


Mammalian and Avian Larval Schistosomatids in Bangladesh: Molecular Characterization, Epidemiology, Molluscan Vectors, and Occurrence of Human Cercarial Dermatitis

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
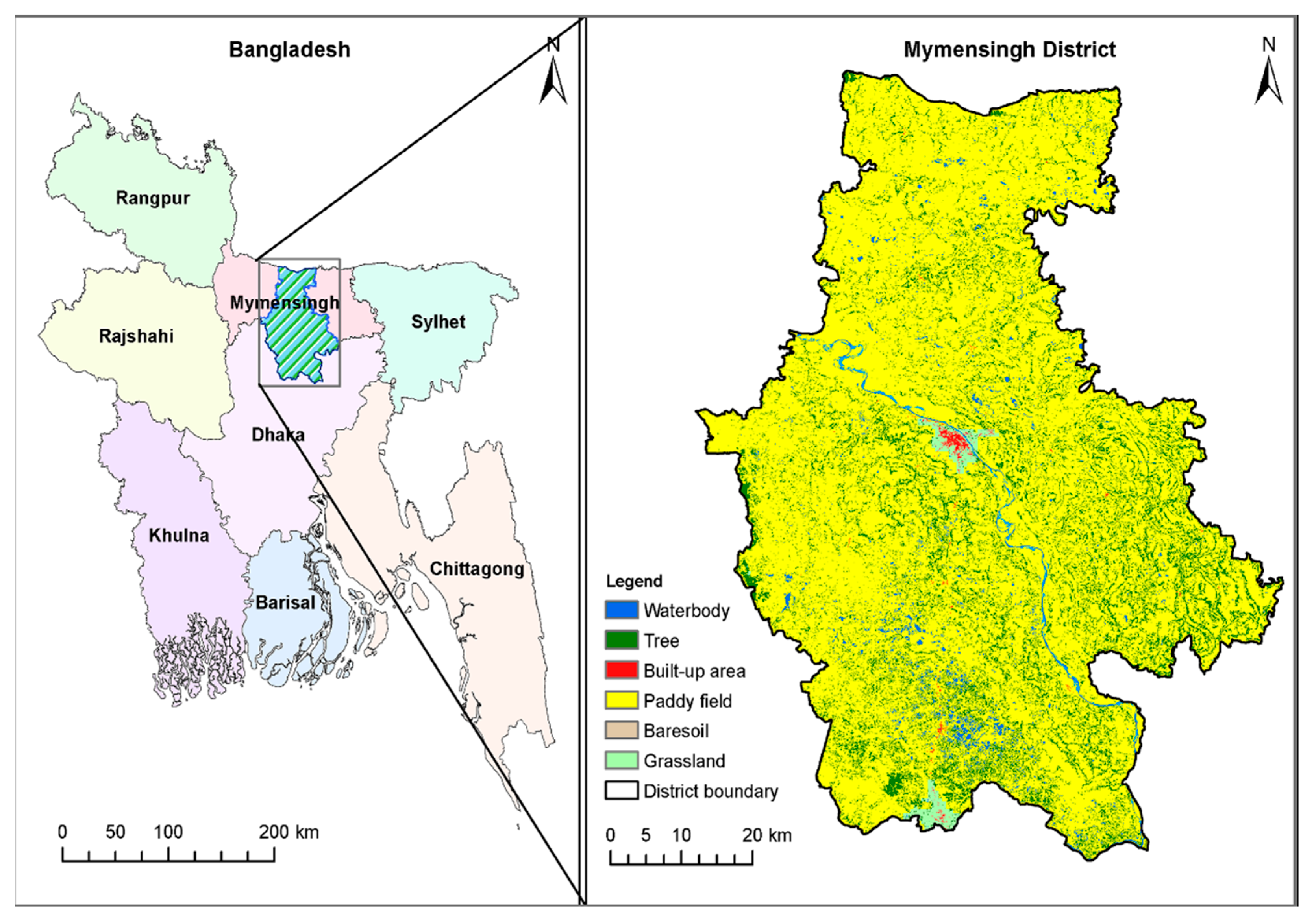
2.2. Study Area, Sample Size, and Collection of Snails
2.3. Shedding and Tentative Identification of SC by Morphological Features
2.4. DNA Extraction
2.5. PCR and Agarose Electrophoresis
2.6. Sequencing
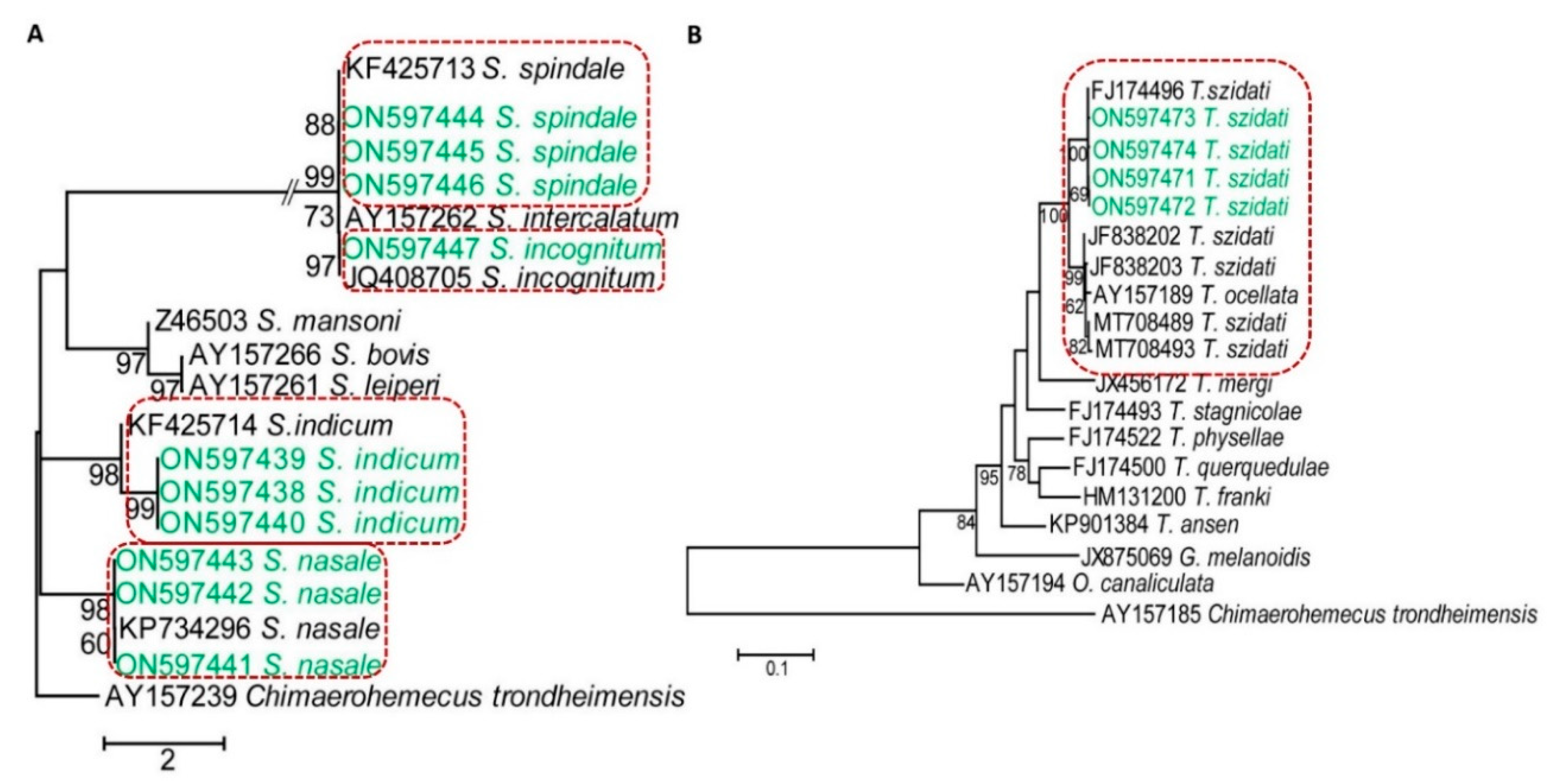
2.7. Genetic Data Analysis and Phylogenetic Study
2.8. Survey of HCD and Case Definition
2.9. Statistical Analysis
3. Results
3.1. Vector Snails of Schistosomatids Prevalent in Bangladesh
3.2. Relative Infection Rates of SC in Different Snail Species in Bangladesh
3.3. Effects of Seasons on the Infection of Snails with Schistosomatids
3.4. Different Habitats of Vector Snails of Schistosomatids
3.5. Confirmation of Species of Schistosomatids by PCR and Sequencing
3.6. Distribution of HCD across the Different Professions in the Study Areas
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anisuzzaman; Tsuji, N. Schistosomiasis and Hookworm Infection in Humans: Disease Burden, Pathobiology and Anthelmintic Vaccines. Parasitol. Int. 2020, 75, 102051. [Google Scholar] [CrossRef] [PubMed]
- Frahm, S.; Anisuzzaman, A.; Prodjinotho, U.F.; Vejzagić, N.; Verschoor, A.; Prazeres da Costa, C. A Novel Cell-Free Method to Culture Schistosoma mansoni from Cercariae to Juvenile Worm Stages for in Vitro Drug Testing. PLoS Negl. Trop. Dis. 2019, 13, e0006590. [Google Scholar] [CrossRef] [PubMed]
- van der Werf, M.J.; de Vlas, S.J.; Brooker, S.; Looman, C.W.; Nagelkerke, N.J.; Habbema, J.D.F.; Engels, D. Quantification of Clinical Morbidity Associated with Schistosome Infection in Sub-Saharan Africa. Acta Trop. 2003, 86, 125–139. [Google Scholar] [CrossRef]
- Hotez, P.J.; Bundy, D.A.P.; Beegle, K.; Brooker, S.; Drake, L.; de Silva, N.; Montresor, A.; Engels, D.; Jukes, M.; Chitsulo, L.; et al. Helminth Infections: Soil-Transmitted Helminth Infections and Schistosomiasis; The International Bank for Reconstruction and Development/The World Bank: Washington, DC, USA, 2006; ISBN 0821361791. [Google Scholar]
