
A Limiting Factor of Sex Attractants of Bactrocera dorsalis (Diptera: Tephritidae), Verified under Laboratory Conditions


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing and Operations
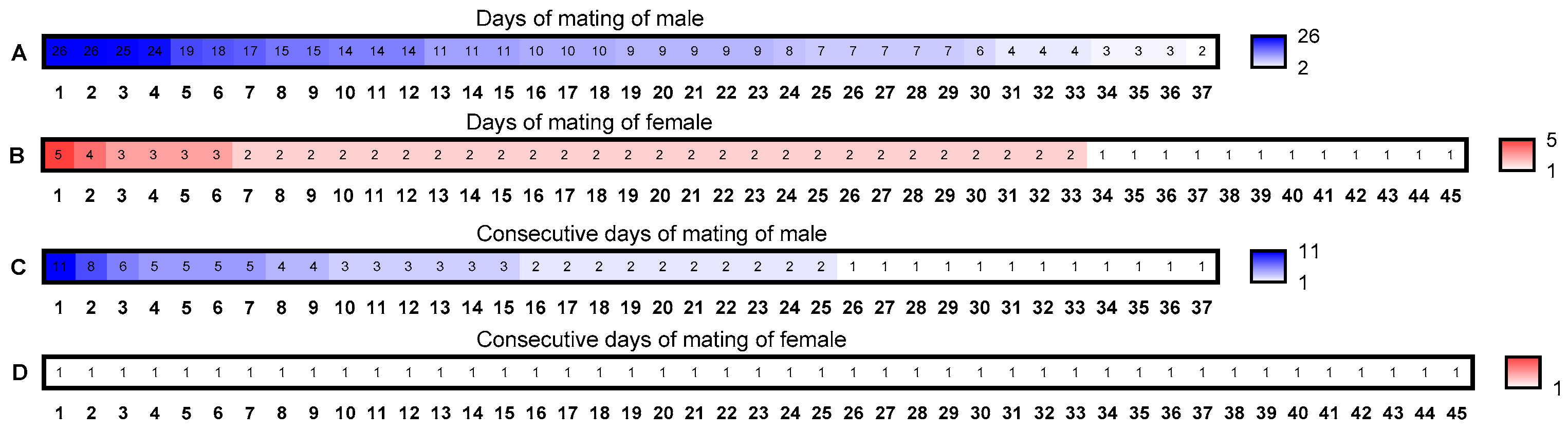
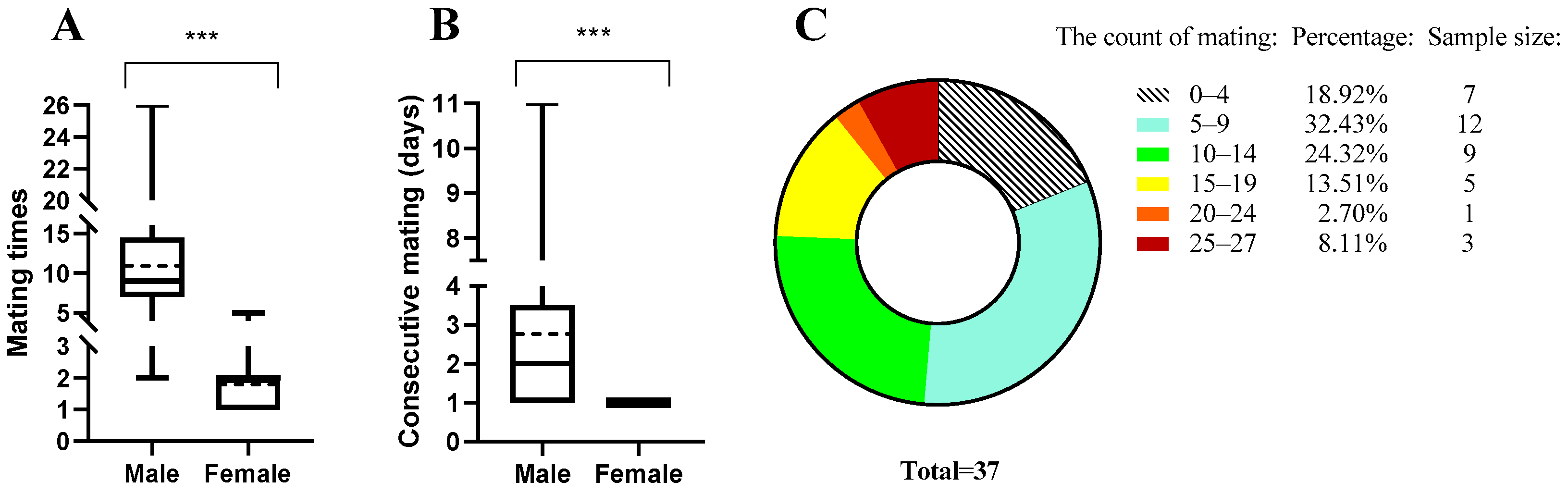
2.2. Mating Frequency Assessment of Male B. dorsalis
2.3. Reproductive Performance of Females Paired with Males with Different Consecutive Mating Modes
2.4. Effect of Mating Opportunity on Control Efficiency
2.5. Statistical Analysis
3. Results
3.1. Mating Ability (Frequency) Assessment of B. dorsalis
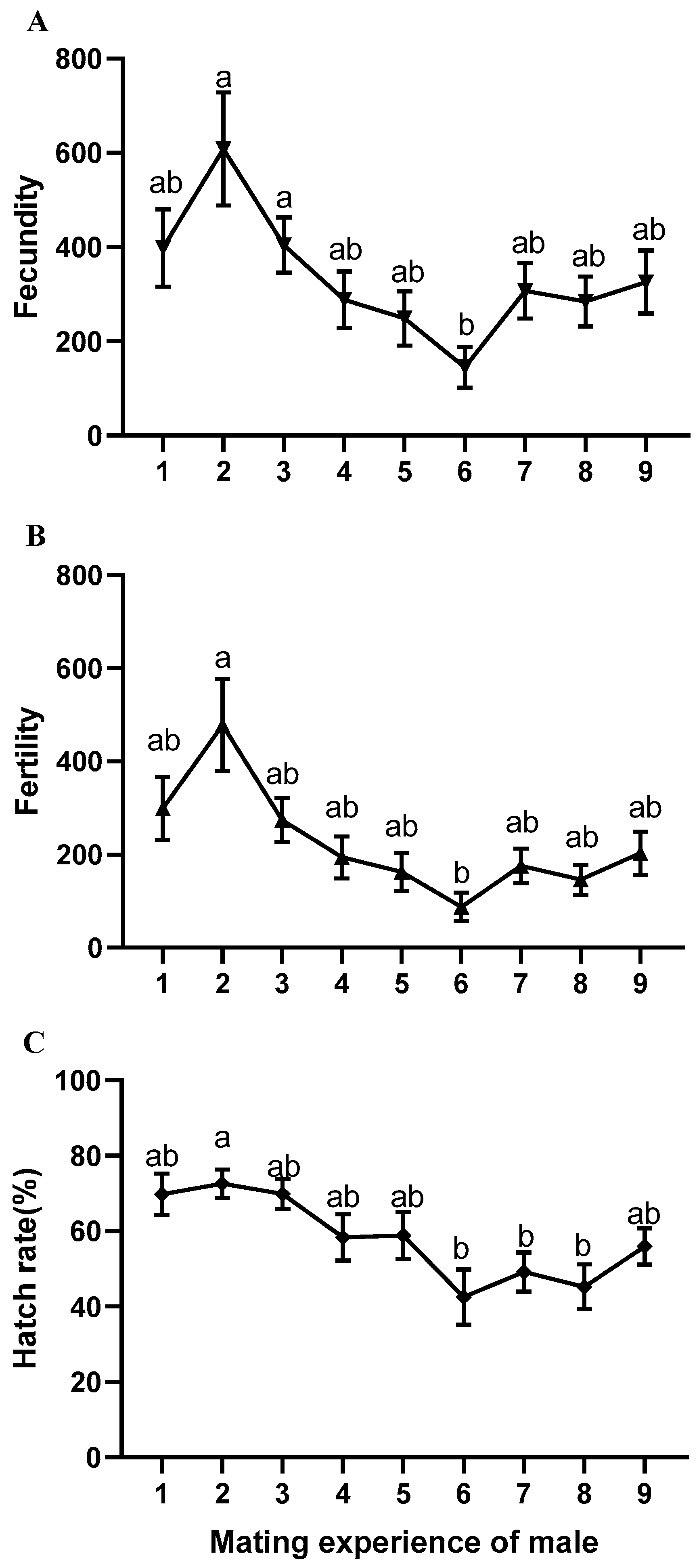
3.2. Reproductive Effect of Male B. dorsalis during Multiple Mating
3.3. Reproductive Fitness Was Influenced by Mating Opportunities with Different Mating Patterns in B. dorsalis
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A
Description
References
- Darwin, C. The Descent of Man, and Selection in Relation to Sex; John Murray: London, UK, 1871. [Google Scholar]
- Wei, D.; Feng, Y.C.; Wei, D.D.; Yuan, G.R.; Dou, W.; Wang, J.J. Female remating inhibition and fitness of Bactrocera dorsalis (Diptera: Tephritidae) associated with male accessory glands. Fla. Entomol. 2015, 98, 52–58. [Google Scholar] [CrossRef]
- Diaz-Munoz, S.L.; Bales, K.L. “Monogamy” in Primates: Variability, Trends, and Synthesis: Introduction to special issue on Primate Monogamy. Am. J. Primatol. 2016, 78, 283–287. [Google Scholar] [CrossRef]
- Lambert, C.T.; Sabol, A.C.; Solomon, N.G. Genetic Monogamy in Socially Monogamous Mammals Is Primarily Predicted by Multiple Life History Factors: A Meta-Analysis. Front. Ecol. Evol. 2018, 6, 139. [Google Scholar] [CrossRef]
- Sutter, A.; Price, T.A.; Wedell, N. The impact of female mating strategies on the success of insect control technologies. Curr. Opin. Insect. Sci. 2021, 45, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Vartak, V.R.; Varma, V.; Sharma, V.K. Effects of polygamy on the activity/rest rhythm of male fruit flies Drosophila melanogaster. Sci. Nat. 2015, 102, 3. [Google Scholar] [CrossRef]
