
Mild Winter Causes Increased Mortality in the Fall Webworm Hyphantria cunea (Lepidoptera: Arctiidae)

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Effects of Climatic Conditions on the Abundance of Overwintered Adult Individuals
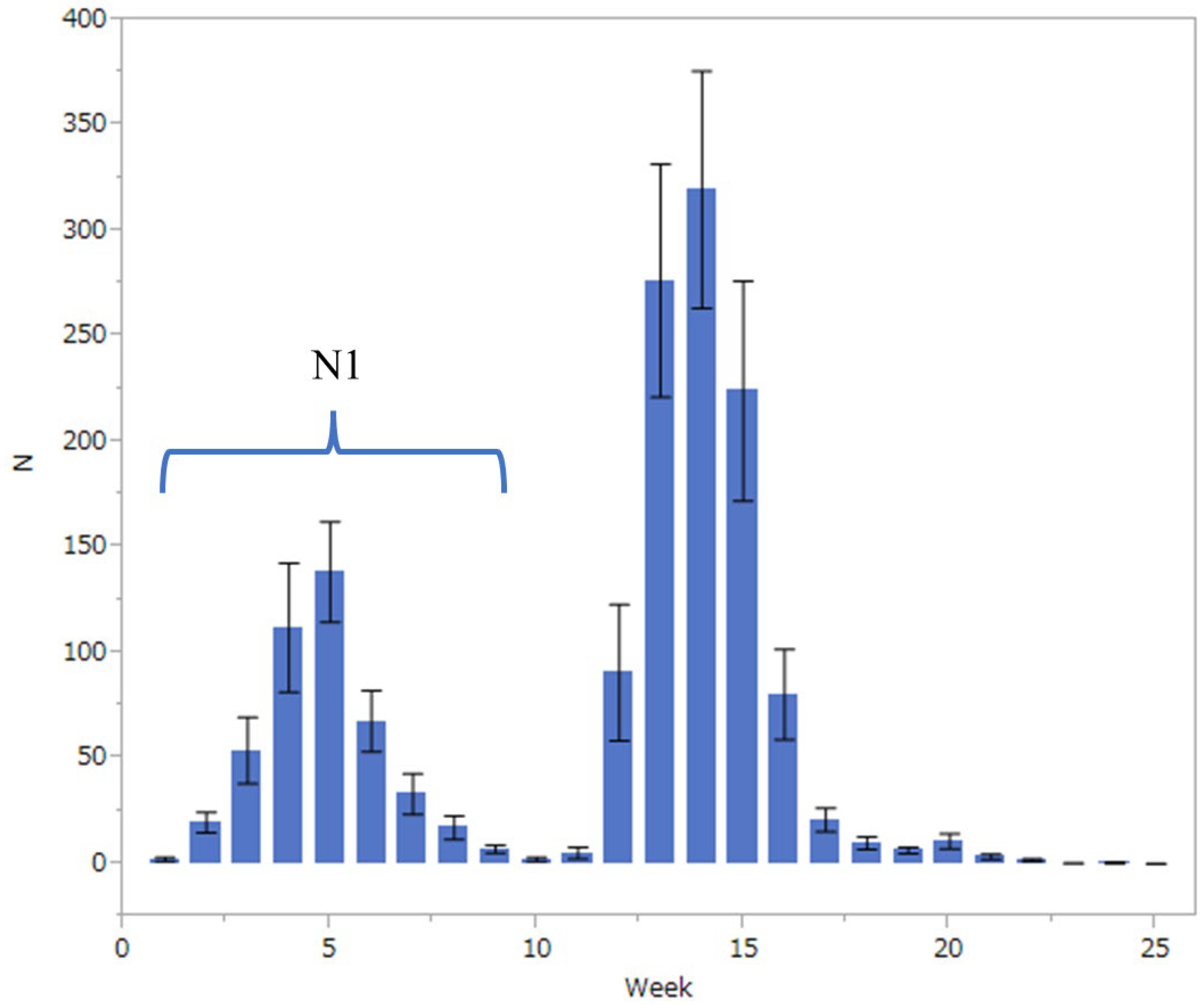
Insect Monitoring
2.2. Climatic Factors
2.3. Effects of Cold Temperatures on Pupal Mortality
2.3.1. Insects and Rearing
2.3.2. Survival, Fungal Infection, and Weight Loss of Pupae in Cold
2.4. Fungal Identification
2.5. Data Analysis
3. Results
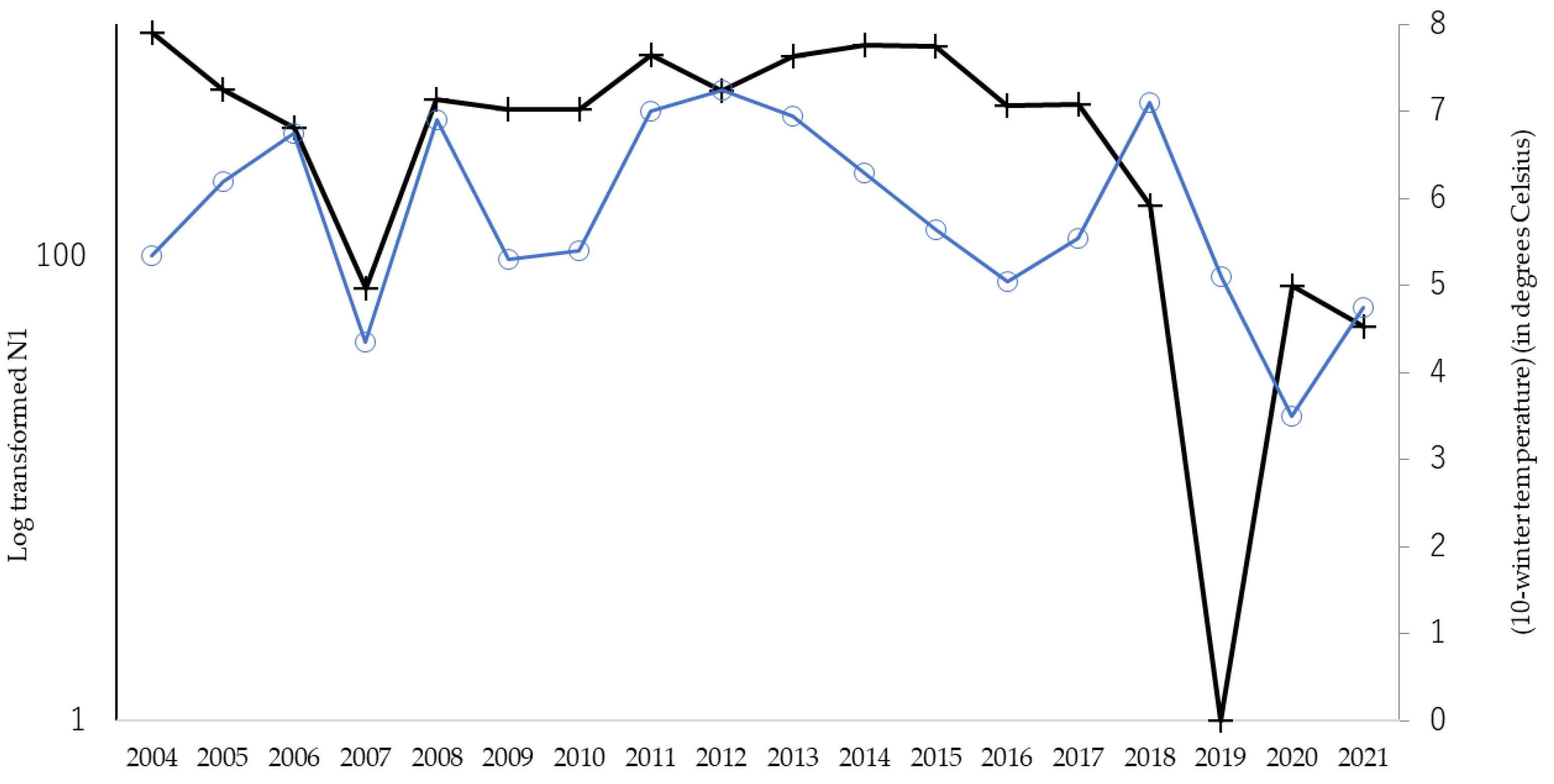
3.1. Relationship between Fall Webworm Outbreaks and Meteorological Conditions
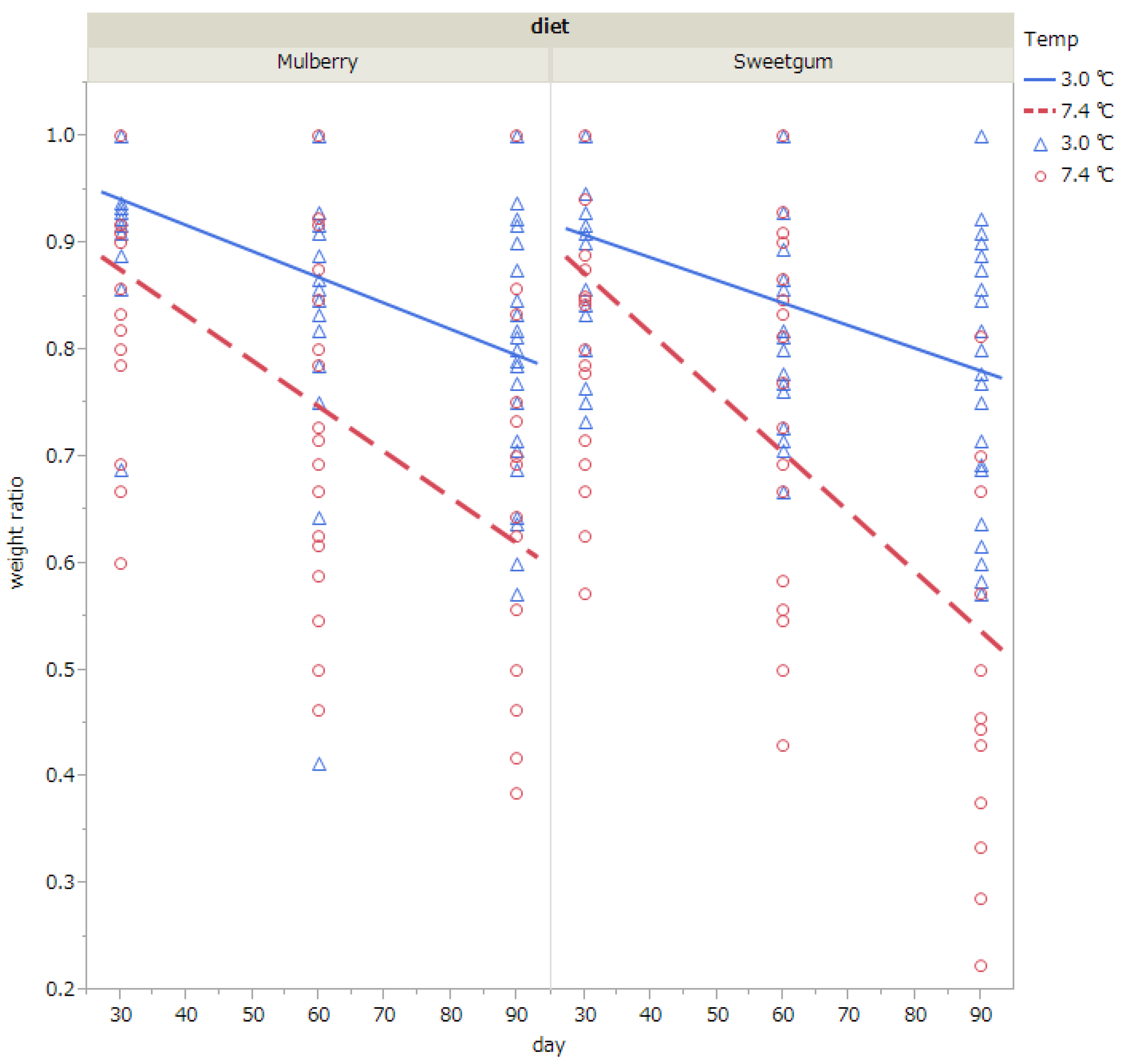
3.2. Survival, Weight Loss, and Fungal Infection of Pupae in Cold
3.3. Identification of Fungi
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Altermatt, F. Climatic warming increases voltinism in European butterflies and moths. Proc. R. Soc. B Biol. Sci. 2010, 277, 1281–1287. [Google Scholar] [CrossRef] [PubMed]
- Mogi, M.; Tuno, N. Impact of climate change on the distribution of Aedes albopictus (Diptera: Culicidae) in northern Japan: Retrospective analyses and future projection. J. Med. Entomol. 2014, 51, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Hill, G.M.; Kawahara, A.Y.; Daniels, J.C.; Bateman, C.C.; Scheffers, B.R. Climate change effects on animal ecology: Butterflies and moths as a case study. Biol. Rev. 2021, 96, 2113–2126. