
Western Corn Rootworm, Plant and Microbe Interactions: A Review and Prospects for New Management Tools


Abstract
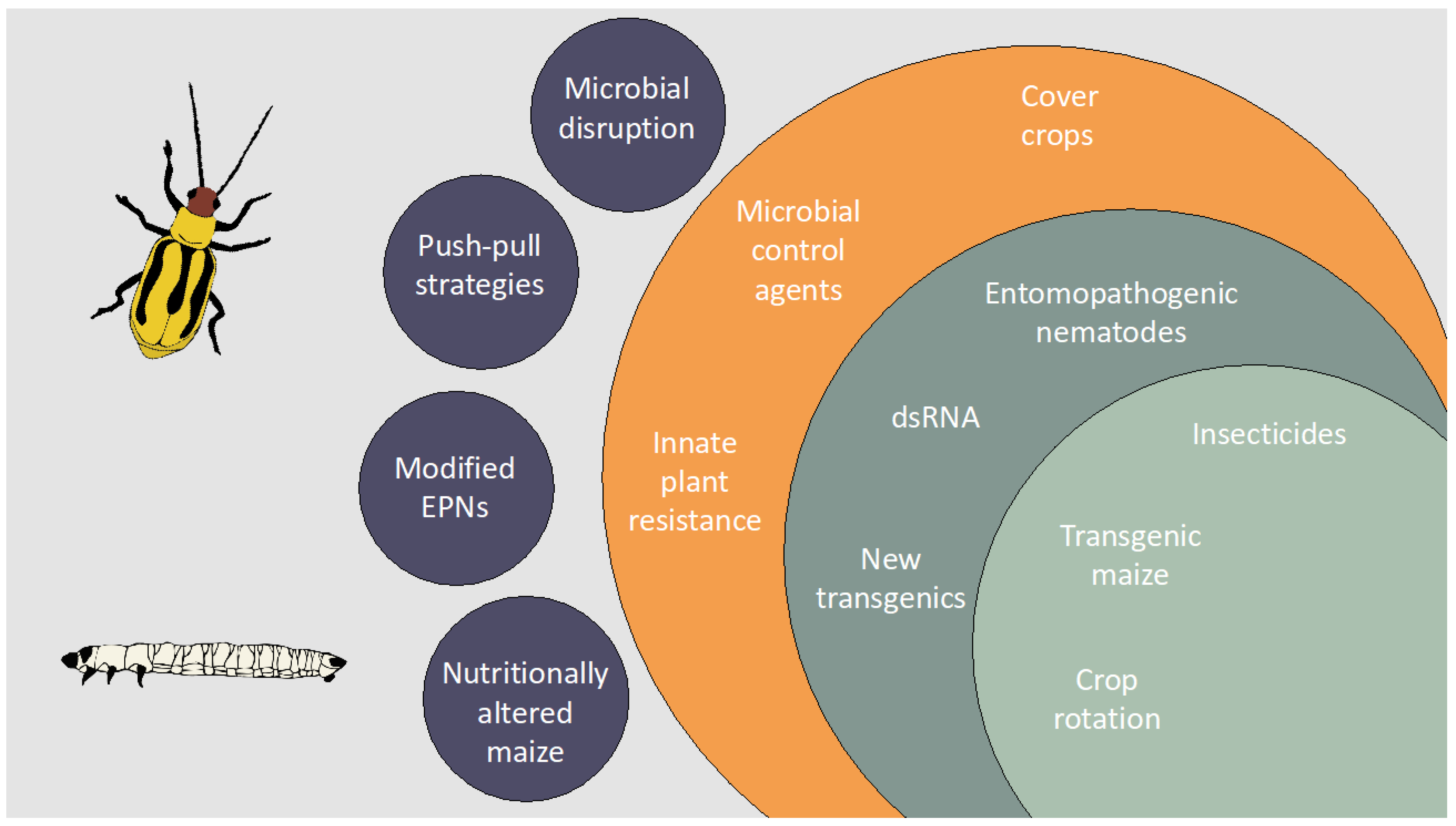
Simple Summary
Abstract
1. Introduction
2. Disrupting WCR Establishment
3. Selecting for Maize Lines with Effective Defenses against WCR
4. Altering Maize Nutritional Value for the WCR
5. Plant-Mediated RNA Interference
6. Enhancing Plant Health-Promoting Microbes
7. Using Soil Microbials to Disrupt WCR Gut Microbiome
8. Using Pathogenic Microbials to Reduce WCR Populations
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gillette, C.P. Diabrotica virgifera Lec. as a corn rootworm. J. Econ. Entomol. 1912, 5, 364–366. [Google Scholar] [CrossRef]
- Metcalf, R.L. Foreword. In Methods for the Study of the Pest Diabrotica; Krysan, J.L., Miller, T.A., Eds.; Springer: New York, NY, USA, 1986; pp. 7–15. [Google Scholar]
- Chiang, H.C. Bionomics of the northern and western corn rootworms. Annu. Rev. Entomol. 1973, 18, 47–72. [Google Scholar] [CrossRef]
- Miller, N.; Estoup, A.; Toepfer, S.; Bourguet, D.; Lapchin, L.; Derridj, S.; Kim, K.S.; Reynaud, P.; Furlan, L.; Guillemaud, T. Multiple transatlantic introductions of the western corn rootworm. Science 2005, 310, 992. [Google Scholar] [CrossRef]
- Wechsler, S.; Smith, D. Has resistance taken root in US corn fields? Demand for insect control. Am. J. Agric. Econ. 2018, 100, 1136–1150. [Google Scholar] [CrossRef]
- Veres, A.; Wyckhuys, K.A.G.; Kiss, J.; Tóth, F.; Burgio, G.; Pons, X.; Avilla, C.; Vidal, S.; Razinger, J.; Bazok, R.; et al. An update of the Worldwide Integrated Assessment (WIA) on systemic pesticides. Part 4: Alternatives in major cropping systems. Environ. Sci. Pollut. Res. 2020, 27, 29867–29899. [Google Scholar] [CrossRef]
- Robert, C.A.M.; Veyrat, N.; Glauser, G.; Marti, G.; Doyen, G.R.; Villard, N.; Gaillard, M.D.P.; Köllner, T.G.; Giron, D.; Body, M.; et al. A specialist root herbivore exploits defensive metabolites to locate nutritious tissues. Ecol. Lett. 2012, 15, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Ball, H.J.; Weekman, G.T. Insecticide resistance in the adult western corn rootworm in Nebraska. J. Econ. Entomol. 1962, 55, 439–441. [Google Scholar] [CrossRef]
- Meinke, L.J.; Siegfried, B.D.; Wright, R.J.; Chandler, L.D. Adult susceptibility of Nebraska western corn rootworm (Coleoptera: Chrysomelidae) populations to selected insecticides. J. Econ. Entomol. 1998, 91, 594–600. [Google Scholar] [CrossRef]
- Pereira, A.E.; Wang, H.; Zukoff, S.N.; Meinke, L.J.; French, B.W.; Siegfried, B.D. Evidence of field-evolved resistance to bifenthrin in western corn rootworm (Diabrotica virgifera virgifera LeConte) populations in western Nebraska and Kansas. PLoS ONE 2015, 10, e0142299. [Google Scholar] [CrossRef]
- Meihls, L.N.; Higdon, M.L.; Siegfried, B.D.; Miller, N.J.; Sappington, T.W.; Ellersieck, M.R.; Spencer, T.A.; Hibbard, B.E. Increased survival of western corn rootworm on transgenic corn within three generations of on-plant greenhouse selection. Proc. Natl. Acad. Sci. USA 2008, 105, 19177. [Google Scholar] [CrossRef]
- Meihls, L.N.; Higdon, M.L.; Ellersieck, M.; Hibbard, B.E. Selection for resistance to mCry3A-expressing transgenic corn in western corn rootworm. J. Econ. Entomol. 2011, 104, 1045–1054. [Google Scholar] [CrossRef] [PubMed]
- Deitloff, J.; Dunbar, M.W.; Ingber, D.A.; Hibbard, B.E.; Gassmann, A.J. Effects of refuges on the evolution of resistance to transgenic corn by the western corn rootworm, Diabrotica virgifera virgifera LeConte. Pest Manag. Sci. 2016, 72, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Frank, D.L.; Zukoff, A.; Barry, J.; Higdon, M.L.; Hibbard, B.E. Development of resistance to eCry3.1Ab-expressing transgenic maize in a laboratory-selected population of western corn rootworm (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2013, 106, 2506–2513. [Google Scholar] [CrossRef]
- Levine, E.; Spencer, J.L.; Isard, S.A.; Onstad, D.W.; Gray, M.E. Adaptation of the western corn rootworm to crop rotation: Evolution of a new strain in response to a management practice. Am. Entomol. 2002, 48, 94–107. [Google Scholar] [CrossRef]
- Khajuria, C.; Ivashuta, S.; Wiggins, E.; Flagel, L.; Moar, W.; Pleau, M.; Miller, K.; Zhang, Y.; Ramaseshadri, P.; Jiang, C.; et al. Development and characterization of the first dsRNA-resistant insect population from western corn rootworm, Diabrotica virgifera virgifera LeConte. PLoS ONE 2018, 13, 7059. [Google Scholar] [CrossRef]
- Meinke, L.J.; Souza, D.; Siegfried, B.D. The Use of Insecticides to Manage the Western Corn Rootworm, Diabrotica virgifera virgifera, LeConte: History, Field-Evolved Resistance, and Associated Mechanisms. Insects 2021, 12, 112. [Google Scholar] [CrossRef]
- EU No. 485. Available online: https://eur-lex.europa.eu/eli/reg_impl/2013/485/oj (accessed on 15 January 2021).
