
The Effect of Resistance to Bt Corn on the Reproductive Output of Spodoptera frugiperda (Lepidoptera: Noctuidae)

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Bioassays
2.3. Statistical Analyses
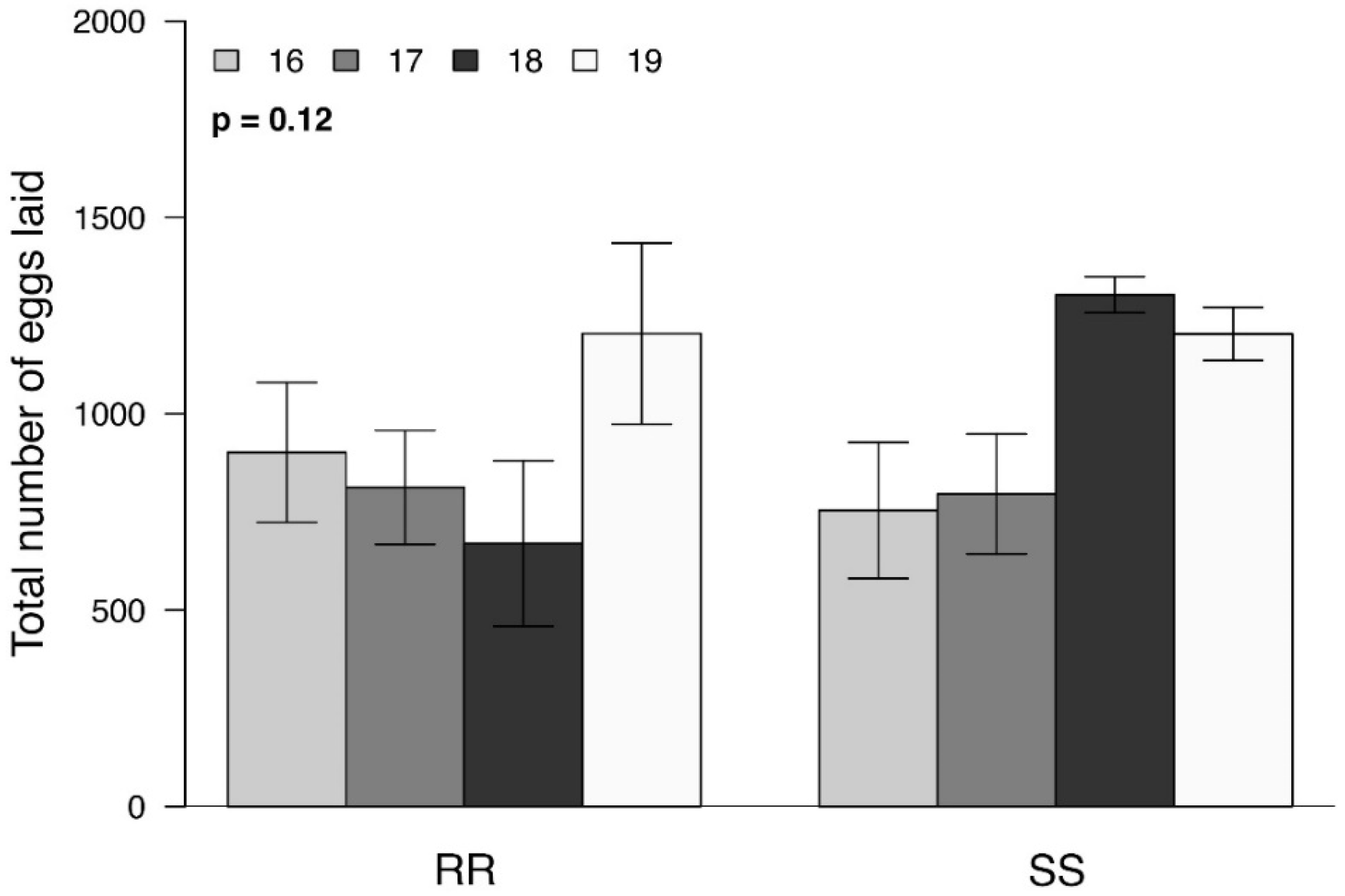
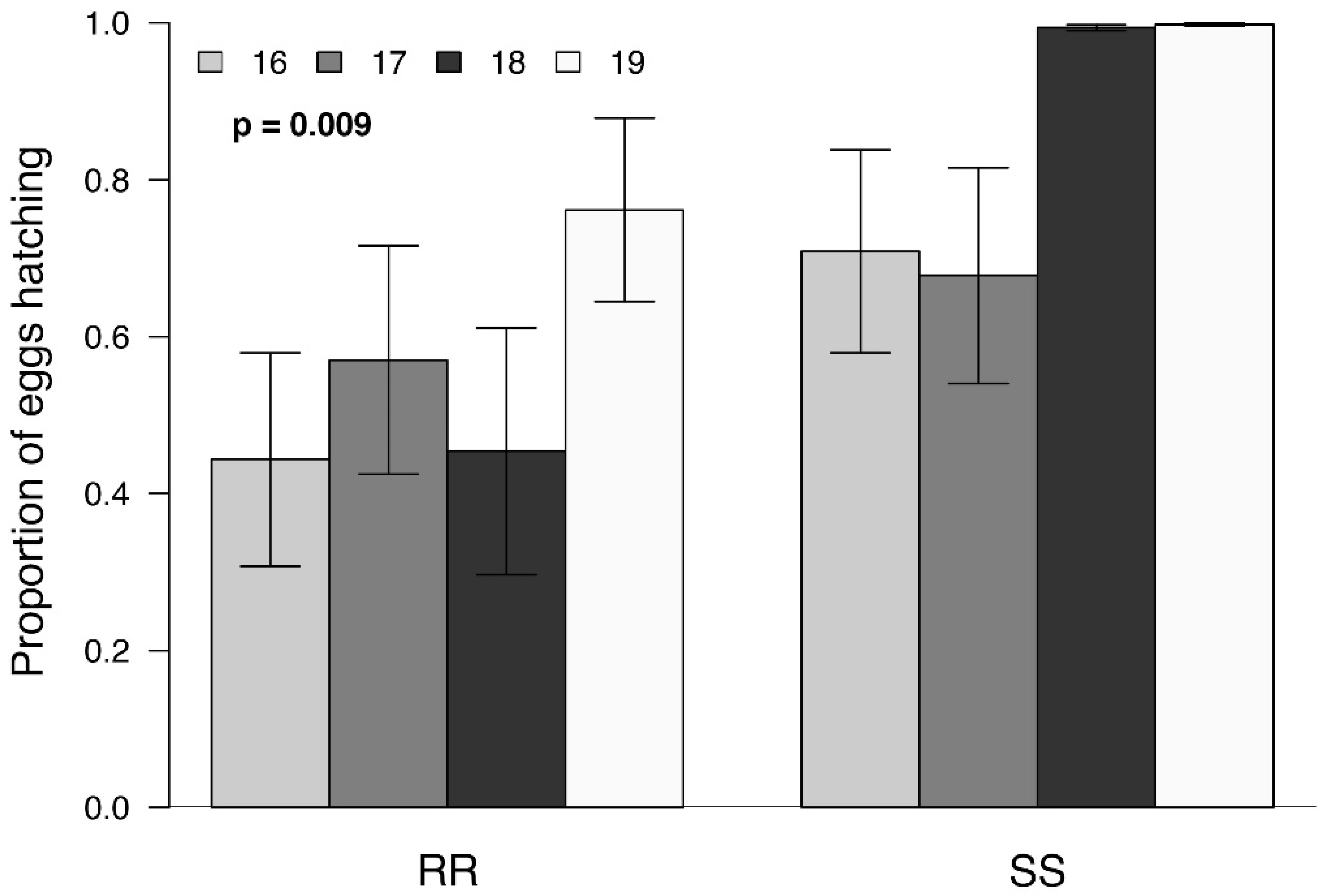
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gould, F. Sustainability of transgenic insecticidal cultivars: Integrating pest genetics and ecology. Annu. Rev. Entomol. 1998, 43, 701–726. [Google Scholar] [CrossRef] [PubMed]
- Okumura, R.S.; Mariano, D.C.; Dallacort, R.; Zorzenoni, T.O.; Zaccheo, P.V.C.; Eugecirc, H.; Lobato, A.K.S. Agronomic efficiency of Bacillus thuringiensis (Bt) maize hybrids in pests control on Lucas do Rio Verde city, State of Mato Grosso, Brazil. Afr. J. Agric. Res. 2013, 8, 2232–2239. [Google Scholar] [CrossRef]
- ISAAA. Global status of commercialized biotech/GM crops in 2017: Biotech crop adoption surges as economic benefits accumulate in 22 years. In ISAAA Brief; ISAAA: Ithaca, NY, USA, 2017; ISBN 978-1-892456-67-2. [Google Scholar]
- Yu, S. Insecticide resistance in the fall armyworm, Spodoptera frugiperda (J.E. Smith). Pestic. Biochem. Physiol. 1991, 39, 84–91. [Google Scholar] [CrossRef]
- Pogue, M.G. A world revision of the genus Spodoptera Guenée: (Lepidoptera: Noctuidae); American Entomological Society: Philadelphia, PA, USA, 2002; Volume 43, pp. 1–202. [Google Scholar]
- Siebert, M.W.; Babock, J.M.; Nolting, S.; Santos, A.C.; Adamczyk, J.J., Jr.; Neese, P.A.; King, J.E.; Jenkins, J.N.; McCarty, J.; Lorenz, G.M.; et al. Efficacy of Cry1F insecticidal protein in maize and cotton for control of fall armyworm (Lepidoptera: Noctuidae). Fla. Entomol. 2008, 91, 555–565. [Google Scholar] [CrossRef][Green Version]
- Blanco, C.A.; Chiaravalle, W.; Dalla-Rizza, M.; Farias, J.R.; García-Degano, M.F.; Gastaminza, G.; Mota-Sánchez, D.; Murúa, M.G.; Omoto, C.; Pieralisi, B.K.; et al. Current situation of pests targeted by Bt crops in Latin America. Curr. Opin. Insect Sci. 2016, 15, 131–138. [Google Scholar] [CrossRef]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (JE Smith)(Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef]
- Fan, J.; Wu, P.; Tian, T.; Ren, Q.; Haseeb, M.; Zhang, R. Potential distribution and niche differentiation of Spodoptera frugiperda in Africa. Insects 2020, 11, 383. [Google Scholar] [CrossRef]