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.H.; Postema, E.G.; Hayes, E.T.; Lippey, M.K.; MacArthur-Waltz, D.J. The complexity of global change and its effects on insects. Curr. Opin. Insect Sci. 2021, 47, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Sendi, J.J.; Johns, R.C.; Takeda, M. Haplotype diversity of mtCOI in the fall webworm Hyphantria cunea (Lepidoptera: Arctiidae) in introduced regions in China, Iran, Japan, Korea, and its homeland, the United States. Appl. Entomol. Zool. 2017, 52, 401–406. [Google Scholar] [CrossRef]
- Masaki, S. Biology of Hyphantria cunea Drury (Lepidoptera: Arctiidae) in Japan: A review. Rev. Plant Prot. Res. 1975, 8, 14–28. [Google Scholar]
- Gomi, T.; Takeda, M. Geographic variation in photoperiodic responses in an introduced insect, Hyphantria cunea Drury (Lepidoptera: Arctiidae) in Japan. Appl. Entomol. Zool. 1991, 25, 483–489. [Google Scholar] [CrossRef]
- Aoki, J. A natural enemy of the fall webworm moth in the mulberry garden, especially the aspect of filamentous fungi. Nihon Sansigaku Zassi 1966, 35, 141–144. [Google Scholar]
- Takezawa, H.; Asami, H.; Mizusawa, Y. Parasitic natural enemies of fall webworms overwintering pupae. Kanto Higashiyama Pest Study Group Annu. Rep. 1969, 16, 134–136. [Google Scholar]
- Kakishima, M.; Tokumasu, S. Biology of Fungi, Classification, Phylogeny, Ecology, Environment, Application; Kyoritsu Shuppan Co., Ltd.: Tokyo, Japan, 2014; 544p. [Google Scholar]
- Itô, Y.; Endo, T. Biology of Hyphantria cunea Drury (Lepidoptera: Arctiidae) in Japan VII. Prediction of adult emergence. Jap. J. Ecol. 1970, 20, 59–62. [Google Scholar]