- Hollis, B.; Keller, L.; Kawecki, T.J. Sexual selection shapes development and maturation rates in Drosophila. Evolution 2017, 71, 304–314. [Google Scholar] [CrossRef] [PubMed]
- Clarke, A.R.; Armstrong, K.F.; Carmichael, A.E.; Milne, J.R.; Raghu, S.; Roderick, G.K.; Yeates, D.K. Invasive phytophagous pests arising through a recent tropical evolutionary radiation: The Bactrocera dorsalis complex of fruit flies. Annu. Rev. Entomol. 2005, 50, 293–319. [Google Scholar] [CrossRef]
- Jiang, T.; Zhou, W.; Meng, Y.; Lin, Y.; Shi, Z. Effect of Methyl Eugenol on Male Longevity, Mating and Female Reproduction of Bactrocera dorsalis. Chin. J. Biol. Control 2010, 26, 409–414. [Google Scholar]
- Huang, K.Y.B.; Atlihan, R.; Gökçe, A.; Huang, J.Y.B.; Chi, H. Demographic Analysis of Sex Ratio on Population Growth of Bactrocera dorsalis (Diptera: Tephritidae) with Discussion of Control Efficacy Using Male Annihilation. J. Econ. Entomol. 2016, 109, 2249–2258. [Google Scholar] [CrossRef]
- Guo, Q.; Chen, J.; Yang, C.; Ji, Q.; Huang, J.; Yang, J.; Zheng, M. Relationship between sex ratio and effective eggs amount of genetic sexual strain of Oriental fruit fly. J. Fujian Agric. For. Univ. (Nat. Sci. Ed.) 2010, 39, 571–573. (In Chinese) [Google Scholar] [CrossRef]
- Howlett, F.M. Chemical reactions of fruit-flies. Bull. Entomol. Res. 1915, 6, 297–305. [Google Scholar] [CrossRef]
- Nishida, R.; Tan, K.H.; Serit, M.; Lajis, N.H.; Sukari, A.M.; Takahashi, S.; Fukami, H. Accumulation of phenylpropanoids in the rectal glands of males of the Oriental fruit fly, Dacus dorsalis. Experientia 1988, 44, 534–536. [Google Scholar] [CrossRef]
- Shelly, T.E.; Dewire, A.-L.M. Chemically mediated mating success in male oriental fruit flies (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 1994, 87, 375–382. [Google Scholar] [CrossRef]
- Nishida, R.; Shelly, T.E.; Kaneshiro, K.Y. Acquisition of female-attracting fragrance by males of oriental fruit fly from a Hawaiian Lei flower, Fagraea berteriana. J. Chem. Ecol. 1997, 23, 2275–2285. [Google Scholar] [CrossRef]