- Storer, N.P. A spatially explicit model simulating western corn rootworm (Coleoptera: Chrysomelidae) adaptation to insect-resistant maize. J. Econ. Entomol. 2003, 5, 1530–1547. [Google Scholar] [CrossRef]
- Hibbard, B.E.; Higdon, M.L.; Duran, D.P.; Schweikert, Y.M.; Ellersieck, M.R. Role of egg density on establishment and plant-to-plant movement by western corn rootworm larvae (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2004, 97, 871–882. [Google Scholar] [CrossRef]
- Hibbard, B.E.; Meihls, L.N.; Ellersieck, M.R.; Onstad, D.W. Density-dependent and density-independent mortality of the western corn rootworm: Impact on dose calculations of rootworm-resistant Bt corn. J. Econ. Entomol. 2010, 103, 77–84. [Google Scholar] [CrossRef]
- Strnad, S.P.; Bergman, M.K. Movement of first-instar western corn rootworms (Coleoptera: Chrysomelidae) in soil. Environ. Entomol. 1987, 4, 975–978. [Google Scholar] [CrossRef]
- Ellsbury, M.M.; Schumacher, T.E.; Gustin, R.D.; Woodson, W.D. Soil compaction effect on corn rootworm populations in maize artificially infested with eggs of western corn rootworm (Coleoptera: Chrysomelidae). Environ. Entomol. 1994, 4, 943–948. [Google Scholar] [CrossRef]
- Macdonald, P.J.; Ellis, C.R. Survival time of unfed, first-instar western corn rootworm (Coleoptera: Chrysomelidae) and the effects of soil type, moisture, and compaction on their mobility in soil. Environ. Entomol. 1990, 3, 666–671. [Google Scholar] [CrossRef]
- Chaddha, S. Influence of Placement of Western Corn Rootworm Eggs on Survivorship, Root Injury and Yield. Master’s Thesis, University of Minnesota, Minneapolis, MN, USA, 1990. [Google Scholar]
- Gustin, R.D.; Schumacher, T.E. Relationship of some soil pore parameters to movement of first-instar western corn rootworm (Coleoptera: Chrysomelidae). Environ. Entomol. 1989, 18, 343–346. [Google Scholar] [CrossRef]
- Strnad, S.P.; Bergman, M.K. Distribution and orientation of western corn rootworm (Coleoptera: Chrysomelidae) larvae in corn roots. Environ. Entomol. 1987, 16, 1193–1198. [Google Scholar] [CrossRef]
- Arce, C.C.M.; Theepan, V.; Schimmel, B.C.J.; Jaffuel, G.; Erb, M.; Machado, R.A.R. Plant-derived CO2 mediates long-distance host location and quality assessment by a root herbivore. BioRxiv 2020. [Google Scholar] [CrossRef]
- Bernklau, E.J.; Bjostad, L.B. Behavioral responses of first-instar western corn rootworm (Coleoptera: Chrysomelidae) to carbon dioxide in a glass bead bioassay. J. Econ. Entomol. 1998, 91, 444–456. [Google Scholar] [CrossRef]
- Bernklau, E.J.; Fromm, E.A.; Bjostad, L.B. Disruption of host location of western corn rootworm larvae (Coleoptera: Chrysomelidae) with carbon dioxide. J. Econ. Entomol. 2004, 97, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Schumann, M.; Patel, A.; Vidal, S. Soil application of an encapsulated CO2 source and its potential for management of western corn rootworm larvae. J. Econ. Entomol. 2014, 107, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Robert, C.A.M.; Erb, M.; Duployer, M.; Zwahlen, C.; Doyen, G.R.; Turlings, T.C.J. Herbivore-induced plant volatiles mediate host selection by a root herbivore. N. Phytol. 2012, 194, 1061–1069. [Google Scholar] [CrossRef]
- Robert, C.A.M.; Erb, M.; Hibbard, B.E.; Wade French, B.; Zwahlen, C.; Turlings, T.C.J. A specialist root herbivore reduces plant resistance and uses an induced plant volatile to aggregate in a density-dependent manner. Funct. Ecol. 2012, 26, 1429–1440. [Google Scholar] [CrossRef]
- Hiltpold, I.; Hibbard, B.E. Neonate larvae of the specialist herbivore Diabrotica virgifera virgifera do not exploit the defensive volatile (E)-β-caryophyllene in locating maize roots. J. Pest Sci. 2016, 89, 853–858. [Google Scholar] [CrossRef]
- Strnad, S.P.; Dunn, P.E. Host search behaviour of neonate western corn rootworm (Diabrotica virgifera virgifera). J. Insect Physiol. 1990, 36, 201–205. [Google Scholar] [CrossRef]
- Bernklau, E.J.; Hibbard, B.E.; Bjostad, L.B. Isolation and characterization of host recognition cues in corn roots for larvae of the western corn rootworm (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2013, 106, 2354–2363. [Google Scholar] [CrossRef]
- Bernklau, E.J.; Hibbard, B.E.; Dick, D.L.; Rithner, C.D.; Bjostad, L.B. Monogalactosyldiacylglycerols as host recognition cues for western corn rootworm larvae (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2015, 108, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Bernklau, E.J.; Bjostad, L.B. Identification of feeding stimulants in corn roots for western corn rootworm (Coleoptera: Chrysomelidae) larvae. J. Econ. Entomol. 2008, 101, 341–351. [Google Scholar] [CrossRef]
- Bernklau, E.J.; Hibbard, B.E.; Bjostad, L.B. Sugar preferences of western corn rootworm larvae in a feeding stimulant blend. J. Appl. Entomol. 2018, 142, 947–958. [Google Scholar] [CrossRef]
