
Limited HIV-1 Subtype C nef 3′PPT Variation in Combination Antiretroviral Therapy Naïve and Experienced People Living with HIV in Botswana

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
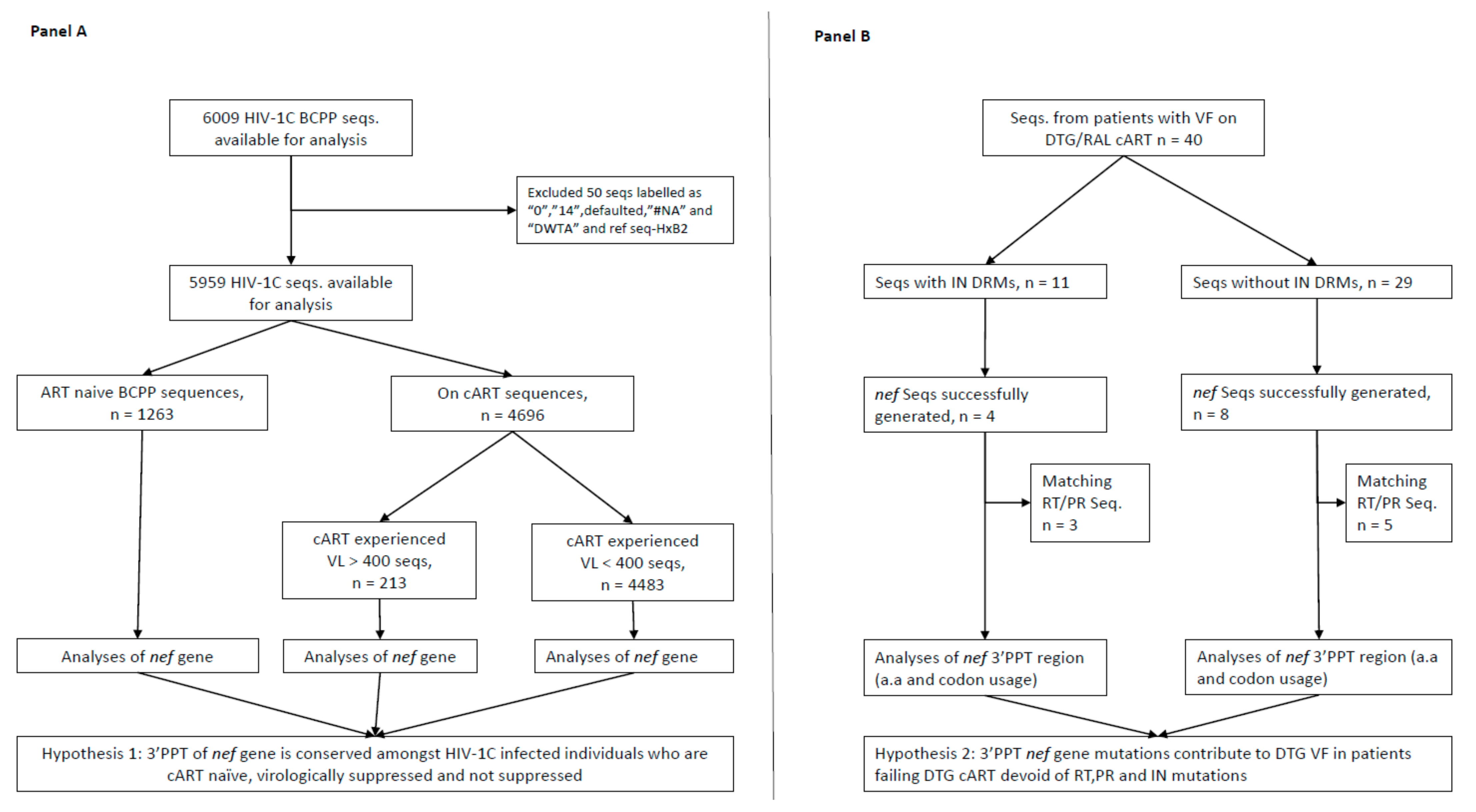
2.1. Selection of Study Population and HIV-1 Genotyping
2.2. Ethical Statement
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anstett, K.; Brenner, B.; Mesplede, T.; Wainberg, M.A. HIV drug resistance against strand transfer integrase inhibitors. Retrovirology 2017, 14, 36. [Google Scholar] [CrossRef]
- DHHS; Panel on Antiretroviral Guidelines for Adults and Adolescents. Guidelines for the Use of Antiretroviral Agents in Adults and Adolescents with HIV; Department of Health and Human Services. 2021. Available online: https://clinicalinfo.hiv.gov/en/guidelines/adult-and-adolescent-arv/whats-new-guidelines (accessed on 13 August 2021).
