
Comparison of the Parasitization of Chelonus inanitus L. (Hymenoptera: Braconidae) in Two Spodoptera Pests and Evaluation of the Procedure for Its Production

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. C. inanitus Development in the Factitious Host E. kuehniella
2.3. Semi-Field Studies on the Efficacy of C. inanitus in S. exigua and S. littoralis
2.4. Statistical Analysis
3. Results
3.1. C. inanitus Development in the Factitious Host E. kuehniella
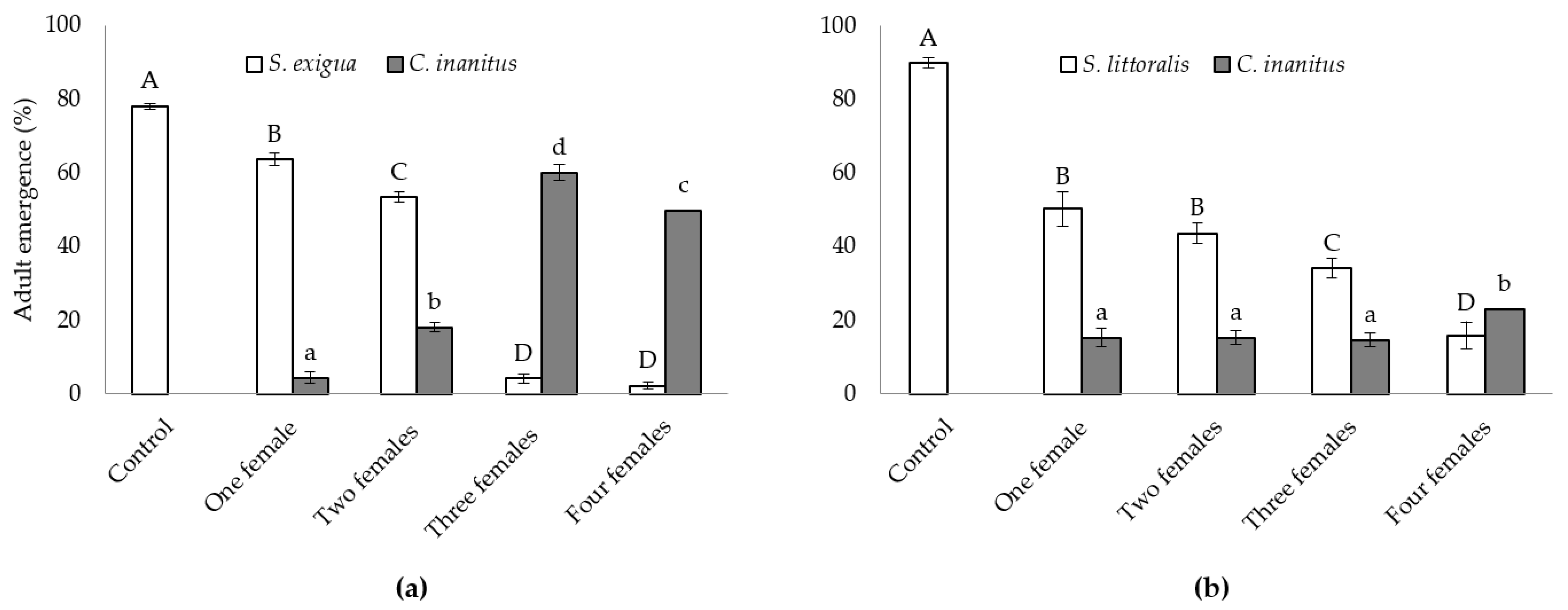
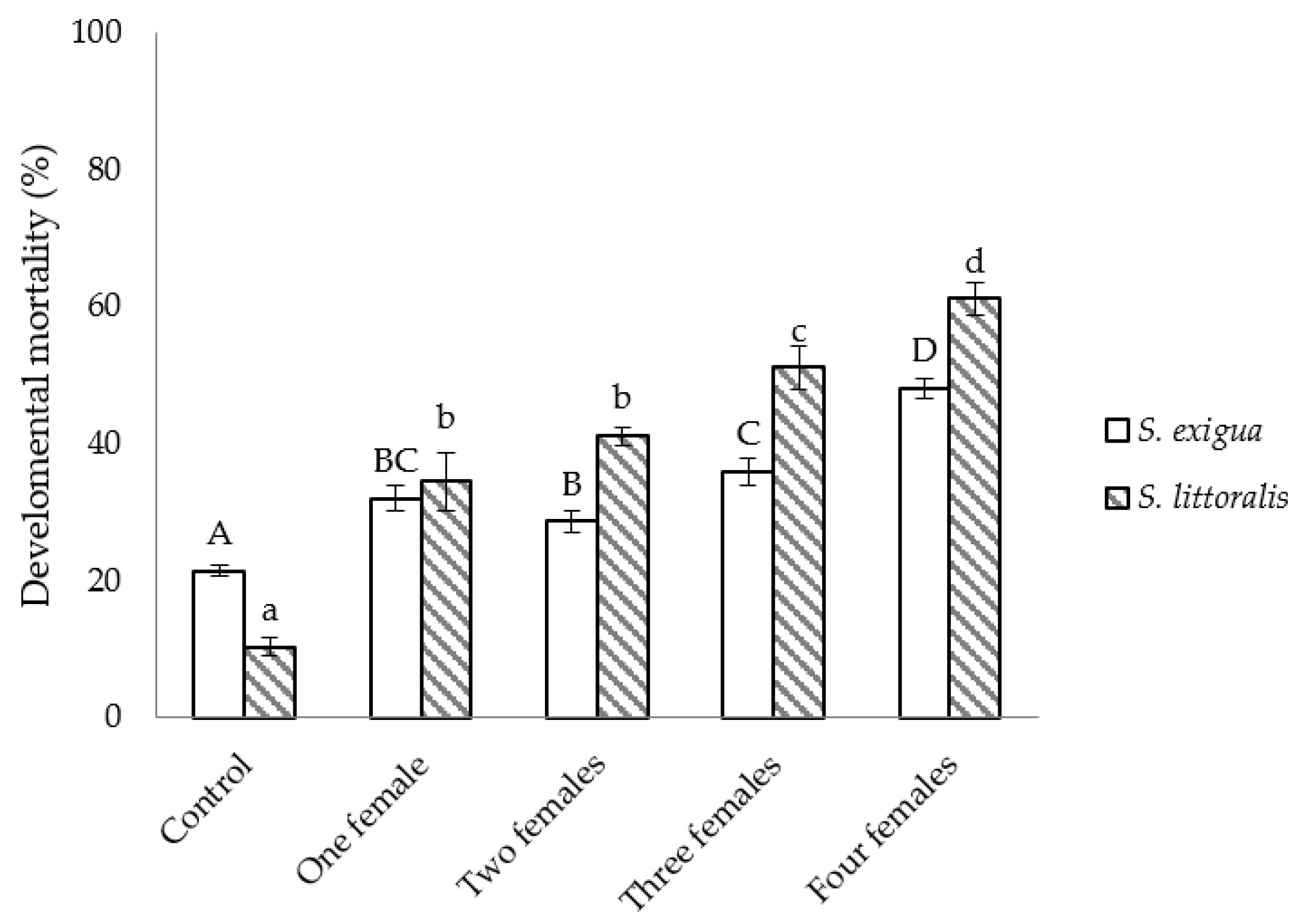
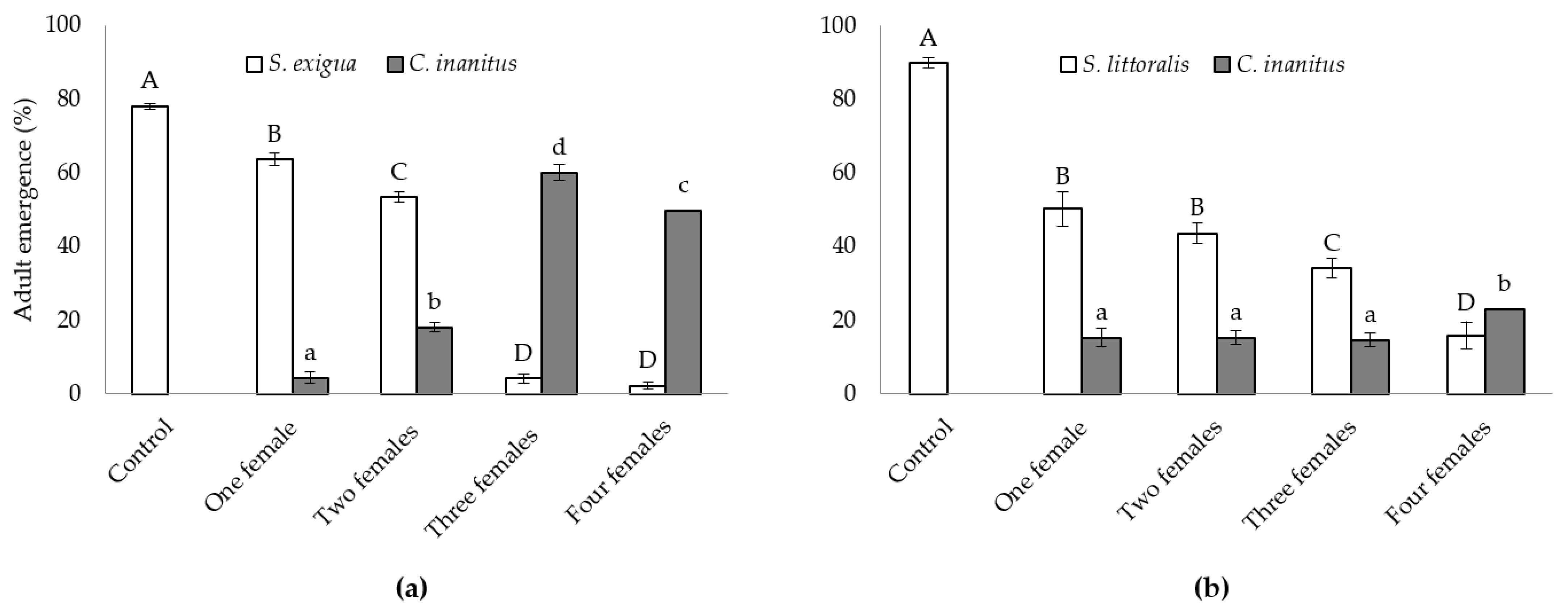
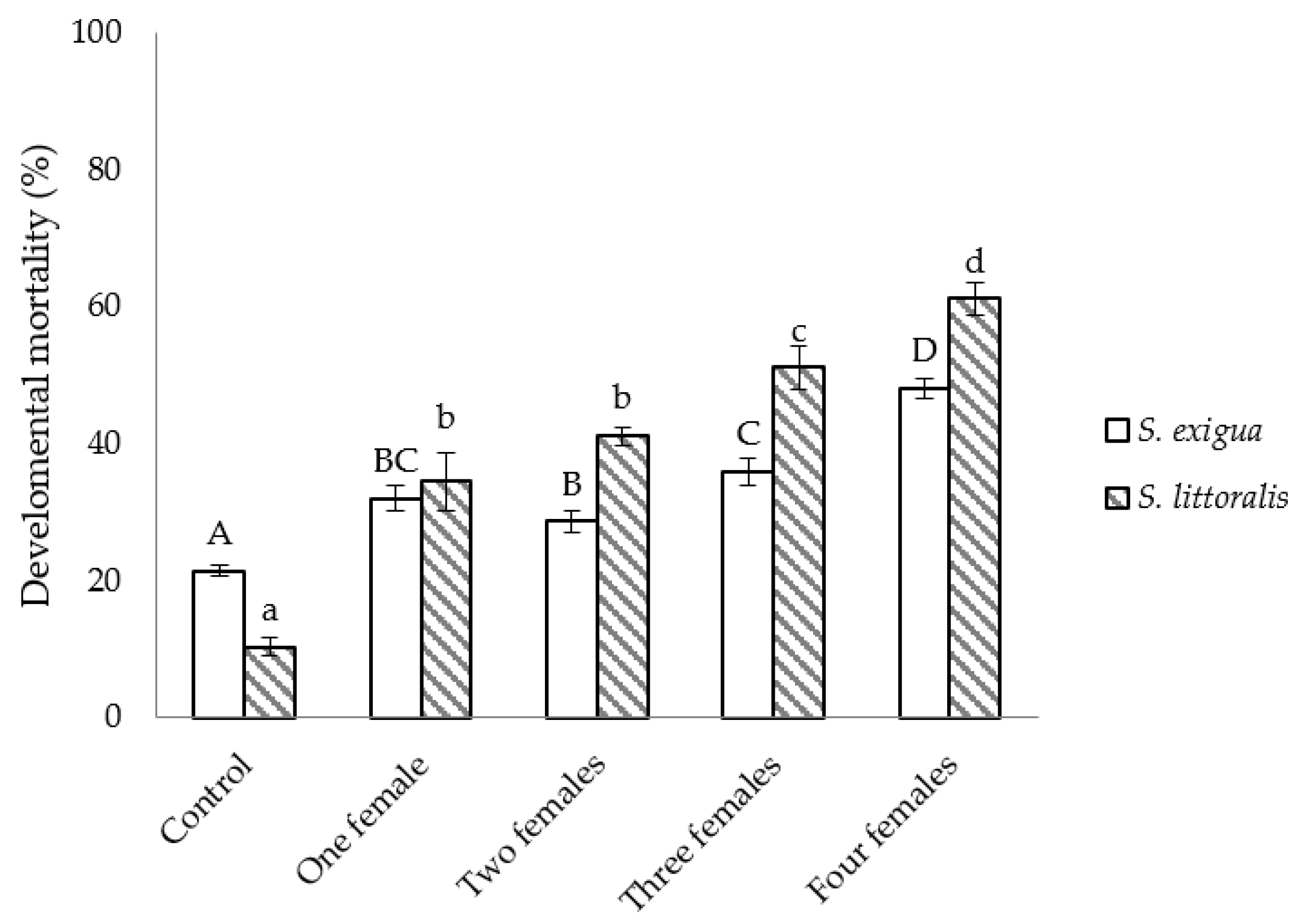