- Tan, K.H. Recaptures of feral Bactrocera dorsalis and B. umbrosa (Diptera: Tephritidae) males after feeding on methyl eugenol. Bull. Entomol. Res. 2020, 110, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Manoukis, N.C.; Vargas, R.I.; Carvalho, L.; Fezza, T.; Wilson, S.; Collier, T.; Shelly, T.E. A field test on the effectiveness of male annihilation technique against Bactrocera dorsalis (Diptera: Tephritidae) at varying application densities. PLoS ONE 2019, 14, e0213337. [Google Scholar] [CrossRef] [PubMed]
- Shelly, T.; Kurashima, R.; Fezza, T. Field capture of male oriental fruit flies (Diptera: Tephritidae) in traps baited with solid dispensers containing varying amounts of methyl eugenol. Fla. Entomol. 2020, 103, 516–518. [Google Scholar] [CrossRef]
- Barclay, H.J.; Hendrichs, J. Models for assessing the male annihilation of Bactrocera spp. with methyl eugenol baits. Ann. Entomol. Soc. Am. 2014, 107, 81–96. [Google Scholar] [CrossRef]
- Reyes-Hernandez, M.; Thimmappa, R.; Abraham, S.; Damodaram, K.J.P.; Perez-Staples, D. Methyl eugenol effects on Bactrocera dorsalis male total body protein, reproductive organs and ejaculate. J. Appl. Entomol. 2019, 143, 177–186. [Google Scholar] [CrossRef]
- Wee, S.L.; Munir, M.Z.A.; Hee, A.K.W. Attraction and consumption of methyl eugenol by male Bactrocera umbrosa Fabricius (Diptera: Tephritidae) promotes conspecific sexual communication and mating performance. Bull. Entomol. Res. 2018, 108, 116–124. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, C.; Hou, B.H.; Ou-Yang, G.C.; Ma, J. Interspecific Competition Between Ceratitis capitata and Two Bactrocera spp. (Diptera: Tephritidae) Evaluated via Adult Behavioral Interference Under Laboratory Conditions. J. Econ. Entomol. 2017, 110, 1145–1155. [Google Scholar] [CrossRef]
- Bateman, M.A. The ecology of fruit flies. Annu. Rev. Entomol. 1972, 17, 493–518. [Google Scholar] [CrossRef]
- Roan, C.C.; Flitters, N.E.; Davis, C.J. Light Intensity and Temperature as Factors Limiting the Mating of the Oriental Fruit Fly. Ann. Entomol. Soc. Am. 1954, 47, 593–594. [Google Scholar] [CrossRef]