- Mawa, P.A.; Kincaid-Smith, J.; Tukahebwa, E.M.; Webster, J.P.; Wilson, S. Schistosomiasis Morbidity Hotspots: Roles of the Human Host, the Parasite and Their Interface in the Development of Severe Morbidity. Front. Immunol. 2021, 12, 751. [Google Scholar] [CrossRef]
- Gray, D.J.; McManus, D.P.; Li, Y.; Williams, G.M.; Bergquist, R.; Ross, A.G. Schistosomiasis Elimination: Lessons from the Past Guide the Future. Lancet Infect. Dis. 2010, 10, 733–736. [Google Scholar] [CrossRef]
- Colley, D.G.; Bustinduy, A.L.; Secor, W.E.; King, C.H. Human Schistosomiasis. Lancet 2014, 383, 2253–2264. [Google Scholar] [CrossRef]
- Horák, P.; Mikeš, L.; Lichtenbergová, L.; Skála, V.; Soldánová, M.; Brant, S.V. Avian Schistosomes and Outbreaks of Cercarial Dermatitis. Clin. Microbiol. Rev. 2015, 28, 165–190. [Google Scholar] [CrossRef]
- Soulsby, E.J.L. Helminths, Arthropods and Protozoa of Domesticated Animals, 7th ed.; Baillière Tindall: London, UK, 1982; ISBN1 0702008206. ISBN2 9780702008207. [Google Scholar]
- Anisuzzaman; Frahm, S.; Prodjinotho, U.F.; Bhattacharjee, S.; Verschoor, A.; Prazeres da Costa, C. Host-Specific Serum Factors Control the Development and Survival of Schistosoma mansoni. Front. Immunol. 2021, 12, 1384. [Google Scholar] [CrossRef]
- Maeda, H.; Hatta, T.; Tsubokawa, D.; Mikami, F.; Nishimaki, T.; Nakamura, T.; Anisuzzaman; Matsubayashi, M.; Ogawa, M.; da Costa, C.P.; et al. Positive Phototropism Is Accelerated in Biomphalaria glabrata Snails by Infection with Schistosoma mansoni. Parasitol. Int. 2018, 67, 609–611. [Google Scholar] [CrossRef] [PubMed]
- Melo, F.L.; Gomes, A.L.; Barbosa, C.S.; Werkhauser, R.P.; Abath, F.G. Development of Molecular Approaches for the Identification of Transmission Sites of Schistosomiasis. Trans. R. Soc. Trop. Med. Hyg. 2006, 100, 1049–1055. [Google Scholar] [CrossRef]
- Knight, M.; Arican-Goktas, H.D.; Ittiprasert, W.; Odoemelam, E.C.; Miller, A.N.; Bridger, J.M. Schistosomes and Snails: A Molecular Encounter. Front. Genet. 2014, 5, 230. [Google Scholar] [CrossRef]