- Bernklau, E.J.; Hibbard, B.E.; Bjostad, L.B. Toxic and behavioural effects of free fatty acids on western corn rootworm (Coleoptera: Chrysomelidae) larvae. J. Appl. Entomol. 2016, 140, 725–735. [Google Scholar] [CrossRef]
- Hu, L.; Mateo, P.; Ye, M.; Zhang, X.; Berset, J.D.; Handrick, V.; Radisch, D.; Grabe, V.; Köllner, T.G.; Gershenzon, J.; et al. Plant iron acquisition strategy exploited by an insect herbivore. Science 2018, 361, 694–697. [Google Scholar] [CrossRef] [PubMed]
- Machado, R.A.R.; Theepan, V.; Robert, C.A.M.; Züst, T.; Hu, L.; Su, Q.; Schimmel, B.C.J.; Erb, M. Complex plant metabolomes guide fitness-relevant foraging decisions of a specialist herbivore. BioRxiv 2020. [Google Scholar] [CrossRef]
- Bernklau, E.J.; Hibbard, B.E.; Norton, A.P.; Bjostad, L.B. Methyl anthranilate as a repellent for western corn rootworm larvae (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2016, 109, 1683–1690. [Google Scholar] [CrossRef]
- Wouters, F.C.; Blanchette, B.; Gershenzon, J.; Vassão, D.G. Plant defense and herbivore counter-defense: Benzoxazinoids and insect herbivores. Phytochem. Rev. 2016, 15, 1127–1151. [Google Scholar] [CrossRef] [PubMed]
- Bernklau, E.J.; Bjostad, L.B.; Hibbard, B.E. Synthetic feeding stimulants enhance insecticide activity against western corn rootworm larvae, Diabrotica virgifera virgifera (Coleoptera: Chrysomelidae). J. Appl. Entomol. 2011, 135, 47–54. [Google Scholar] [CrossRef]
- Hibbard, B.E.; Peairs, F.B.; Pilcher, S.D.; Schroeder, M.E.; Jewett, D.K.; Bjostad, L.B. Germinating corn extracts and 6-methoxy-2-benzoxazolinone: Western corn rootworm (Coleoptera: Chrysomelidae) larval attractants evaluated with soil insecticides. J. Econ. Entomol. 1995, 88, 716–724. [Google Scholar] [CrossRef]
- Khan, Z.R.; Pickett, J.A. The “push-pull” strategy for stemborer management: A case study in exploiting biodiversity and chemical ecology. In Ecological Engineering for Pest Management: Advances in Habitat Manipulation for Arthropods; Gurr, G.M., Wratten, S.D., Altieri, M.A., Eds.; CABI: Oxon, UK, 2004; pp. 155–164. [Google Scholar]
- Bernklau, E.J.; Hibbard, B.E.; Bjostad, L.B. Repellent effects of methyl anthranilate on western corn rootworm larvae (Coleoptera: Chrysomelidae) in soil bioassays. J. Econ. Entomol. 2019, 112, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Puebla, F.A.A.; Bernal, J.S. Resistance and tolerance to root herbivory in maize were mediated by domestication, spread, and breeding. Front. Plant Sci. 2020, 27, 223. [Google Scholar] [CrossRef]
- Smith, C.M. Plant Resistance to Insects: A Fundamental Approach; Wiley: New York, NY, USA, 1989. [Google Scholar]
- Painter, R.H. Insect Resistance in Crop Plants; Macmillian: New York, NY, USA, 1951. [Google Scholar] [CrossRef]
- Howe, G.A.; Jander, G. Plant immunity to insect herbivores. Annu. Rev. Plant Biol. 2008, 59, 41–66. [Google Scholar] [CrossRef]
- Loranger, J.; Meyer, S.T.; Shipley, B.; Kattge, J.; Loranger, H.; Roscher, C.; Weisser, W.W. Predicting invertebrate herbivory from plant traits: Evidence from 51 grassland species in experimental monocultures. Ecology 2012, 93, 2674–2682. [Google Scholar] [CrossRef] [PubMed]
- Iason, G.R.; Dicke, M.; Hartley, S.E. The Ecology of Plant Secondary Metabolites: From Genes to Global Processes; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Hanley, M.E.; Lamont, B.B.; Fairbanks, M.M.; Rafferty, C.M. Plant structural traits and their role in anti-herbivore defence. Perspect. Plant Ecol. Evol. Syst. 2007, 8, 157–178. [Google Scholar] [CrossRef]
- Strauss, S.Y.; Agrawal, A.A. The ecology and evolution of plant tolerance to herbivory. Trends Ecol. Evol. 1999, 14, 179–185. [Google Scholar] [CrossRef]
- Newingham, B.A.; Callaway, R.M.; Bassirirad, H. Allocating nitrogen away from a herbivore: A novel compensatory response to root herbivory. Oecologia 2007, 153, 913–920. [Google Scholar] [CrossRef]
- Xue, K.; Serohijos, R.C.; Devare, M.; Duxbury, J.; Lauren, J.; Thies, J.E. Short-term carbon allocation and root lignin of Cry3Bb Bt and nonBt corn in the presence of corn rootworm. Appl. Soil Ecol. 2012, 57, 16–22. [Google Scholar] [CrossRef]
- Robert, C.A.M.; Ferrieri, R.A.; Schirmer, S.; Babst, B.A.; Schueller, M.J.; Machado, R.A.R.; Arce, C.C.M.; Hibbard, B.E.; Gershenzon, J.; Turlings, T.C.J.; et al. Induced carbon reallocation and compensatory growth as root herbivore tolerance mechanisms. Plant Cell Environ. 2014, 37, 2613–2622. [Google Scholar] [CrossRef]
- Robert, C.A.M.; Schirmer, S.; Barry, J.; French, W.B.; Hibbard, B.E.; Gershenzon, J. Belowground herbivore tolerance involves delayed overcompensatory root regrowth in maize. Entomol. Exp. Appl. 2015, 157, 113–120. [Google Scholar] [CrossRef]