3.2. Semi-Field Studies on the Efficacy of C. inanitus in S. exigua and S. littoralis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- EPPO. PM 7/124 (1) Spodoptera littoralis, Spodoptera litura, Spodoptera frugiperda, Spodoptera eridania. EPPO Bull. 2015, 45, 410–444. [Google Scholar] [CrossRef]
- Gabarra, R.; Arnó, J.; Riudavets, J. Tomate. In Control Biológico de Plagas Agrícolas; Jacas, J., Urbaneja, A., Eds.; Phytoma España: Valencia, Spain, 2008; pp. 410–422. (In Spanish) [Google Scholar]
- van der Blom, J. Pimiento bajo abrigo. In Control Biológico de Plagas Agrícolas; Jacas, J., Urbaneja, A., Eds.; Phytoma España: Valencia, Spain, 2008; pp. 399–409. (In Spanish) [Google Scholar]
- Robledo Camacho, A.; van der Blom, J.; Sánchez Martínez, J.A.; Torres Giménez, S. Biological Control in Horticultural Crops; Coexphal: Almería, Spain, 2009. (In Spanish) [Google Scholar]
- Colomer, I.; Aguado, P.; Medina, P.; Heredia, R.M.; Fereres, A.; Belda, J.E.; Viñuela, E. Field trail measuring the compatibility of methoxyfenozide and flonicamid with Orius laevigatus Fieber (Hemiptera: Anthocoridae) and Amblyseius swirskii (Athias-Henriot) (Acari: Phytoseiidae) in a commercial pepper greenhouse. Pest Manag. Sci. 2011, 67, 1237–1244. [Google Scholar] [CrossRef]
- EPPO. A2 Quarantine List. Available online: https://www.eppo.int/ACTIVITIES/plant_quarantine/A2_list (accessed on 1 June 2021).
- CABI. Invasive Species Compendium. Spodoptera exigua (Beet Armyworm). Available online: https://www.cabi.org/isc/datasheet/29808 (accessed on 1 June 2021).
- CABI. Invasive Species Compendium. Spodoptera littoralis (Cotton Leafworm). Available online: https://www.cabi.org/isc/datasheet/51070 (accessed on 1 June 2021).
- FAUNA EUROPAEA. Spodoptera littoralis. Available online: https://fauna-eu.org (accessed on 1 June 2021).
- OJEU (Official Journal of the European Union). Directive 2009/128/EC of the European Parliament and of the Council of 21 October 2009, establishing a framework for Community action to achieve the sustainable use of pesticides. Off. J. Eur. Union 2009, 309, 71–86. [Google Scholar]
- IRAC. Insecticide Resistance Action Committee. IRAC Mode of Action Classification Scheme, Version 10.1; Available online: https://irac-online.org/mode-of-action (accessed on 1 June 2021).