- MoHW. Handbook of the Botswana 2016 Integrated HIV Clinical Care Guidelines. Available online: https://www.moh.gov.bw/Publications/Handbook_HIV_treatment_guidelines.pdf (accessed on 15 April 2020).
- Raffi, F.; Rachlis, A.; Stellbrink, H.J.; Hardy, W.D.; Torti, C.; Orkin, C.; Bloch, M.; Podzamczer, D.; Pokrovsky, V.; Pulido, F.; et al. Once-daily dolutegravir versus raltegravir in antiretroviral-naive adults with HIV-1 infection: 48 week results from the randomised, double-blind, non-inferiority SPRING-2 study. Lancet 2013, 381, 735–743. [Google Scholar] [CrossRef]
- WHO. Update of Recommendations on First- and Second-Line Antiretroviral Regimens; (WHO/CDS/HIV/19.15); Licence: CC BY-NC-SA 3.0 IGO; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Oliveira, M.; Ibanescu, R.I.; Anstett, K.; Mesplede, T.; Routy, J.P.; Robbins, M.A.; Brenner, B.G.; The Montreal Primary HIV (PHI) Cohort Study Group. Selective resistance profiles emerging in patient-derived clinical isolates with cabotegravir, bictegravir, dolutegravir, and elvitegravir. Retrovirology 2018, 15, 56. [Google Scholar] [CrossRef]
- Seatla, K.K.; Maruapula, D.; Choga, W.T.; Ntsipe, T.; Mathiba, N.; Mogwele, M.; Kapanda, M.; Nkomo, B.; Ramaabya, D.; Makhema, J.; et al. HIV-1 Subtype C Drug Resistance Mutations in Heavily Treated Patients Failing Integrase Strand Transfer Inhibitor-Based Regimens in Botswana. Viruses 2021, 13, 594. [Google Scholar] [CrossRef]
- Malet, I.; Subra, F.; Charpentier, C.; Collin, G.; Descamps, D.; Calvez, V.; Marcelin, A.G.; Delelis, O. Mutations Located outside the Integrase Gene Can Confer Resistance to HIV-1 Integrase Strand Transfer Inhibitors. mBio 2017, 8, e00922-17. [Google Scholar] [CrossRef]
- Wijting, I.E.A.; Lungu, C.; Rijnders, B.J.A.; van der Ende, M.E.; Pham, H.T.; Mesplede, T.; Pas, S.D.; Voermans, J.J.C.; Schuurman, R.; van de Vijver, D.; et al. HIV-1 Resistance Dynamics in Patients With Virologic Failure to Dolutegravir Maintenance Monotherapy. J. Infect. Dis. 2018, 218, 688–697. [Google Scholar] [CrossRef]
- Anderson, S.J.; Lenburg, M.; Landau, N.R.; Garcia, J.V. The cytoplasmic domain of CD4 is sufficient for its down-regulation from the cell surface by human immunodeficiency virus type 1 Nef. J. Virol. 1994, 68, 3092–3101. [Google Scholar] [CrossRef]
- Garcia, J.V.; Miller, A.D. Serine phosphorylation-independent downregulation of cell-surface CD4 by nef. Nature 1991, 350, 508–511. [Google Scholar] [CrossRef]
- Schwartz, O.; Maréchal, V.; Le Gall, S.; Lemonnier, F.; Heard, J.-M. Endocytosis of major histocompatibility complex class I molecules is induced by the HIV–1 Nef protein. Nat. Med. 1996, 2, 338–342. [Google Scholar] [CrossRef]