- Bigger, J.H.; Holbert, J.R.; Flint, W.P.; Lang, A.L. Resistance of certain corn hybrids to attack of southern corn rootworm. J. Econ. Entomol. 1938, 21, 103–107. [Google Scholar] [CrossRef]
- Bigger, J.H.; Snelling, R.O.; Blanchard, R.A. Resistance of corn strains to the southern corn rootworm, Diabrotica duodecimpunctata F. J. Econ. Entomol. 1941, 34, 605–613. [Google Scholar] [CrossRef]
- Wilson, R.L.; Peters, D.C. Plant Introductions of Zea mays as sources of corn rootworm tolerance. J. Econ. Entomol. 1973, 66, 101–104. [Google Scholar] [CrossRef]
- Tollefson, J.J. Evaluating maize for resistance to Diabrotica virgifera virgifera Leconte (Coleoptera: Chrysomelidae). Maydica 2007, 52, 311–318. [Google Scholar]
- Branson, T.F. Resistance in the grass tribe Maydeae to larvae of the western corn rootworm. Ann. Entomol. Soc. Am. 1971, 64, 861–863. [Google Scholar] [CrossRef]
- Branson, T.F.; Guss, P.L. Potential for utilizing resistance from relatives of cultivated crops. Proc. North Cent. Branch Entomol. Soc. Am. 1972, 27, 91–95. [Google Scholar]
- Branson, T.F.; Reyes, R.J. The Association of Diabrotica spp. with Zea diploperennis. J. Kans. Entomol. Soc. 1983, 56, 97–99. [Google Scholar]
- Branson, T.F.; Fisher, J.R.; Kahler, A.L.; Sutter, G.R. Host plant resistance to corn rootworms. In Proceedings of the 17th Annual Illinois Corn Breeder School; Allerton House: Champaign, IL, USA, 1981. [Google Scholar]
- Branson, T.F.; Sutter, G.R.; Fisher, J.R. Comparison of a tolerant and a susceptible maize inbred under artificial infestations of Diabrotica virgifera virgifera: Yield and adult emergence. Environ. Entomol. 1982, 11, 371–372. [Google Scholar] [CrossRef]
- Branson, T.F. Larval feeding behavior and host-plant resistance in maize. In Methods for the Study of Pest Diabortica; Krysan, J.L., Miller, T.A., Eds.; Springer: New York, NY, USA, 1986; pp. 159–182. [Google Scholar]
- Fitzgerald, P.J.; Ortman, E.E. Breeding for resistance to the western corn rootworm. In Proceedings of the Annual Hybrid Corn Industry Research Conference; Heckendorn, W., Sutherland, J.I., Eds.; American Seed Trade Association: Washington, DC, USA, 1964; pp. 46–60. [Google Scholar]
- Fitzgerald, P.J.; Ortman, E.E. Two-year performance of inbreds and their single crosses grown under corn rootworm infestation. Proc. North Cent. Branch Entomol. Soc. Am. 1965, 20, 46–47. [Google Scholar]
- Hills, T.M.; Peters, D.C. A method of evaluating postplanting insecticide treatments for control of western corn rootworm larvae. J. Econ. Entomol. 1971, 64, 764–765. [Google Scholar] [CrossRef]
- Kahler, A.L.; Olness, A.E.; Sutter, G.R.; Dybing, C.D.; Devine, O.J. Root damage by western corn rootworm and nutrient content in maize. Agron. J. 1985, 77, 769–774. [Google Scholar] [CrossRef]
- Kahler, A.L.; Telkamp, R.E.; Penny, L.H.; Branson, T.F.; Fitzgerald, P.J. Registration of NGSDCRW1(S2)C4 Maize Germplasm. Crop Sci. 1985, 25, 202. [Google Scholar] [CrossRef]
- Ortman, E.E.; Gerloff, E.D. Rootworm resistance: Problems in measuring and its relationship to performance. In Proceedings of the 25th Annual Corn and Sorghum Research Conference, Washington, DC, USA, 8–10 December 1970; Sutherland, J.I., Falasca, R.J., Eds.; pp. 161–174. [Google Scholar]
- Ortman, E.E.; Branson, T.F. Growth pouches for studies of host plant resistance to larvae of corn rootworms. J. Econ. Entomol. 1976, 69, 380–382. [Google Scholar] [CrossRef]
- Ortman, E.E.; Branson, T.F.; Gerloff, E.D. Techniques, accomplishments, and future potential of host plant resistance to Diabrotica. In Proceedings of the Summer Institute on Biological Control of Plant Insects and Diseases; Maxwell, F.G., Harris, F.A., Eds.; University Press: Jackson, MS, USA, 1974; pp. 344–358. [Google Scholar]
- Riedell, W.E. Western corn rootworm damage in maize: Greenhouse technique and plant response. Crop Sci. 1989, 29, 412–415. [Google Scholar] [CrossRef]
- Riedell, W.E.; Evenson, P.D. Rootworm feeding tolerance in single-cross maize hybrids from different eras. Crop Sci. 1993, 33, 951–955. [Google Scholar] [CrossRef]
- Prischmann, D.A.; Dashiell, K.E.; Schneider, D.J.; Hibbard, B.E. Field screening maize germplasm for resistance and tolerance to western corn rootworms (Col.: Chrysomelidae). J. Appl. Entomol. 2007, 131, 406–415. [Google Scholar] [CrossRef]
- Assabgui, R.A.; Arnason, J.T.; Hamilton, R.I. Hydroxamic acid content in maize (Zea mays) roots of 18 Ontario recommended hybrids and prediction of antibiosis to the western corn rootworm, Diabrotica virgifera virgifera LeConte (Coleoptera: Chrysomelidae). Can. J. Plant Sci. 1993, 73, 359–363. [Google Scholar] [CrossRef]
- Assabgui, R.A.; Arnason, J.T.; Hamilton, R.I. Hydroxamic acid content and plant development of maize (Zea mays L.) in relation to damage by the western corn rootworm, Diabrotica virgifera virgifera LeConte. Can. J. Plant Sci. 1995, 75, 51–856. [Google Scholar] [CrossRef]