- Downes, S.; Mahon, R.J.; Rossiter, L.; Kauter, G.; Leven, T.; Fitt, G.; Baker, G. Adaptive management of pest resistance by Helicoverpa species (Noctuidae) in Australia to the Cry2Ab Bt toxin in Bollgard II® cotton. Evol. Appl. 2010, 3, 574–584. [Google Scholar] [CrossRef]
- Downes, S.; Walsh, T.; Tay, W.T. Bt resistance in Australian insect pest species. Curr. Opin. Insect Sci. 2016, 15, 78–83. [Google Scholar] [CrossRef]
- Storer, N.P.; Babcock, J.M.; Schlenz, M.; Meade, T.; Thompson, G.D.; Bing, J.W.; Huckaba, R.M. Discovery and characterization of field resistance to Bt maize: Spodoptera frugiperda (Lepidoptera: Noctuidae) in Puerto Rico. J. Econ. Entomol. 2010, 103, 1031–1038. [Google Scholar] [CrossRef]
- Farias, J.R.; Andow, D.A.; Horikoshi, R.J.; Sorgatto, R.J.; Fresia, P.; dos Santos, A.C.; Omoto, C. Field-evolved resistance to Cry1F maize by Spodoptera frugiperda (Lepidoptera: Noctuidae) in Brazil. Crop Prot. 2014, 64, 150–158. [Google Scholar] [CrossRef]
- Huang, F.; Qureshi, J.A.; Meagher, R.L., Jr.; Reisig, D.D.; Head, G.P.; Andow, D.A.; Ni, X.; Kerns, D.; Buntin, G.D.; Niu, Y.; et al. Cry1f resistance in fall armyworm Spodoptera frugiperda: Single gene versus pyramided Bt maize. PLoS ONE 2014, 9, e112958. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, L.A.; Hernández-Juàrez, A.; Flores, M.; Cerna, E.; Landeros, J.; Frías, G.A.; Harris, M.K. Evaluation of foliar damage by Spodoptera frugiperda (Lepidoptera: Noctuidae) to genetically modified corn (Poales: Poaceae) in Mexico. Fla. Entomol. 2016, 99, 276–280. [Google Scholar] [CrossRef]
- Gassmann, A.J.; Carrière, Y.; Tabashnik, B.E. Fitness costs of insect resistance to Bacillus thuringiensis. Annu. Rev. Entomol. 2009, 54, 147–163. [Google Scholar] [CrossRef]
- Fatoretto, J.C.; Michel, A.P.; Silva Filho, M.C.; Silva, N. Adaptive potential of fall armyworm (Lepidoptera: Noctuidae) limits Bt trait durability in Brazil. J. Integr. Pest Manag. 2017, 8, 17. [Google Scholar] [CrossRef]
- Carrière, Y.; Ellers-Kirk, C.; Biggs, R.W.; Nyboer, M.E.; Unnithan, G.C.; Dennehy, T.J.; Tabashnik, B.E. Cadherin-based resistance to Bacillus thuringiensis cotton in hybrid strains of pink bollworm: Fitness costs and incomplete resistance. J. Econ. Entomol. 2006, 99, 1925–1935. [Google Scholar] [CrossRef]
- Zhao, X.C.; Wu, K.M.; Liang, G.M.; Guo, Y.Y. Modified female calling behaviour in Cry1Ac-resistant Helicoverpa armigera (Lepidoptera: Noctuidae). Pest Manag. Sci. Former. Pestic. Sci. 2009, 65, 353–357. [Google Scholar] [CrossRef]