- Meats, A.; Pike, N.; An, X.; Raphael, K.; Wang, W.Y.S. The effects of selection for early (day) and late (dusk) mating lines of hybrids of Bactrocera tryoni and Bactrocera neohumeralis. Genetica 2003, 119, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Poramarcom, R.; Boake, C.R.B. Behavioural influences on male mating success in the Oriental fruit fly, Dacus dorsalis Hendel. Anim. Behav. 1991, 42, 453–460. [Google Scholar] [CrossRef]
- Londoño-Nieto, C.; García-Roa, R.; Garcia-Co, C.; González, P.; Carazo, P. Thermal phenotypic plasticity of pre- and post-copulatory male harm buffers sexual conflict in wild Drosophila melanogaster. eLife 2023, 12, e84759. [Google Scholar] [CrossRef] [PubMed]
- Holland, B.; Rice, W.R. Experimental removal of sexual selection reverses intersexual antagonistic coevolution and removes a reproductive load. Proc. Natl. Acad. Sci. USA 1999, 96, 5083–5088. [Google Scholar] [CrossRef]
- Iglesias-Carrasco, M.; Fox, R.J.; Vega-Trejo, R.; Jennions, M.D.; Head, M.L. An experimental test for body size-dependent effects of male harassment and an elevated copulation rate on female lifetime fecundity and offspring performance. J. Evol. Biol. 2019, 32, 1262–1273. [Google Scholar] [CrossRef]
- Perez-Staples, D.; Abraham, S. Postcopulatory Behavior of Tephritid Flies. Annu. Rev. Entomol. 2023, 68, 89–108. [Google Scholar] [CrossRef]
- Gliksman, D.; Yuval, B. Intromission Induces and Insemination Reduces Female Immune Response in the Medfly. J. Insect Behav. 2010, 23, 149–158. [Google Scholar] [CrossRef]
- Reyes-Hernandez, M.; Cordova-Garcia, G.; Diaz-Fleischer, F.; Flores-Estevez, N.; Perez-Staples, D. Oviposition after sex: Mated Anastrepha ludens (Diptera: Tephritidae) females increase oviposition without receiving an ejaculate. Can. Entomo. 2021, 153, 524–537. [Google Scholar] [CrossRef]
- Taylor, M.L.; Wigmore, C.; Hodgson, D.J.; Wedell, N.; Hosken, D.J. Multiple mating increases female fitness in Drosophila simulans. Anim. Behav. 2008, 76, 963–970. [Google Scholar] [CrossRef]
- Ji, Q.E.; Chen, J.H.; McInnis, D.O.; Guo, Q.L. The effect of methyl eugenol exposure on subsequent mating performance of sterile males of Bactrocera dorsalis. J. Appl. Entomol. 2013, 137, 238–243. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Consecutive Mating Times of Male | N | Fecundity | Fertility | Oviposition Period (Days) | Longevity (Days) |