- Rogers, R.R.; Owens, J.C.; Tollefson, J.J.; Witkowski, J.F. Evaluation of commercial corn hybrids for tolerance to corn rootworms. Environ. Entomol. 1975, 4. [Google Scholar] [CrossRef]
- Assabgui, R.A.; Arnason, J.T.; Hamilton, R.I. Field evaluations of hydroxamic acids as antibiosis factors in elite maize inbreds to the western corn rootworm (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1995, 88, 1482–1493. [Google Scholar] [CrossRef]
- Xie, Y.S.; Arnason, J.T.; Philogene, B.J.R.; Lambert, J.D.H.; Atkinson, J.; Morand, P. Role of 2, 4-dihydroxy-7-methoxy-1, 4-benzoxazin-3-one (DIMBOA) in the resistance of maize to western corn rootworm, Diabrotica virgifera virgifera (Leconte) (Coleoptera: Chrysomelidae). Can. Entomol. 1990, 122, 1177–1186. [Google Scholar] [CrossRef]
- Xie, Y.; Arnason, T.J.; Philogène, B.J.R.; Olechowski, H.T.; Hamilton, R. Variation of Hydroxamic acid content in maize roots in relation to geographic origin of maize germplasm and resistance to western corn rootworm (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1992, 85, 2478–2485. [Google Scholar] [CrossRef]
- Xie, Y.; Arnason, J.T.; Philogéne, B.J.R.; Atkinson, J.; Morand, P. Behavioral responses of western corn rootworm larvae to naturally occurring and synthetic hydroxamic acids. J. Chem. Ecol. 1992, 18, 945–957. [Google Scholar] [CrossRef] [PubMed]
- Arnason, J.T.; Larsen, J.; Assabgui, R.; Xie, Y.; Atkinson, J.; Philogene, B.J.R.; Hamilton, R.I. Mechanisms of resistance in maize to western corn rootworm. In Insect resistant maize: Recent advances and utilization. In Proceedings of the International Symposium Held at the International Maize and Wheat Improvement Center (CIMMYT), El Bátan, Tunisia, 27 November–3 December 1994; pp. 96–100. [Google Scholar]
- Moellenbeck, D.J.; Bergvinson, B.D.; Darrah, L.L. Advances in rating and phytochemical screening for corn rootworm resistance. In Insect resistant maize: Recent advances and utilization. In Proceedings of the International Symposium Held at the International Maize and Wheat Improvement Center (CIMMYT), El Bátan, Tunisia, 27 November–3 December 1994; pp. 203–210. [Google Scholar]
- Praiswater, T.W.; Hibbard, B.E.; Barry, B.D.; Darrah, L.L.; Smith, V.A. An implement for dislodging maize roots from the soil for corn rootworm (Coleoptera: Chrysomelidae) damage evaluations. J. Kans. Entomol. Soc. 1997, 70, 335–338. [Google Scholar] [CrossRef]
- Hibbard, B.E.; Darrah, L.L.; Barry, B.D. Combining ability of resistance leads and identification of a new resistance source for western corn rootworm (Coleoptera: Chrysomelidae) larvae in corn. Maydica 1999, 44, 133–139. [Google Scholar]
- Hibbard, B.E.; Barry, B.D.; Darrah, L.L.; Jackson, J.J.; Chandler, L.D.; French, L.K.; Mihm, J.A. Controlled field infestations with western corn rootworm (Coleoptera: Chrysomelidae) eggs in Missouri: Effects of egg strains, infestation dates, and infestation levels on corn root damage. J. Kans. Entomol. Soc. 1999, 72, 214–221. [Google Scholar] [CrossRef]
- Hibbard, B.E.; Willmot, D.B.; Garcia, F.S.A.; Darrah, L.L. Registration of the maize germplasm CRW3(S1)C6 with resistance to western corn rootworm. J. Plant Regist. 2007, 1, 151–152. [Google Scholar] [CrossRef]
- Rogers, R.R.; Russell, W.A.; Owens, J.C. Evaluation of a vertical-pull technique in population improvement of maize for corn rootworm tolerance. Crop Sci. 1976, 16, 591–594. [Google Scholar] [CrossRef]
- Knutson, R.J.; Hibbard, B.E.; Barry, B.D.; Smith, V.A.; Darrah, L.L. Comparison of screening techniques for western corn rootworm (Coleoptera: Chrysomelidae) host-plant resistance. J. Econ. Entomol. 1999, 92, 714–722. [Google Scholar] [CrossRef]
- Abel, C.A.; Berhow, M.A.; Wilson, R.L.; Binder, B.F.; Hibbard, B.E. Evaluation of conventional resistance to European corn borer (Lepidoptera: Crambidae) and western corn rootworm (Coleoptera: Chrysomelidae) in experimental maize lines developed from a backcross breeding program. J. Econ. Entomol. 2000, 93, 1814–1821. [Google Scholar] [CrossRef]
- Garcia, F.S.A.; Dashiell, K.E.; Prischmann, D.A.; Bohn, M.O.; Hibbard, B.E. Conventional screening overlooks resistance sources: Rootworm damage of diverse inbred lines and their B73 hybrids is unrelated. J. Econ. Entomol. 2009, 102, 1317–1324. [Google Scholar] [CrossRef]
- El Khishen, A.A.; Bohn, M.O.; Voldseth, P.D.A.; Dashiell, K.E.; French, B.W.; Hibbard, B.E. Native resistance to western corn rootworm (Coleoptera: Chrysomelidae) larval feeding: Characterization and mechanisms. J. Econ. Entomol. 2009, 102, 2350–2359. [Google Scholar] [CrossRef]
- Bernklau, E.J.; Hibbard, B.E.; Bjostad, L.B. Antixenosis in maize reduces feeding by western corn rootworm larvae (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2010, 103, 2052–2060. [Google Scholar] [CrossRef]