- Umeya, K.; Masaki, S. Biology of Hyphantria cunea (Drury) (Lepidoptera: Arctiidae) in Japan VII. Delayed development of summer pupae. Res. Bull. Plant Prot. Serv. Jpn. 1969, 7, 1–16. [Google Scholar]
- Itô, Y.; Miyashita, K.; Yamada, H. Biology of Hyphantria Cunea Drury (Lepidoptera: Arctiidae) in Japan: VI. Effect of Temperature on Development of Immature Stages. Appl. Entomol. Zool. 1968, 3, 163–175. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Pörtner, H.O.; Roberts, D.C.; Adams, H.; Adler, C.; Aldunce, P.; Ali, E.; Begum, R.A.; Betts, R.; Kerr, R.B.; Biesbroek, R.; et al. Climate Change 2022: Impacts, Adaptation, and Vulnerability; Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change 2022; Cambridge University Press: Cambridge, UK, 2022. [Google Scholar]
- Aoki, J. Study on the pathogenic mechanism of silk moth Aspergillus (1) Development and pathogenic state of silk moth. Nihon Sansigaku Zassi 1960, 29, 425–430. [Google Scholar]
- Gomi, T.; Adachi, K.; Shimizu, A.; Tanimoto, K.; Kawabata, E.; Takeda, M. Northerly shift in voltinism watershed in Hyphantria cunea (Drury) (Lepidoptera: Arctiidae) along the Japan Sea coast: Evidence of global warming? Appl. Entomol. Zool. 2009, 44, 357–362. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameter Coefficients | |||||
|---|---|---|---|---|---|
| Parameter | Definition | Estimate | Std. Error | t-Value | p |
| (Intercept) | 157.91 | 55.05 | 2.87 | 0.005 | |