- Watkins, R.L.; Zou, W.; Denton, P.W.; Krisko, J.F.; Foster, J.L.; Garcia, J.V. In vivo analysis of highly conserved Nef activities in HIV-1 replication and pathogenesis. Retrovirology 2013, 10, 125. [Google Scholar] [CrossRef]
- Das, A.T.; Berkhout, B.; Paraskevis, D. How Polypurine Tract Changes in the HIV-1 RNA Genome Can Cause Resistance against the Integrase Inhibitor Dolutegravir. mBio 2018, 9, e00006-18. [Google Scholar] [CrossRef]
- Julias, J.G.; McWilliams, M.J.; Sarafianos, S.G.; Alvord, W.G.; Arnold, E.; Hughes, S.H. Effects of Mutations in the G Tract of the Human Immunodeficiency Virus Type 1 Polypurine Tract on Virus Replication and RNase H Cleavage. J. Virol. 2004, 78, 13315–13324. [Google Scholar] [CrossRef]
- Jones, F.D.; Hughes, S.H. In vitro analysis of the effects of mutations in the G-tract of the human immunodeficiency virus type 1 polypurine tract on RNase H cleavage specificity. Virology 2007, 360, 341–349. [Google Scholar] [CrossRef][Green Version]
- Malet, I.; Delelis, O.; Nguyen, T.; Leducq, V.; Abdi, B.; Morand-Joubert, L.; Calvez, V.; Marcelin, A.G. Variability of the HIV-1 3′ polypurine tract (3′PPT) region and implication in integrase inhibitor resistance. J. Antimicrob. Chemother. 2019, 74, 3440–3444. [Google Scholar] [CrossRef]
- Wei, Y.; Sluis-Cremer, N. Mutations in the HIV-1 3′-Polypurine Tract and Integrase Strand-Transfer Inhibitor Resistance. Antimicrob. Agents Chemother. 2021, 65, e02432-20. [Google Scholar] [CrossRef]
- Acharya, A.; Tagny, C.T.; Mbanya, D.; Fonsah, J.Y.; Nchindap, E.; Kenmogne, L.; Jihyun, M.; Njamnshi, A.K.; Kanmogne, G.D. Variability in HIV-1 Integrase Gene and 3’-Polypurine Tract Sequences in Cameroon Clinical Isolates, and Implications for Integrase Inhibitors Efficacy. Int. J. Mol. Sci. 2020, 21, 1553. [Google Scholar] [CrossRef]
- Seatla, K.K.; Choga, W.T.; Mogwele, M.; Diphoko, T.; Maruapula, D.; Mupfumi, L.; Musonda, R.M.; Rowley, C.F.; Avalos, A.; Kasvosve, I.; et al. Comparison of an in-house ‘home-brew’ and commercial ViroSeq integrase genotyping assays on HIV-1 subtype C samples. PLoS ONE 2019, 14, e0224292. [Google Scholar] [CrossRef]
- Jones, B.R.; Miller, R.L.; Kinloch, N.N.; Tsai, O.; Rigsby, H.; Sudderuddin, H.; Shahid, A.; Ganase, B.; Brumme, C.J.; Harris, M.; et al. Genetic Diversity, Compartmentalization, and Age of HIV Proviruses Persisting in CD4(+) T Cell Subsets during Long-Term Combination Antiretroviral Therapy. J. Virol. 2020, 94, e01786-19. [Google Scholar] [CrossRef]