- Šimić, D.; Ivezić, M.; Brkić, I.; Raspudić, E.; Brmež, M.; Majić, I.; Brkić, A.; Ledenčan, T.; Tollefson, J.J.; Hibbard, B.E. Environmental and genotypic effects for western corn rootworm tolerance traits in American and European maize trials. Maydica 2007, 52, 425–430. [Google Scholar]
- Ivezic, M.; Tollefson, J.J.; Raspudic, E.; Brkic, I.; Brmez, M.; Hibbard, B.E. Evaluation of corn hybrids for tolerance to corn rootworm (Diabrotica virgifera virgifera LeConte) larval feeding. Cereal Res. Commun. 2006, 34, 1101–1107. [Google Scholar] [CrossRef]
- Ivezić, M.; Tollefson, J.J.; Raspudić, E.; Hibbard, B.E.; Brkić, I. Evaluation of Croation corn hybrids for tolerance to corn rootworm (Diabrotica virgifera virgifera LeConte) larval feeding. In Proceedings of the 21st IWGO Conference and VIII Diabrotica subgroup meeting, Venice, Italy, 27 October–3 November 2001; pp. 205–212. [Google Scholar]
- Ivezić, M.; Raspudić, E.; Brmež, M.; Majić, I.; Brkić, I.; Tollefson, J.J.; Bohn, M.; Hibbard, B.E.; Šimić, D. A review of resistance breeding options targeting western corn rootworm (Diabrotica virgifera virgifera LeConte). Agric. For. Entomol. 2009, 11, 307–311. [Google Scholar] [CrossRef]
- Bohn, M. How to improve rootworm resistance in corn. In Proceedings of the 41st Illinois Corn Breeders School; Illinois Corn Breeders School: Champaign-Urbana, IL, USA, 2005; pp. 183–194. [Google Scholar]
- Rogers, R.R.; Russell, W.A.; Owens, J.C. Relationship of corn rootworm (Diabrotica) tolerance to yield in the Isss (Iowa stiff stalk synthetic) maize population. Iowa State J. Res. 1976, 51, 125–129. [Google Scholar]
- Bohn, M.O.; Marroquin, J.J.; Garcia, F.S.; Dashiell, K.; Willmot, D.B.; Hibbard, B.E. Quantitative trait loci mapping of western corn rootworm (Coleoptera: Chrysomelidae) host plant resistance in two populations of doubled haploid lines in maize (Zea mays L.). J. Econ. Entomol. 2018, 111, 435–444. [Google Scholar] [CrossRef]
- Russell, W.A.; Owens, J.C.; Peters, D.C.; Rogers, R.R. Registration of maize germplasm 1 (Reg. Nos. GP 72 and GP 73). Crop Sci. 1976, 16, 886–887. [Google Scholar] [CrossRef]
- Rogers, R.R.; Russell, W.A.; Owens, J.C. Expected gains from selection in maize for resistance to corn rootworms. Maydica 1977, 22, 27–36. [Google Scholar]
- Russell, W.A.; Penny, L.H.; Guthrie, W.D.; Dicke, F.F. Registration of maize germplasm inbreds 1 (Reg. Nos. GP 1 to 5). Crop Sci. 1971, 11, 140. [Google Scholar] [CrossRef]
- Russell, W.A.; Penny, L.H.; Sprague, G.F.; Guthrie, W.D.; Dicke, F.F. Registration of corn parental lines (Reg. nos. PL 1 to 13). Crop Sci. 1971, 11, 143. [Google Scholar] [CrossRef]
- Owens, J.C.; Peters, D.C.; Hallauer, A.R. Corn rootworm tolerance in maize. Environ. Entomol. 1974, 3, 767–772. [Google Scholar] [CrossRef]
- Branson, T.F.; Welch, V.A.; Sutter, G.R.; Fisher, J.R. Resistance to larvae of Diabrotica virgifera virgifera in three experimental maize hybrids. Environ. Entomol. 1983, 12, 1509–1512. [Google Scholar] [CrossRef]
- Brkić, A.; Šimić, D.; Jambrović, A.; Zdunić, Z.; Ledenĉan, T.; Raspudić, E.; Brmeţ, M.; Brkić, J.; Mazur, M.; Galić, V. QTL analysis of western corn rootworm resistance traits in maize ibm population grown in continuous maize. Genetika 2020, 52, 137–148. [Google Scholar] [CrossRef]
- Prasanna, B.M. Diversity in global maize germplasm: Characterization and utilization. J. Biosci. 2012, 37, 843–855. [Google Scholar] [CrossRef]
- Meihls, L.N.; Kaur, H.; Jander, G. Natural variation in maize defense against insect herbivores. Cold Spring Harb. Symp. Quant. Biol. 2012, 77, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Mohan, S.; Ma, P.W.K.; Pechan, T.; Bassford, E.R.; Williams, W.P.; Luthe, D.S. Degradation of the S. frugiperda peritrophic matrix by an inducible maize cysteine protease. J. Insect Physiol. 2006, 52, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Mohan, S.; Ma, P.W.K.; Williams, W.P.; Luthe, D.S. A naturally occurring plant cysteine protease possesses remarkable toxicity against insect pests and synergizes Bacillus thuringiensis toxin. PLoS ONE 2008, 3, e1786. [Google Scholar] [CrossRef]
- Duque, C.L.; Loades, K.W.; Tooker, J.F.; Brown, K.M.; Williams, P.W.; Luthe, D.S. A maize inbred exhibits resistance against western corn rootwoorm, Diabrotica virgifera virgifera. J. Chem. Ecol. 2017, 43, 1108–1123. [Google Scholar] [CrossRef]
- Ramstein, G.P.; Jensen, S.E.; Buckler, E.S. Breaking the curse of dimensionality to identify causal variants in Breeding 4. Theor. Appl. Genet. 2019, 132, 559–567. [Google Scholar] [CrossRef]
- Branson, T.F.; Ortman, E.E. Host range of larvae of the western corn rootworm. J. Econ. Entomol. 1967, 60, 201–203. [Google Scholar] [CrossRef]