- Rollinson, D.; Webster, J.P.; Webster, B.; Nyakaana, S.; Jørgensen, A.; Stothard, J.R. Genetic Diversity of Schistosomes and Snails: Implications for Control. Parasitology 2009, 136, 1801–1811. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, M.E.; Hellström, M.; Kariuki, H.C.; Olsen, A.; Thomsen, P.F.; Mejer, H.; Willerslev, E.; Mwanje, M.T.; Madsen, H.; Kristensen, T.K.; et al. Environmental DNA for Improved Detection and Environmental Surveillance of Schistosomiasis. Proc. Natl. Acad. Sci. USA 2019, 116, 8931–8940. [Google Scholar] [CrossRef] [PubMed]
- Hung, Y.W.; Remais, J. Quantitative Detection of Schistosomajaponicum Cercariae in Water by Real-Time PCR. PLoS Negl. Trop. Dis. 2008, 2, e337. [Google Scholar] [CrossRef]
- Dias-Neto, E.; Pontes, L.A.; Rabello, A. Detection by Polymerase Chain Reaction of Schistosoma mansoni DNA in Human Serum and Feces. Am. J. Trop. Med. Hyg. 2002, 66, 157–162. [Google Scholar] [CrossRef]
- Malek, E.A.; Cheng, T.C. Medical and Economic Malacology; Academic Press Inc.: New York, NY, USA, 1974; ISBN 0124661505. [Google Scholar]
- Lakshmanan, B.; Devada, K.; Joseph, S.; Aravindakshan, T.V.; Sabu, L. Copro-PCR Based Detection of Bovine Schistosome Infection in India. J. Helminthol. 2016, 90, 102–107. [Google Scholar] [CrossRef]
- Brant, S.V.; Loker, E.S. Molecular Systematics of the Avian Schistosome Genus Trichobilharzia (Trematoda: Schistosomatidae) in North America. J. Parasitol. 2009, 95, 941–963. [Google Scholar] [CrossRef] [PubMed]
- Anisuzzaman; Islam, M.K.; Alim, M.A.; Miyoshi, T.; Hatta, T.; Yamaji, K.; Matsumoto, Y.; Fujisaki, K.; Tsuji, N. Longistatin, a Plasminogen Activator, Is Key to the Availability of Blood-Meals for Ixodid Ticks. PLoS Pathog. 2011, 7, e1001312. [Google Scholar] [CrossRef] [PubMed]
- Lockyer, A.E.; Olson, P.D.; Østergaard, P.; Rollinson, D.; Johnston, D.A.; Attwood, S.W.; Southgate, V.R.; Horak, P.; Snyder, S.D.; Le, T.H.; et al. The Phylogeny of the Schistosomatidae Based on Three Genes with Emphasis on the Interrelationships of Schistosoma Weinland, 1858. Parasitology 2003, 126, 203–224. [Google Scholar] [CrossRef]
- Schuster, R.K.; Aldhoun, J.A.; O’Donovan, D. Gigantobilharzia melanoidis n.Sp. (Trematoda: Schistosomatidae) from Melanoides tuberculata (Gastropoda: Thiaridae) in the United Arab Emirates. Parasitol. Res. 2014, 113, 959–972. [Google Scholar] [CrossRef] [PubMed]