- MAPA (Spanish Ministry of Agriculture). Phytosanitary Products Registration. Available online: https://www.mapa.gob.es/es/agricultura/temas/sanidad-vegetal/productos-fitosanitarios/registro/menu.asp (accessed on 30 May 2021). (In Spanish).
- Smagghe, G.; Pineda, S.; Carton, B.; del Estal, P.; Budia, F.; Viñuela, E. Toxicity and kinetics of methoxyfenozide in greenhouse-selected Spodoptera exigua (Lepidoptera: Noctuidae). Pest Manag. Sci. 2003, 59, 1203–1209. [Google Scholar] [CrossRef]
- Cabello, T. Control biológico de Noctuidos y otros Lepidópteros. In Control Biológico de Plagas Agrícolas; Jacas, J., Urbaneja, A., Eds.; Phytoma España: Valencia, Spain, 2008; pp. 279–306. (In Spanish) [Google Scholar]
- Quiu, B.; Zhong-Shi, Z.; Shu-Ping, L.; Zai-Fu, X. Effect of temperature on development, survival and fecundity of Microplitis manila (Hymenoptera: Braconidae). Environ. Entomol. 2012, 41, 657–664. [Google Scholar] [CrossRef]
- Che, W.; Shi, T.; Wu, Y.; Yang, Y. Insecticide resistance status of field populations of Spodoptera exigua (Lepidoptera: Noctuidae) from China. J. Econ. Entomol. 2013, 106, 1855–1862. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.M.; Zhao, Y.X.; Sun, H.; Ni, H.; Liu, C.; Wang, X.; Gao, C.F. Monitoring and mechanisms of insecticide resistance in Spodoptera exigua (Lepidoptera: Noctuidae) with special reference to diamides. Pestic. Biochem. Phys. 2021, 174, 104831. [Google Scholar] [CrossRef] [PubMed]
- MAPA (Spanish Ministry of Agriculture). Phytosanitary Control Tools. Available online: https://www.mapa.gob.es/app/omdfocb/default.aspx (accessed on 1 September 2021). (In Spanish).
- García-Martín, M.; Gámez, M.; Torres-Ruiz, A.; Cabello, T. Functional response of Chelonus oculator (Hymenoptera: Braconidae) to temperature and its consequences to parasitism. Community Ecol. 2008, 9, 45–51. [Google Scholar] [CrossRef]
- Martín Gil, A.; Alcázar Alba, M.D.; Trujillo Giménez, E. Guide of Integrated Crop Protection in Solanaceae; MAPA: Madrid, Spain, 2021; p. 239. (In Spanish) [Google Scholar]
- Grossnikalus-Bürgin, C.; Wyler, T.; Pfister-Wilhelm, R.; Lanzrein, B. Biology and morphology of the parasitoid Chelonus inanitus and effects on the development of its host Spodoptera littoralis. Invertebr. Reprod. Dev. 1994, 25, 143–158. [Google Scholar] [CrossRef]
- Gauld, I.; Bolton, B. The Hymenoptera; Oxford University Press: Oxford, UK, 1996. [Google Scholar]
- García-Marí, F.; Costa-Comelles, J.; Ferragut, F.J. Las Plagas Agrícolas; Agropubli SL Phytoma España: Valencia, Spain, 1994. (In Spanish) [Google Scholar]
- CABI. Chelonus inanitus. Available online: https://www.cabi.org/isc/datasheet/12711 (accessed on 15 May 2021).