- Branson, T.F.; Ortman, E.E. The host range of larvae of the western corn rootworm: Further studies. J. Econ. Entomol. 1970, 63, 800–803. [Google Scholar] [CrossRef]
- Clark, T.L.; Hibbard, B.E. Comparison of nonmaize hosts to support western corn rootworm (Coleoptera: Chrysomelidae) larval biology. Environ. Entomol. 2004, 33, 681–689. [Google Scholar] [CrossRef]
- Oyediran, I.O.; Hibbard, B.E.; Clark, T.L. Prairie grasses as hosts of the western corn rootworm (Coleoptera: Chrysomelidae). Environ. Entomol. 2004, 33, 740–747. [Google Scholar] [CrossRef]
- Wilson, T.A.; Hibbard, B.E. Host suitability of nonmaize agroecosystem grasses for the western corn rootworm (Coleoptera: Chrysomelidae). Environ. Entomol. 2004, 33, 1102–1108. [Google Scholar] [CrossRef]
- Moeser, J.; Vidal, S. How to measure the food utilization of subterranean insects: A case study with the western corn rootworm (Diabrotica virgifera virgifera). J. Appl. Entomol. 2005, 129, 60–63. [Google Scholar] [CrossRef]
- Hibbard, B.E.; Schweikert, Y.M.; Higdon, M.L.; Ellersieck, M.R. Maize phenology affects establishment, damage, and development of the western corn rootworm (Coleoptera: Chrysomelidae). Environ. Entomol. 2008, 37, 1558–1564. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, W.S.; Hanway, J.J.; Benson, G.O. How a Corn Plant Develops; Iowa State University: Ames, IA, USA, 1992. [Google Scholar]
- Law, J.H. Insects, oxygen, and iron. Biochem. Biophys. Res. Commun. 2002, 292, 1191–1195. [Google Scholar] [CrossRef]
- Chege, P.G.; Clark, T.L.; Hibbard, B.E. Alternate host phenology affects survivorship, growth, and development of western corn rootworm (Coleoptera: Chrysomelidae) larvae. Environ. Entomol. 2005, 34, 1441–1447. [Google Scholar] [CrossRef]
- Huynh, M.P.; Hibbard, B.E.; Vella, M.; Lapointe, S.L.; Niedz, R.P.; Shelby, K.S.; Coudron, T.A. Development of an improved and accessible diet for western corn rootworm larvae using response surface modeling. Sci. Rep. 2019, 9, 6009. [Google Scholar] [CrossRef] [PubMed]
- Hannon, G.J. RNA interference. Nature 2002, 418, 244–251. [Google Scholar] [CrossRef]
- Katoch, R.; Sethi, A.; Thakur, N.; Murdock, L.L. RNAi for insect control: Current perspective and future challenges. Appl. Biochem. Biotechnol. 2013, 171, 847–873. [Google Scholar] [CrossRef]
- Ivashuta, S.; Zhang, Y.; Wiggins, B.E.; Ramaseshadri, P.; Segers, G.C.; Johnson, S.; Meyer, S.E.; Kerstetter, R.A.; McNulty, B.C.; Bolognesi, R.; et al. Environmental RNAi in herbivorous insects. RNA 2015, 21, 840–850. [Google Scholar] [CrossRef]
- Turner, C.T.; Davy, M.W.; Diarmid, M.R.M.; Plummer, K.M.; Birch, N.P.; Newcomb, R.D. RNA interference in the light brown apple moth, Epiphyas postvittana (Walker) induced by double-stranded RNA feeding. Insect Mol. Biol. 2006, 15, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, R.; Sivakumar, S.; Agrawal, N.; Malhotra, P.; Bhatnagar, R.K. Silencing of midgut aminopeptidase N of Spodoptera litura by double-stranded RNA establishes its role as Bacillus thuringiensis toxin receptor. J. Biol. Chem. 2002, 6, 46849–46851. [Google Scholar] [CrossRef]
- Bucher, G.; Scholten, J.; Klingler, M. Parental RNAi in tribolium (Coleoptera). Curr. Biol. 2002, 5, R85–R86. [Google Scholar] [CrossRef]
- Tomoyasu, Y.; Denell, R.E. Larval RNAi in Tribolium (Coleoptera) for analyzing adult development. Dev. Genes Evol. 2004, 214, 575–578. [Google Scholar] [CrossRef]
- Soares, C.A.G.; Lima, C.M.R.; Dolan, M.C.; Piesman, J.; Beard, C.B.; Zeidner, N.S. Capillary feeding of specific dsRNA induces silencing of the isac gene in nymphal Ixodes scapularis ticks. Insect Mol. Biol. 2005, 14, 443–452. [Google Scholar] [CrossRef]
- Baum, J.A.; Bogaert, T.; Clinton, W.; Heck, G.R.; Feldmann, P.; Ilagan, O.; Johnson, S.; Plaetinck, G.; Munyikwa, T.; Pleau, M.; et al. Control of coleopteran insect pests through RNA interference. Nat. Biotechnol. 2007, 25, 1322–1326. [Google Scholar] [CrossRef]
- Bolognesi, R.; Ramaseshadri, P.; Anderson, J.; Bachman, P.; Clinton, W.; Flannagan, R.; Ilagan, O.; Lawrence, C.; Levine, S.; Moar, W.; et al. Characterizing the mechanism of action of double-stranded RNA activity against western corn rootworm (Diabrotica virgifera virgifera LeConte). PLoS ONE 2012, 7, e47534. [Google Scholar] [CrossRef]
- Rangasamy, M.; Siegfried, B.D. Validation of RNA interference in western corn rootworm Diabrotica virgifera virgifera LeConte (Coleoptera: Chrysomelidae) adults. Pest Manag. Sci. 2012, 68, 587–591. [Google Scholar] [CrossRef] [PubMed]
- Vélez, A.M.; Fishilevich, E.; Rangasamy, M.; Khajuria, C.; McCaskill, D.G.; Pereira, A.E.; Gandra, P.; Frey, M.L.F.; Worden, S.E.; Whitlock, S.L.; et al. Control of western corn rootworm via RNAi traits in maize: Lethal and sublethal effects of Sec23 dsRNA. Pest Manag. Sci. 2020, 76, 1500–1512. [Google Scholar] [CrossRef] [PubMed]