- Gaolathe, T.; Wirth, K.E.; Holme, M.P.; Makhema, J.; Moyo, S.; Chakalisa, U.; Yankinda, E.K.; Lei, Q.; Mmalane, M.; Novitsky, V.; et al. Botswana’s progress toward achieving the 2020 UNAIDS 90-90-90 antiretroviral therapy and virological suppression goals: A population-based survey. Lancet HIV 2016, 3, e221–e230. [Google Scholar] [CrossRef]
- Makhema, J.; Wirth, K.E.; Pretorius Holme, M.; Gaolathe, T.; Mmalane, M.; Kadima, E.; Chakalisa, U.; Bennett, K.; Leidner, J.; Manyake, K.; et al. Universal Testing, Expanded Treatment, and Incidence of HIV Infection in Botswana. N. Engl. J. Med. 2019, 381, 230–242. [Google Scholar] [CrossRef]
- Libin, P.J.K.; Deforche, K.; Abecasis, A.B.; Theys, K. VIRULIGN: Fast codon-correct alignment and annotation of viral genomes. Bioinformatics 2018, 35, 1763–1765. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Proc. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]



{kind=link}
{kind=link}
| Basic Characteristics | * HIV-1C Diagnosed Participant Sequences Available for Analysis (n = 6009) | ** Sequences from Participants with VF on DTG cART (n = 40) | |
|---|---|---|---|
| Age (years), median (Q1, Q3) | 40 (33, 48) | 41 (26, 45) | |
| † Gender | Female n (%) | 4241 (71%) | 15 (44%) |
| Male n (%) | 1757 (29%) | 19 (56%) | |
| Unknown n (%) | 11 (0.2%) | ** N/A | |
| Median log 10 HIV-1 RNA (Q1, Q3) copies/mL | 1.60 (1.60, 2.60) | 4.53 (3.98, 5.10) | |
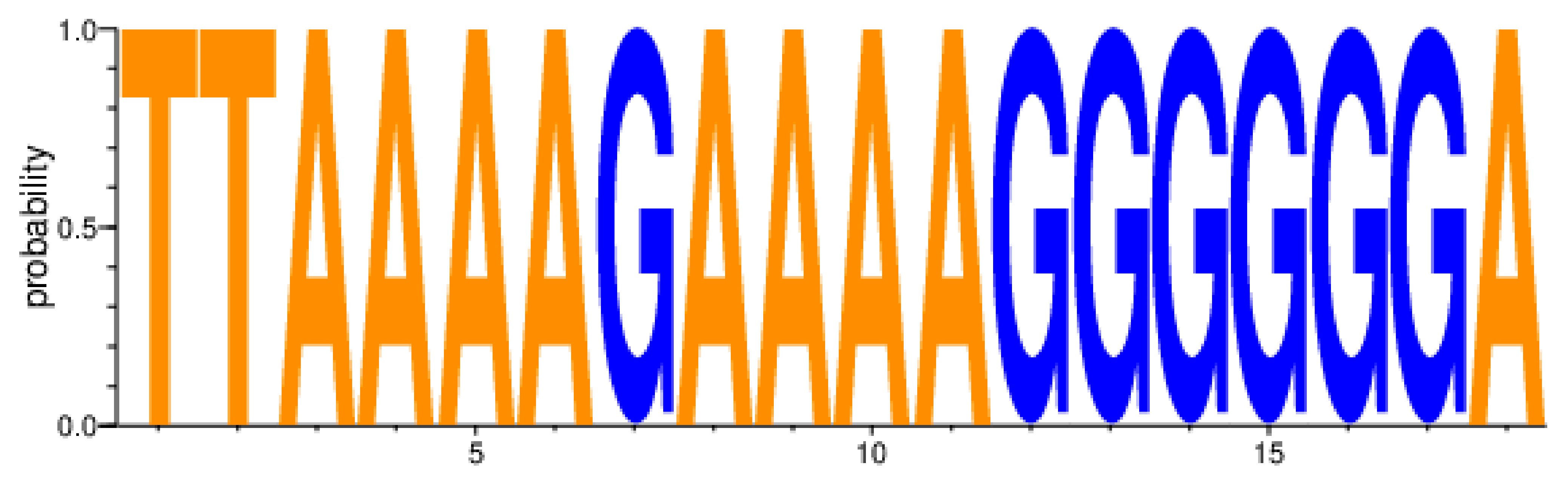
| Β Major DRMs | 3′PPT of the HIV-1 nef Gene | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HxB2_nct_positions | 9067 | 9068 | 9069 | 9070 | 9071 | 9072 | 9073 | 9074 | 9075 | 9076 | 9077 | 9078 | 9079 | 9080 | 9081 | 9082 | 9083 | 9084 | |||
| HxB2_NEF_nct position | 271 | 272 | 273 | 274 | 275 | 276 | 277 | 278 | 279 | 280 | 281 | 282 | 283 | 284 | 285 | 286 | 287 | 288 | |||
| HXB2_NEF_gene_seqs. | RT (NRTI; NNRTI) | PI | INSTI | T | T | A | A | A | A | G | A | A | A | A | G | G | G | G | G | G | A |
| ¥ 139-0001-8 | M41L, D67N, K70KR, V75M, M184V, L210W, T215Y, K219E; A98G, Y181C, G190A | M46I, I47V, I54L, L76V, I84V, Q58E, N83D | E138K, S147G, Q148R, N155H, (E157Q) | T | T | A | A | A | A | G | A | A | A | A | G | G | G | G | G | G | A |
| ¥ 139-0002-8 | ± K70R, M184V;K219N/Y181C (20APRIL2009) ± D67N, K70R, M184V/NONE (18AUG2016) | ± V32I, I47V, I54L, I84V (20 APRIL2009) ± V32I, I47V, I54L, I84V (18AUG2016) | E138K, G140A, S147G, Q148R, (T97A) | T | T | A | A | A | A | G | A | A | A | A | G | G | G | G | G | G | A |
| ¥ 139-0004-6 | ± M41L, T69G, K70R, M184V, T215C, K219E; A98G, K101E | ± M46I, I54V, L76V, V82A | T66A, G118R, E138EAKT | T | T | A | A | A | A | G | A | A | A | A | G | G | G | G | G | G | A |
| 139-0005-3 | * M184V; A98G | * Q58E | * ND | T | T | A | A | A | A | G | A | A | A | A | G | G | G | G | G | G | A |