|---|---|---|---|---|---|
| 1st | 20 | 398.70 ± 81.79 ab | 299.00 ± 67.28 ab | 22.35 ± 3.10 b | 46.10 ± 3.73 a |
| 2nd | 20 | 650.85 ± 123.11 a | 501.15 ± 101.38 a | 35.55 ± 3.80 a | 59.45 ± 4.29 a |
| 3rd | 15 | 443.67 ± 64.90 ab | 293.80 ± 48.38 ab | 21.93 ± 2.71 b | 43.07 ± 4.04 a |
| 4th | 8 | 223.38 ± 84.71 ab | 144.13 ± 66.32 ab | 14.38 ± 4.33 b | 46.38 ± 8.07 a |
| 5th | 5 | 155.60 ± 66.21 b | 103.60 ± 44.79 b | 16.60 ± 6.24 b | 56.00 ± 9.32 a |
| Mating Patterns | Fecundity | Egg Hatching Rate (%) | Oviposition Period (Days) | Longevity (Days) |
|---|---|---|---|---|
| Polyandry (N = 21) | 552.38 ± 61.88 b | 78.14 ± 2.30 ab | 36.19 ± 3.27 a | 68.81 ± 5.97 a |
| Monogamy (N = 19) | 522.79 ± 77.22 b | 73.85 ± 2.67 bc | 30.89 ± 3.90 a | 53.74 ± 4.45 a |
| Mating again after separation of 2 weeks (N = 27) | 766.80 ± 57.74 a | 81.63 ± 1.74 a | 39.33 ± 2.21 a | 57.44 ± 2.12 a |
| Mating once and then living alone (N = 19) | 505.00 ± 59.85 b | 69.45 ± 3.48 c | 35.32 ± 2.10 a | 57.00 ± 2.70 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Q.; Yi, X.; Wang, X.; Zheng, X.; Lu, W. A Limiting Factor of Sex Attractants of Bactrocera dorsalis (Diptera: Tephritidae), Verified under Laboratory Conditions. Insects 2023, 14, 715. https://doi.org/10.3390/insects14080715
Chen Q, Yi X, Wang X, Zheng X, Lu W. A Limiting Factor of Sex Attractants of Bactrocera dorsalis (Diptera: Tephritidae), Verified under Laboratory Conditions. Insects. 2023; 14(8):715. https://doi.org/10.3390/insects14080715
Chicago/Turabian StyleChen, Qi, Xiaolong Yi, Xiaoyun Wang, Xialin Zheng, and Wen Lu. 2023. "A Limiting Factor of Sex Attractants of Bactrocera dorsalis (Diptera: Tephritidae), Verified under Laboratory Conditions" Insects 14, no. 8: 715. https://doi.org/10.3390/insects14080715
APA StyleChen, Q., Yi, X., Wang, X., Zheng, X., & Lu, W. (2023). A Limiting Factor of Sex Attractants of Bactrocera dorsalis (Diptera: Tephritidae), Verified under Laboratory Conditions. Insects, 14(8), 715. https://doi.org/10.3390/insects14080715