- Zhang, L.; Leonard, B.R.; Chen, M.; Clark, T.; Anilkumar, K.; Huang, F. Fitness costs and stability of Cry1Ab resistance in sugarcane borer, Diatraea saccharalis (F.). J. Invertebr. Pathol. 2014, 117, 26–32. [Google Scholar] [CrossRef]
- Delisle, J.; Bouchard, A. Male larval nutrition in Choristoneura rosaceana (lepidoptera: Tortricidae): An important factor in reproductive success. Oecologia 1995, 104, 508–517. [Google Scholar] [CrossRef]
- Higginson, D.M.; Morin, S.; Nyboer, M.E.; Biggs, R.W.; Tabashnik, B.E.; Carrière, Y. Evolutionary trade-offs of insect resistance to Bacillus thuringiensis crops: Fitness cost affecting paternity. Evolution 2005, 59, 915–920. [Google Scholar] [CrossRef]
- Zhang, W.; Ma, L.; Zhong, F.; Wang, Y.; Guo, Y.; Lu, Y.; Liang, G. Fitness costs of reproductive capacity and ovarian development in a Bt-resistant strain of the cotton bollworm Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). Pest Manag. Sci. 2015, 71, 870–877. [Google Scholar] [CrossRef] [PubMed]
- Royer, L.; McNeil, J. Male investment in the european corn borer, Ostrinia nubilalis (Lepidoptera: Pyralidae): Impact on female longevity and reproductive performance. Funct. Ecol. 1993, 7, 209–215. [Google Scholar] [CrossRef]
- Butler, M., IV; Macdiarmid, A.; Gnanalingam, G. The effect of parental size on spermatophore production, egg quality, fertilization success, and larval characteristics in the Caribbean Spiny lobster, Panulirus argus. ICES J. Mar. Sci. 2015, 72, i115–i123. [Google Scholar] [CrossRef]
- Duplouy, A.; Hanski, I. Small spermatophore size and reduced female fitness in an isolated butterfly population. Ecol. Entomol. 2015, 40, 167–174. [Google Scholar] [CrossRef]
- He, Y.; Miyata, T. Variations in sperm number in relation to larval crowding and spermatophore size in the armyworm, Pseudaletia separata. Ecol. Entomol. 1997, 22, 41–46. [Google Scholar] [CrossRef]
- Svärd, L.; Wiklund, C. Different ejaculate delivery strategies in first versus subsequent matings in the swallowtail butterfly Papilio machaon L. Behav. Ecol. Sociobiol. 1986, 18, 325–330. [Google Scholar] [CrossRef]
- Wedell, N.; Karlsson, B. Paternal investment directly affects female reproductive effort in an insect. Proc. R. Soc. Lond. B Biol. Sci. 2003, 270, 2065–2071. [Google Scholar] [CrossRef] [PubMed]
- Cahenzli, F.; Erhardt, A. Nectar amino acids enhance reproduction in male butterflies. Oecologia 2013, 171, 197–205. [Google Scholar] [CrossRef]