| ¥ 139-0012-9 | ± M184V, M41L, T215Y; ND (9 June 2010) | ± M46I,V82A Accessory; L10F, L24I (9 June 2010) | N155NH (D232DN) | T | T | A | A | A | A | G | A | A | A | A | G | G | G | G | G | G | A |
| 139-0013-0 | ± M184V, A62V, M41L (28 July 2011); ± E138K (3 February 2016), ± * E138K (24 January 2018) | ± V11IV (28 July 2011) | ND | T | T | A | A | A | A | G | A | A | A | A | G | G | G | G | G | G | A |
| 139-0015-4 | * ND | * ND | * ND | T | T | A | A | A | A | G | A | A | A | A | G | G | G | G | G | G | A |
| 139-0017-2 | * ND | * ND | * ND | T | T | A | A | A | A | G | A | A | A | A | G | G | G | G | G | G | A |
| 139-0018-3 | ± D67G, K70E, M184V; Y181C, G190A | * ND | * ND | T | T | A | A | A | A | G | A | A | A | A | G | G | G | G | G | G | A |
| 139-0021-4 | * K65N; V179D | * ND | * ND | T | T | A | A | A | A | G | A | A | A | A | G | G | G | G | G | G | A |
| 139-0026-8 | * ND | * ND | * ND | T | T | A | A | A | A | G | A | A | A | A | G | G | G | G | G | G | A |
| 139-0119-5 | * ND; K103N, P225H | * ND | * ND | T | T | A | A | A | A | G | A | A | A | A | G | G | G | G | G | G | A |
| Leucine, Leu, L | Lysine, Lys, K | Glutamic acid, Glu, E | Lysine, Lys, K | Glycine, Gly, G | Glycine, Gly, G | ||||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seatla, K.K.; Maruapula, D.; Choga, W.T.; Morerinyane, O.; Lockman, S.; Novitsky, V.; Kasvosve, I.; Moyo, S.; Gaseitsiwe, S. Limited HIV-1 Subtype C nef 3′PPT Variation in Combination Antiretroviral Therapy Naïve and Experienced People Living with HIV in Botswana. Pathogens 2021, 10, 1027. https://doi.org/10.3390/pathogens10081027
Seatla KK, Maruapula D, Choga WT, Morerinyane O, Lockman S, Novitsky V, Kasvosve I, Moyo S, Gaseitsiwe S. Limited HIV-1 Subtype C nef 3′PPT Variation in Combination Antiretroviral Therapy Naïve and Experienced People Living with HIV in Botswana. Pathogens. 2021; 10(8):1027. https://doi.org/10.3390/pathogens10081027
Chicago/Turabian StyleSeatla, Kaelo K., Dorcas Maruapula, Wonderful T. Choga, Olorato Morerinyane, Shahin Lockman, Vladimir Novitsky, Ishmael Kasvosve, Sikhulile Moyo, and Simani Gaseitsiwe. 2021. "Limited HIV-1 Subtype C nef 3′PPT Variation in Combination Antiretroviral Therapy Naïve and Experienced People Living with HIV in Botswana" Pathogens 10, no. 8: 1027. https://doi.org/10.3390/pathogens10081027
APA StyleSeatla, K. K., Maruapula, D., Choga, W. T., Morerinyane, O., Lockman, S., Novitsky, V., Kasvosve, I., Moyo, S., & Gaseitsiwe, S. (2021). Limited HIV-1 Subtype C nef 3′PPT Variation in Combination Antiretroviral Therapy Naïve and Experienced People Living with HIV in Botswana. Pathogens, 10(8), 1027. https://doi.org/10.3390/pathogens10081027