- Ohsaki, B.; Shingyouchi, T.; Sato, Y.; Kainoh, Y. Host recognition by the egg larval parasitoid Chelonus inanitus: Effects of physical and chemical cues. Entomol. Exp. Appl. 2020, 168, 742–751. [Google Scholar] [CrossRef]
- Hegazi, E.M.; Altahtawy, M.; Hammad, S.M.; El-Sawaf, S.K. Notes on the biology of Chelonus inanitus (L.) (Hymen., Braconidae). Z. Angew. Entomol. 1974, 75, 291–294. [Google Scholar] [CrossRef]
- Charnov, E.L. Sex ratio in aged-structured populations. In The Theory of Sex Allocation; Charnov, E.L., Ed.; Princeton University Press: Princeton, NJ, USA, 2020; pp. 93–103. [Google Scholar]
- Bézier, A.; Annaheim, M.; Herbinière, J.; Wetterwald, C.; Gyapay, G.; Bernard-Samain, S.; Wincker, P.; Roditi, I.; Heller, M.; Belghazi, M.; et al. Polydnaviruses of braconid wasps derive from an ancestral Nudivirus. Science 2009, 323, 926–930. [Google Scholar] [CrossRef] [Green Version]
- Kaeslin, M.; Reinhard, M.; Bühler, D.; Roth, T.; Pfister-Wilhelm, R.; Lanzrein, B. Venom of the egg-larval parasitoid Chelonus inanitus is a complex mixture and has multiple biological effects. J. Insect Physiol. 2010, 56, 686–694. [Google Scholar] [CrossRef] [PubMed]
- Kaeslin, M.; Pfister-Wilhelm, R.; Lanzrein, B. Influence of the parasitoid Chelonus inanitus and its polydnavirus on host nutritional physiology and implications for parasitoid development. J. Insect Physiol. 2005, 51, 1330–1339. [Google Scholar] [CrossRef]
- Kaeslin, M.; Wehrle, I.; Grossniklaus-Bürgin, C.; Wyler, T.; Guggisberg, U.; Schittny, J.C.; Lanzrein, B. Stage-dependent strategies of host invasion in the egg-larval parasitoid Chelonus inanitus. J. Insect Physiol. 2005, 51, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Lanzrein, B.; Hammock, B. Degradation of juvenile hormone III in vitro by non-parasitized and parasitized Spodoptera exigua (Noctuidae) and by the endoparasitoid Chelonus inanitus (Braconidae). J. Insect Physiol. 1995, 41, 993–1000. [Google Scholar] [CrossRef]
- Pfister-Wilhem, R.; Lanzrein, B. Stage dependent influences of polydnaviruses and the parasitoid larva on host ecdysteroids. J. Insect Physiol. 2009, 55, 707–715. [Google Scholar] [CrossRef]
- Poitout, S.; Bues, R. Rearing of lepidopteran larvae of twenty-eight Noctuidae and two Arctidae species on artificial diet. Rearing details per species. Ann. Zool. Ecol. Anim. 1974, 6, 431–441. (In French) [Google Scholar]
- Statgraphics Technologies Inc. Statgraphics Centurion 18; Statgraphics Technologies Inc.: The Plains, VA, USA, 2020. [Google Scholar]
- Schneider, M.I.; Viñuela, E. Improvements in rearing method for Hyposoter didymator (Hymenoptera: Ichneumonidae), considering sex allocation and sex determination theories used for Hymenoptera. Biol. Control 2007, 43, 271–277. [Google Scholar] [CrossRef]
- Etzel, L.K.; Legner, E. Culture and colonization. In Handbook of Biological Control: Principles and Applications of Biological Control; Bellows, T.S., Fisher, T.S., Eds.; Elsevier: Amsterdam, The Netherlands, 1999; pp. 125–197. [Google Scholar]
- Zhou, Y.; Abram, P.K.; Boivin, G.; Brodeur, J. Increasing host age does not have the expected negative effects on the fitness parameters of an egg parasitoid. Entomol. Exp. Appl. 2014, 151, 106–111. [Google Scholar] [CrossRef]
- Vinson, S.B.; Iwantsch, G.F. Host suitability for insect parasitoids. Ann. Rev. Entomol. 1980, 25, 397–419. [Google Scholar] [CrossRef]