- Jouet, D.; Kolářová, L.; Patrelle, C.; Ferté, H.; Skírnisson, K. Trichobilharzia anseri n. Sp. (Schistosomatidae: Digenea), a New Visceral Species of Avian Schistosomes Isolated from Greylag Goose (Anser anser L.) in Iceland and France. Infect. Genet. Evol. 2015, 34, 298–306. [Google Scholar] [CrossRef]
- Korsunenko, A.; Chrisanfova, G.; Lopatkin, A.; Beer, S.A.; Voronin, M.; Ryskov, A.P.; Semyenova, S.K. Genetic Differentiation of Cercariae Infrapopulations of the Avian Schistosome Trichobilharzia szidati Based on RAPD Markers and Mitochondrial Cox1 Gene. Parasitol. Res. 2012, 110, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Jothikumar, N.; Mull, B.J.; Brant, S.V.; Loker, E.S.; Collinson, J.; Secor, W.E.; Hill, V.R. Real-Time PCR and Sequencing Assays for Rapid Detection and Identification of Avian Schistosomes in Environmental Samples. Appl. Environ. Microbiol. 2015, 81, 4207–4215. [Google Scholar] [CrossRef]
- Littlewood, D.T.J.; Johnston, D.A. Molecular Phylogenetics of the Four Schistosoma Species Groups Determined with Partial 28S Ribosomal RNA Gene Sequences. Parasitology 1995, 111, 167–175. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Sprinthall, R.C. Basic Statistical Analysis, 9th ed.; Pearson: New York, NY, USA, 2011; ISBN 0205052177. [Google Scholar]
- Singh, V.; Kaul, S.C.; Wadhwa, R.; Pati, P.K. Evaluation and Selection of Candidate Reference Genes for Normalization of Quantitative RT-PCR in Withania Somnifera (L.) Dunal. PLoS ONE 2015, 10, e0118860. [Google Scholar] [CrossRef]
- Zbikowska, E. Is There a Potential Danger of swimmer’s Itch in Poland? Parasitol. Res. 2002, 89, 59–62. [Google Scholar] [CrossRef]
- Islam, Z.; Alam, M.; Akter, S.; Roy, B.; Mondal, M. Distribution Patterns of Vector Snails and Trematode Cercaria in Their Vectors in Some Selected Areas of Mymensingh. J. Environ. Sci. Nat. Resour. 2013, 5, 37–46. [Google Scholar] [CrossRef]
- Gordon, C.; Nukpezah, D.; Tweneboah-Lawson, E.; Ofori, B.D.; Yirenya-Tawiah, D.; Pabi, O.; Ayivor, J.S.; Koranteng, S.; Darko, D.; Mensah, A.M. West Africa–Water Resources Vulnerability Using a Multidimensional Approach: Case Study of Volta Basin; Pielke, R.A.B.T.-C.V., Ed.; Academic Press: Oxford, UK, 2013; pp. 283–309. ISBN 978-0-12-384704-1. [Google Scholar]