- Santos-Amaya, O.F.; Tavares, C.S.; Monteiro, H.M.; Teixeira, T.P.; Guedes, R.N.; Alves, A.P.; Pereira, E.J. Genetic basis of Cry1F resistance in two Brazilian populations of fall armyworm, Spodoptera frugiperda. Crop Prot. 2016, 81, 154–162. [Google Scholar] [CrossRef]
- Santos-Amaya, O.F.; Rodrigues, J.V.; Souza, T.C.; Tavares, C.S.; Campos, S.O.; Guedes, R.N.; Pereira, E.J. Resistance to dual-gene Bt maize in Spodoptera frugiperda: Selection, inheritance and cross-resistance to other transgenic events. Sci. Rep. 2015, 5, 18243. [Google Scholar] [CrossRef]
- Kasten, P.; Precetti, A.; Parra, J. Dados biológicos comparativos de Spodoptera frugiperda (J.E. Smith, 1797) em duas dietas artificiais e substrato natural. Rev. Agric. 1978, 53, 68–78. [Google Scholar]
- Jakka, S.R.K.; Knight, V.R.; Jurat-Fuentes, J.L. Fitness costs associated with field-evolved resistance to Bt maize in Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Econ. Entomol. 2014, 107, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Horikoshi, R.J.; Bernardi, O.; Bernardi, D.; Okuma, D.M.; Farias, J.R.; Miraldo, L.L.; Amaral, F.S.A.; Omoto, C. Near-isogenic Cry1F-resistant strain of Spodoptera frugiperda (Lepidoptera: Noctuidae) to investigate fitness cost associated with resistance in Brazil. J. Econ. Entomol. 2016, 109, 854–859. [Google Scholar] [CrossRef] [PubMed]
- Dangal, V.; Huang, F. Fitness costs of Cry1F resistance in two populations of fall armyworm, Spodoptera frugiperda (JE Smith), collected from Puerto Rico and Florida. J. Invertebr. Pathol. 2015, 127, 81–86. [Google Scholar] [CrossRef]
- Gao, Y.; Kang, L. Operational sex ratio and alternative reproductive behaviours in Chinese bushcricket, Gampsocleis gratiosa. Ethology 2006, 112, 325–331. [Google Scholar] [CrossRef]
- Duplouy, A.; Woestmann, L.; Gallego Zamorano, J.; Saastamoinen, M. Impact of male condition on his spermatophore and consequences for female reproductive performance in the Glanville fritillary butterfly. Insect Sci. 2018, 25, 284–296. [Google Scholar] [CrossRef] [PubMed]
- Caballero-Mendieta, N.; Cordero, C. Male mating costs in a butterfly that produces small ejaculates. Physiol. Entomol. 2013, 38, 318–325. [Google Scholar] [CrossRef]
- Boggs, C.L.; Gilbert, L.E. Male contribution to egg production in butterflies: Evidence for transfer of nutrients at mating. Science 1979, 206, 83–84. [Google Scholar] [CrossRef] [PubMed]