- Rechav, Y. Biological and ecological studies of the parasitoid Chelonus inanitus [Hym.: Braconidae] in Israel. IV. Oviposition, host preferences and sex ratio. Entomophaga 1978, 23, 95–102. [Google Scholar] [CrossRef]
- King, B.H. Breeding strategies in females of the parasitoid wasp Spalangia endius: Effects of mating status and size. J. Insect Behav. 2002, 15, 181–193. [Google Scholar] [CrossRef]
- Ode, P.J.; Heinz, K.M. Host-size-dependent sex ratio theory and improving mass-reared parasitoid sex ratios. Biol. Control 2002, 24, 31–41. [Google Scholar] [CrossRef]
- Fuester, R.W.; Swan, K.S.; Dunning, K.; Taylor, P.B.; Ramaseshiah, G. Male-biased sex ratios in Glyptapanteles flavicoxis (Hymenoptera: Braconidae), a parasitoid of the gypsy moth (Lepidoptera: Lymantriidae). Ann. Entomol. Soc. Am. 2003, 96, 553–559. [Google Scholar] [CrossRef] [Green Version]
- Azam, A.; Kunimi, Y.; Inoue, M.N.; Nakai, M. Effect of granulovirus infection of Spodoptera litura (Lepidoptera: Noctuidae) larvae on development of the endoparasitoid Chelonus inanitus (Hymenoptera: Braconidae). Appl. Entomol. Zool. 2016, 51, 479–488. [Google Scholar] [CrossRef]
- Rechav, Y.; Orion, T. The development of the immature stages of Chelonus inanitus. Ann. Entomol. Soc. Am. 1975, 68, 457–462. [Google Scholar] [CrossRef]
- van Lenteren, J.C.; Hale, A.; Klapwijk, J.N.; Van Schelt, J.; Steinberg, S. Guidelines for quality control of commercially produced natural enemies. In Quality Control and Production of Biological Control Agents: Theory and Testing Procedures; van Lenteren, J.C., Ed.; CABI: London, UK, 2003. [Google Scholar]
- Ameri, M.; Rasekh, A.; Michaud, J.P.; Allahyari, H. Morphometric indicators for quality assessment in the aphid parasitoid, Lysiphlebus fabarum (Braconidae: Aphidiinae). Eur. J. Entomol. 2013, 110, 519–525. [Google Scholar] [CrossRef] [Green Version]
- Azidah, A.A.; Sofian-Azirun, M. Size of Spodoptera exigua (Lepidoptera: Noctuidae) larvae reared on various host plants. Malays. J. Sci. 2007, 26, 93–96. [Google Scholar]
- Roque-Romero, L.; Cisneros, J.; Rojas, J.C.; Ortiz-Carreon, F.R.; Malo, E.A. Attraction of Chelonus insularis to host and host habitat volatiles during the search of Spodoptera frugiperda eggs. Biol. Control 2020, 140, 104100. [Google Scholar] [CrossRef]
- Abram, P.K.; Brodeur, J.; Urbaneja, A.; Tena, A. Nonreproductive effects of insect parasitoids on their hosts. Ann. Rev. Entomol. 2019, 64, 259–276. [Google Scholar] [CrossRef]
- Kaser, J.M.; Nielsen, A.L.; Abram, P.K. Biological control effects of non-reproductive host mortality caused by insect parasitoids. Ecol. Appl. 2018, 28, 1081–1093. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Meng, L.; Li, B. Non-reproductive effects of two parasitoid species on the oriental armyworm Mythimna separata on wheat and maize plants. BioControl 2019, 64, 115–124. [Google Scholar] [CrossRef]
- Abram, P.K.; Brodeur, J.; Burte, V.; Boivin, G. Parasitoid-induced host egg abortion: An underappreciated component of biological control services provided by egg parasitoids. Biol. Control 2016, 98, 52–60. [Google Scholar] [CrossRef]
- Morales, J.; Medina, P.; Viñuela, E. The influence of two endoparasitic wasps, Hyposoter didymator and Chelonus inanitus on the growth and food consumption of their host larva Spodoptera littoralis. BioControl 2007, 52, 145–160. [Google Scholar] [CrossRef]