| Jan–Feb T | Average daily temperature of Jan. to Feb. | −18.82 | 9.21 | 19.58 | 0.043 |
| Jun-highT-last | Cumulative value of daily average temperature above 25 ° C minus 25 ° C in June of previous year | −3.13 | 0.70 | −4.45 | <0.0001 |
| May T | Cumulative value of daily average temperature above 10 ° C minus 10 ° C in May | 0.15 | 0.13 | 1.11 | 0.268 |
| Jun T | Cumulative value of daily average temperature above 10 ° C minus 10 ° C in June | 0.04 | 0.05 | 0.83 | 0.409 |
| NovMarT | Average daily temperature from Nov. to March. | −9.68 | 10.98 | −0.88 | 0.380 |
| Duration of Exposure | 30 | 60 | 90 | Total | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Temp | Diet | N * | D | F ** | N | D | F ** | N | D | F ** | N | D | F ** |
| 7.4 °C | Sweetgum | 26 | 0 | 0 | 28 | 2 | 2 | 30 | 14 | 14 | 84 | 16 | 16 |
| Mulberry | 30 | 2 | 2 | 26 | 3 | 2 | 22 | 6 | 6 | 78 | 11 | 10 | |
| 3.0 °C | Sweetgum | 26 | 0 | 0 | 22 | 0 | 0 | 46 | 4 | 2 | 94 | 4 | 2 |
| Mulberry | 28 | 0 | 0 | 23 | 0 | 0 | 31 | 3 | 2 | 82 | 3 | 2 | |
| χ2 | p | ||
|---|---|---|---|
| 46.47 | <0.0001 | ||
| Exposure temperature (T) | 12.14 | 0.0005 | |
| Duration of exposure (D) | 16.06 | <0.0001 | |
| Food plant (F) | 6.98 | 0.008 | |
| T × D | 0.24 | 0.621 | |
| T × F | 1.03 | 0.311 | |
| D × F | 14.5 | 0.0001 |
| 3.0 °C | 7.4 °C | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sweetgum | Mulberry | Sweetgum | Mulberry | |||||||||
| Duration of Exposure | N | Mean | SD | N | Mean | SD | N | Mean | SD | N | Mean | SD |