- Hellriegel, B.; Bernasconi, G. Female-mediated differential sperm storage in a fly with complex spermathecae, Scatophaga stercoraria. Anim. Behav. 2000, 59, 311–317. [Google Scholar] [CrossRef][Green Version]
- Seth, R.K.; Kaur, J.J.; Rao, D.K.; Reynolds, S.E. Sperm transfer during mating, movement of sperm in the female reproductive tract, and sperm precedence in the common cutworm Spodoptera litura. Physiol. Entomol. 2002, 27, 1–14. [Google Scholar] [CrossRef]
- Watanabe, M.; Bon’no, M.; Hachisuka, A. Eupyrene sperm migrates to spermatheca after apyrene sperm in the swallowtail butterfly, Papilio xuthus L. (Lepidoptera: Papilionidae). J. Ethol. 2000, 18, 91–99. [Google Scholar] [CrossRef]
- Sih, A.; Krupa, J.; Travers, S. An experimental study on the effects of predation risk and feeding regime on the mating behavior of the water strider. Am. Nat. 1990, 135, 284–290. [Google Scholar] [CrossRef]
- Yuval, B.; Bouskila, A. Temporal dynamics of mating and predation in mosquito swarms. Oecologia 1993, 95, 65–69. [Google Scholar] [CrossRef]
- Rodrigues, A.R.; Torres, J.B.; Siqueira, H.A.; Teixeira, V.W. Podisus nigrispinus (Dallas) (Hemiptera: Pentatomidae) requires long matings for successful reproduction. Neotrop. Entomol. 2009, 38, 746–753. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Souza Ribas, N.; McNeil, J.N.; Araújo, H.D.; de Souza Ribas, B.; Lima, E. The Effect of Resistance to Bt Corn on the Reproductive Output of Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2022, 13, 196. https://doi.org/10.3390/insects13020196
de Souza Ribas N, McNeil JN, Araújo HD, de Souza Ribas B, Lima E. The Effect of Resistance to Bt Corn on the Reproductive Output of Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects. 2022; 13(2):196. https://doi.org/10.3390/insects13020196
Chicago/Turabian Stylede Souza Ribas, Natália, Jeremy N. McNeil, Hernane Dias Araújo, Bruna de Souza Ribas, and Eraldo Lima. 2022. "The Effect of Resistance to Bt Corn on the Reproductive Output of Spodoptera frugiperda (Lepidoptera: Noctuidae)" Insects 13, no. 2: 196. https://doi.org/10.3390/insects13020196
APA Stylede Souza Ribas, N., McNeil, J. N., Araújo, H. D., de Souza Ribas, B., & Lima, E. (2022). The Effect of Resistance to Bt Corn on the Reproductive Output of Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects, 13(2), 196. https://doi.org/10.3390/insects13020196