- Rechav, Y. Biological and ecological studies of the parasitoid Chelonus inanitus (L) (Hymenoptera: Braconidae) in Israel II. Releases of adults in a cotton field. J. Entomol. Soc. South. Afr. 1976, 39, 83–85. [Google Scholar]

{kind=link}
{kind=link}
| Parameter | NFP | HA | Statistics | NFP | HA | NFP × HA | ||
|---|---|---|---|---|---|---|---|---|
| 4-Day-Old | 1-Day-Old | Mean | ||||||
| C. inanitus adults (%) | 1–3 females | 37.1 ± 5.8 | 34.8 ± 3.9 | 35.9 ± 3.4 | F | 3.75 | 4.52 | 6.80 |
| 5 females | 33.9 ± 4.8 a | 56.5 ± 4.4 b | 45.2 ± 4.1 | p | 0.06 | 0.04 | 0.01 | |
| Mean | 35.5 ± 3.7 | 45.6 ± 3.8 | ||||||
| C. inanitus females (%) | 1–3 females | 25.2 ± 4.1 | 15.0 ± 4.2 | 20.1 ± 3.1 | F | 3.11 | 5.67 | 0.02 |
| 5 females | 31.9 ± 4.1 | 22.7 ± 3.9 | 27.3 ± 3.0 | p | 0.09 | 0.02 | 0.90 | |
| Mean | 28.6 ± 2.9 b | 18.8 ± 2.9 a | ||||||
| E. kuehniella adults (%) | 1–3 females | 38.9 ± 5.7 | 46.7 ± 5.0 | 42.8 ± 3.8 B | F | 17.8 | 0.03 | 3.08 |
| 5 females | 26.8 ± 5.2 | 17.0 ± 3.7 | 21.9 ± 3.3 A | p | <0.001 | 0.84 | 0.09 | |
| Mean | 32.8 ± 4.0 | 31.8 ± 4.6 | ||||||
| Developmental mortality (%) 1 | 1–3 females | 24.1 ± 3.2 | 18.6 ± 3.2 | 21.4 ± 2.2 A | F | 4.42 | 2.15 | 0.16 |
| 5 females | 39.6 ± 4.9 | 26.6 ± 3.0 | 33.1 ± 3.9 B | p | 0.04 | 0.15 | 0.69 | |
| Mean | 31.8 ± 7.1 | 22.6 ± 2.3 | ||||||
| Sex | HA | Statistics | Sex | HA | Sex × HA | ||
|---|---|---|---|---|---|---|---|
| 4-Day-Old | 1-Day-Old | Mean | |||||
| Male | 43.4 ± 0.4 | 46.9 ± 1.0 | 45.3 ± 0.7 A | F | 10.51 | 18.38 | 1.12 |
| Female | 45.8 ± 1.2 | 51.5 ± 1.6 | 48.3 ± 1.2 B | p | 0.03 | <0.001 | 0.30 |
| Mean | 44.6 ± 0.7 a | 48.8 ± 1.0 b | |||||
| Sex | HS | Statistics | Sex | HS | Sex × HS | |||
|---|---|---|---|---|---|---|---|---|
| S. exigua | S. littoralis | E. kuehniella | Mean | |||||
| Male | 4.61 ± 0.06 | 4.89 ± 0.04 | 4.49 ± 0.04 | 4.66 ± 0.06 | F | 2.72 | 35.83 | 0.38 |
| Female | 4.68 ± 0.08 | 5.00 ± 0.01 | 4.52 ± 0.07 | 4.73 ± 0.08 | p | 0.12 | <0.001 | 0.69 |
| Mean | 4.64 ± 0.05 b | 4.95 ± 0.05 c | 4.50 ± 0.04 a | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magaña, A.J.; Dáder, B.; Sancho, G.; Adán, Á.; Morales, I.; Viñuela, E. Comparison of the Parasitization of Chelonus inanitus L. (Hymenoptera: Braconidae) in Two Spodoptera Pests and Evaluation of the Procedure for Its Production. Insects 2022, 13, 99. https://doi.org/10.3390/insects13010099
Magaña AJ, Dáder B, Sancho G, Adán Á, Morales I, Viñuela E. Comparison of the Parasitization of Chelonus inanitus L. (Hymenoptera: Braconidae) in Two Spodoptera Pests and Evaluation of the Procedure for Its Production. Insects. 2022; 13(1):99. https://doi.org/10.3390/insects13010099
Chicago/Turabian StyleMagaña, Antonio Jesús, Beatriz Dáder, Gonzalo Sancho, Ángeles Adán, Ignacio Morales, and Elisa Viñuela. 2022. "Comparison of the Parasitization of Chelonus inanitus L. (Hymenoptera: Braconidae) in Two Spodoptera Pests and Evaluation of the Procedure for Its Production" Insects 13, no. 1: 99. https://doi.org/10.3390/insects13010099
APA StyleMagaña, A. J., Dáder, B., Sancho, G., Adán, Á., Morales, I., & Viñuela, E. (2022). Comparison of the Parasitization of Chelonus inanitus L. (Hymenoptera: Braconidae) in Two Spodoptera Pests and Evaluation of the Procedure for Its Production. Insects, 13(1), 99. https://doi.org/10.3390/insects13010099