- Pinto, H.A.; Brant, S.V.; de Melo, A.L. Physa marmorata (Mollusca: Physidae) as a Natural Intermediate Host of Trichobilharzia (Trematoda: Schistosomatidae), a Potential Causative Agent of Avian Cercarial Dermatitis in Brazil. Acta Trop. 2014, 138, 38–43. [Google Scholar] [CrossRef]
- Anisuzzaman; Alim, M.A.; Rahman, M.H.; Mondal, M.M.H. Helminth Parasites in Indigenous Ducks: Seasonal Dynamics and Effects on Production Performance. J. Bangladesh Agric. Univ. 2005, 3, 283–290. [Google Scholar] [CrossRef]
- Labony, S.S.; Paul, S.; Alim, M.A.; Hossain, M.S.; Inoue, T.; Ritu, S.N.; Alam, M.Z.; Alam, M.; Kawada, H.; Hasan, M.M.; et al. Research Note: Genetic Analysis, Pathology, and Vectors of Echinostomiasis, a Zoonotic Helminth Infection in Chickens in Bangladesh. Poult. Sci. 2022, 101, 101682. [Google Scholar] [CrossRef] [PubMed]
- Hamid, M.A.; Ahmed, S.; Rahman, M.A.; Hossain, K.M. Status of Buffalo Production in Bangladesh Compared to SAARC Countries. Asian J. Anim. Sci. 2016, 10, 313–329. [Google Scholar] [CrossRef]
- Hamid, D.M. Duck Genetic Resources, Their Improvement and Conservation in Bangladesh: A Review. SAARC J. Agric. 2020, 17, 31–42. [Google Scholar] [CrossRef]
- Faltýnková, A.; Niewiadomska, K.; Santos, M.; Valtonen, E. Furcocercous Cercariae (Trematoda) from Freshwater Snails in Central Finland. Acta Parasitol. 2007, 52, 310–317. [Google Scholar] [CrossRef]
- Jones, B.P.; Norman, B.F.; Borrett, H.E.; Attwood, S.W.; Mondal, M.M.H.; Walker, A.J.; Webster, J.P.; Rajapakse, R.P.V.J.; Lawton, S.P. Divergence across Mitochondrial Genomes of Sympatric Members of the Schistosoma indicum Group and Clues into the Evolution of Schistosoma spindale. Sci. Rep. 2020, 10, 2480. [Google Scholar] [CrossRef]
- Latchumikanthan, A.; Pothiappan, P.; Ilayabharathi, D.; Das, S.S.; Kumar, D.; Ilangovan, C. Occurrence of Schistosoma nasale Infection in Bullocks of Puducherry. J. Parasit. Dis. 2014, 38, 238–240. [Google Scholar] [CrossRef][Green Version]