| 30 | 25 | 0.913 | 0.085 | 25 | 0.949 | 0.070 | 26 | 0.840 | 0.121 | 25 | 0.882 | 0.124 |
| 60 | 23 | 0.834 | 0.092 | 23 | 0.851 | 0.124 | 26 | 0.766 | 0.169 | 23 | 0.733 | 0.150 |
| 90 | 42 | 0.783 | 0.112 | 28 | 0.802 | 0.108 | 16 | 0.485 | 0.163 | 16 | 0.629 | 0.174 |
| Whole Model: d.f. = 6, F-Value = 31.46, p < 0.0001. | ||
|---|---|---|
| Factors | t-Value | p |
| Exposure temperature (T) | −8.6 | <0.0001 |
| Duration of exposure (D) | −11 | <0.0001 |
| Food plant (F) | −2 | 0.0441 |
| T × F | −0.57 | 0.5723 |
| D × T | −4.3 | <0.0001 |
| F × D | −0.58 | 0.5641 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsuura, T.; Bangay, R.; Tuno, N. Mild Winter Causes Increased Mortality in the Fall Webworm Hyphantria cunea (Lepidoptera: Arctiidae). Insects 2023, 14, 534. https://doi.org/10.3390/insects14060534
Matsuura T, Bangay R, Tuno N. Mild Winter Causes Increased Mortality in the Fall Webworm Hyphantria cunea (Lepidoptera: Arctiidae). Insects. 2023; 14(6):534. https://doi.org/10.3390/insects14060534
Chicago/Turabian StyleMatsuura, Takahiro, Rohit Bangay, and Nobuko Tuno. 2023. "Mild Winter Causes Increased Mortality in the Fall Webworm Hyphantria cunea (Lepidoptera: Arctiidae)" Insects 14, no. 6: 534. https://doi.org/10.3390/insects14060534
APA StyleMatsuura, T., Bangay, R., & Tuno, N. (2023). Mild Winter Causes Increased Mortality in the Fall Webworm Hyphantria cunea (Lepidoptera: Arctiidae). Insects, 14(6), 534. https://doi.org/10.3390/insects14060534