Following Coronavirus (COVID-19) Infection in the UK: 3 March 2022, Office for National Statistics. Available online: https://www.ons.gov.uk/peoplepopulationandcommunity/healthandsocialcare/conditionsanddiseases/bulletins/prevalenceofongoingsymptomsfollowingcoronaviruscovid19infectionintheuk/3march2022 (accessed on 7 February 2022).
- Falasinnu, T.; Chaichian, Y.; Simard, J.F. Impact of Sex on Systemic Lupus Erythematosus-Related Causes of Premature Mortality in the United States. J. Women’s Health 2017, 26, 1214–1221. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, T.M.; Andersen, N.H.; Torp-Pedersen, C.; Søgaard, P.; Kragholm, K.H. Kawasaki disease, autoimmune disorders, and cancer: A register-based study. Eur. J. Pediatr. 2020, 180, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Greenhalgh, T.; Jimenez, J.L.; Prather, K.A.; Tufekci, Z.; Fisman, D.; Schooley, R. Ten scientific reasons in support of airborne transmission of SARS-CoV-2. Lancet 2021, 397, 1603–1605. [Google Scholar] [CrossRef]
- Desforges, M.; Gurdasani, D.; Hamdy, A.; Leonardi, A.J. Uncertainty around the Long-Term Implications of COVID-19. Pathogens 2021, 10, 1267. [Google Scholar] [CrossRef]
- Leonardi, A.J.; Argyropoulos, C.P.; Hamdy, A.; Proenca, R.B. Understanding the Effects of Age and T-Cell Differentiation on COVID-19 Severity: Implicating a Fas/FasL-mediated Feed-Forward Controller of T-Cell Differentiation. Front. Immunol. 2022, 13, 606. [Google Scholar] [CrossRef]
- Showers, W.M.; Leach, S.M.; Kechris, K.; Strong, M. Longitudinal analysis of SARS-CoV-2 spike and RNA-dependent RNA polymerase protein sequences reveals the emergence and geographic distribution of diverse mutations. Infect. Genet. Evol. 2021, 97, 105153, ISSN 1567-1348. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamdy, A.; Leonardi, A. Superantigens and SARS-CoV-2. Pathogens 2022, 11, 390. https://doi.org/10.3390/pathogens11040390
Hamdy A, Leonardi A. Superantigens and SARS-CoV-2. Pathogens. 2022; 11(4):390. https://doi.org/10.3390/pathogens11040390
Chicago/Turabian StyleHamdy, Adam, and Anthony Leonardi. 2022. "Superantigens and SARS-CoV-2" Pathogens 11, no. 4: 390. https://doi.org/10.3390/pathogens11040390
APA StyleHamdy, A., & Leonardi, A. (2022). Superantigens and SARS-CoV-2. Pathogens, 11(4), 390. https://doi.org/10.3390/pathogens11040390