- Bunnag, T.; Thirachandra, S.; Impand, P.; Vorasanta, P.; Imlarp, S. Schistosoma incognitum and Its Zoonotic Potential Role in Phitsanulok and Phichit Provinces, Northern Thailand. Southeast Asian J. Trop. Med. Public Health 1983, 14, 163–170. [Google Scholar]
- Kouřilová, P.; Hogg, K.G.; Kolářová, L.; Mountford, A.P. Cercarial Dermatitis Caused by Bird Schistosomes Comprises Both Immediate and Late Phase Cutaneous Hypersensitivity Reactions. J. Immunol. 2004, 172, 3766–3774. [Google Scholar] [CrossRef]
- Kolářová, L.; Horák, P.; Skírnisson, K.; Marečková, H.; Doenhoff, M. Cercarial Dermatitis, a Neglected Allergic Disease. Clin. Rev. Allergy Immunol. 2013, 45, 63–74. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Labony, S.S.; Hossain, M.S.; Hatta, T.; Dey, A.R.; Mohanta, U.K.; Islam, A.; Shahiduzzaman, M.; Hasan, M.M.; Alim, M.A.; Tsuji, N.; et al. Mammalian and Avian Larval Schistosomatids in Bangladesh: Molecular Characterization, Epidemiology, Molluscan Vectors, and Occurrence of Human Cercarial Dermatitis. Pathogens 2022, 11, 1213. https://doi.org/10.3390/pathogens11101213
Labony SS, Hossain MS, Hatta T, Dey AR, Mohanta UK, Islam A, Shahiduzzaman M, Hasan MM, Alim MA, Tsuji N, et al. Mammalian and Avian Larval Schistosomatids in Bangladesh: Molecular Characterization, Epidemiology, Molluscan Vectors, and Occurrence of Human Cercarial Dermatitis. Pathogens. 2022; 11(10):1213. https://doi.org/10.3390/pathogens11101213
Chicago/Turabian StyleLabony, Sharmin Shahid, Md. Shahadat Hossain, Takeshi Hatta, Anita Rani Dey, Uday Kumar Mohanta, Ausraful Islam, Md. Shahiduzzaman, Muhammad Mehedi Hasan, Md. Abdul Alim, Naotoshi Tsuji, and et al. 2022. "Mammalian and Avian Larval Schistosomatids in Bangladesh: Molecular Characterization, Epidemiology, Molluscan Vectors, and Occurrence of Human Cercarial Dermatitis" Pathogens 11, no. 10: 1213. https://doi.org/10.3390/pathogens11101213
APA StyleLabony, S. S., Hossain, M. S., Hatta, T., Dey, A. R., Mohanta, U. K., Islam, A., Shahiduzzaman, M., Hasan, M. M., Alim, M. A., Tsuji, N., & Anisuzzaman. (2022). Mammalian and Avian Larval Schistosomatids in Bangladesh: Molecular Characterization, Epidemiology, Molluscan Vectors, and Occurrence of Human Cercarial Dermatitis. Pathogens, 11(10), 1213. https://doi.org/10.3390/pathogens11101213