- Khajuria, C.; Vélez, A.M.; Rangasamy, M.; Wang, H.; Fishilevich, E.; Frey, M.L.F.; Carneiro, N.P.; Gandra, P.; Narva, K.E.; Siegfried, B.D. Parental RNA interference of genes involved in embryonic development of the western corn rootworm, Diabrotica virgifera virgifera LeConte. Insect Biochem. Mol. Biol. 2015, 62, 54–62. [Google Scholar] [CrossRef]
- Vélez, A.M.; Fishilevich, E.; Matz, N.; Storer, N.P.; Narva, K.E.; Siegfried, B.D. Parameters for successful parental RNAi as an insect pest management tool in western corn rootworm, Diabrotica virgifera virgifera. Genes 2017, 8, 7. [Google Scholar] [CrossRef]
- Ramaseshadri, P.; Segers, G.; Flannagan, R.; Wiggins, E.; Clinton, W.; Ilagan, O.; McNulty, B.; Clark, T.; Bolognesi, R. Physiological and cellular responses caused by RNAi-mediated suppression of Snf7 orthologue in western corn rootworm (Diabrotica virgifera virgifera) larvae. PLoS ONE 2013, 8, e54270. [Google Scholar] [CrossRef]
- Hu, X.; Richtman, N.M.; Zhao, J.Z.; Duncan, K.E.; Niu, X.; Procyk, L.A.; Oneal, M.A.; Kernodle, B.M.; Steimel, J.P.; Crane, V.C.; et al. Discovery of midgut genes for the RNA interference control of corn rootworm. Sci. Rep. 2016, 6, 542. [Google Scholar] [CrossRef]
- Li, H.; Khajuria, C.; Rangasamy, M.; Gandra, P.; Fitter, M.; Geng, C.; Woosely, A.; Hasler, J.; Schulenberg, G.; Worden, S.; et al. Long dsRNA but not siRNA initiates RNAi in western corn rootworm larvae and adults. J. Appl. Entomol. 2015, 139, 432–445. [Google Scholar] [CrossRef]
- Niu, X.; Kassa, A.; Hu, X.; Robeson, J.; McMahon, M.; Richtman, N.M.; Steimel, J.P.; Kernodle, B.M.; Crane, V.C.; Sandahl, G.; et al. Control of western corn rootworm (Diabrotica virgifera virgifera) reproduction through plant-mediated RNA interference. Sci. Rep. 2017, 7, 12591. [Google Scholar] [CrossRef] [PubMed]
- Head, G.P.; Carroll, M.W.; Evans, S.P.; Rule, D.M.; Willse, A.R.; Clark, T.L.; Storer, N.P.; Flannagan, R.D.; Samuel, L.W.; Meinke, L.J. Evaluation of SmartStax and SmartStax PRO maize against western corn rootworm and northern corn rootworm: Efficacy and resistance management. Pest Manag. Sci. 2017, 73, 1883–1899. [Google Scholar] [CrossRef]
- Anderson, J.A.; Mickelson, J.; Challender, M.; Moellring, E.; Sult, T.; TeRonde, S.; Walker, C.; Wang, Y.; Maxwell, C.A. Agronomic and compositional assessment of genetically modified DP23211 maize for corn rootworm control. GM Crop. Food 2020, 11, 206–214. [Google Scholar] [CrossRef]
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setälä, H.; Van Der Putten, W.H.; Wall, D.H. Ecological linkages between aboveground and belowground biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef] [PubMed]
- Blundell, R.; Schmidt, J.E.; Igwe, A.; Cheung, A.L.; Vannette, R.L.; Gaudin, A.C.M.; Casteel, C.L. Organic management promotes natural pest control through altered plant resistance to insects. Nat. Plants 2020, 6, 483–491. [Google Scholar] [CrossRef]
- Mueller, U.G.; Sachs, J.L. Engineering microbiomes to improve plant and animal health. Trends Microbiol. 2015, 23, 606–617. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Robert, C.A.M.; Cadot, S.; Zhang, X.; Ye, M.; Li, B.; Manzo, D.; Chervet, N.; Steinger, T.; Van Der Heijden, M.G.A.; et al. Root exudate metabolites drive plant-soil feedbacks on growth and defense by shaping the rhizosphere microbiota. Nat. Commun. 2018, 9, 2738. [Google Scholar] [CrossRef] [PubMed]
- Willis, A.; Rodrigues, B.F.; Harris, P.J.C. The ecology of arbuscular mycorrhizal fungi. CRC Crit. Rev. Plant Sci. 2013, 32, 1–20. [Google Scholar] [CrossRef]
- Javaid, A. Arbuscular mycorrhizal mediated nutrition in plants. J. Plant Nutr. 2009. [Google Scholar] [CrossRef]
- Murrell, E.G.; Ray, S.; Lemmon, M.E.; Luthe, D.S.; Kaye, J.P. Cover crop species affect mycorrhizae-mediated nutrient uptake and pest resistance in maize. Renew. Agric. Food Syst. 2020, 35, 467–474. [Google Scholar] [CrossRef]
- Gehring, C.; Bennett, A. Mycorrhizal fungal-plant-insect interactions: The importance of a community approach. Environ. Entomol. 2009, 38, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Koricheva, J.; Gange, A.C.; Jones, T. Effects of mycorrhizal fungi on insect herbivores: A meta-analysis. Ecology 2009, 90, 2088–2097. [Google Scholar] [CrossRef] [PubMed]
- Bennett, A.E.; Garcia, A.J.; Bever, J.D. Three-way interactions among mutualistic mycorrhizal fungi, plants, and plant enemies: Hypotheses and synthesis. Am. Nat. 2006, 167, 141–152. [Google Scholar] [CrossRef]
- Jaffuel, G.; Imperiali, N.; Shelby, K.; Herrera, C.R.; Geisert, R.; Maurhofer, M.; Loper, J.; Keel, C.; Turlings, T.C.J.; Hibbard, B.E. Protecting maize from rootworm damage with the combined application of arbuscular mycorrhizal fungi, Pseudomonas bacteria and entomopathogenic nematodes. Sci. Rep. 2019, 9, 3127. [Google Scholar] [CrossRef]
- Vukicevich, E.; Lowery, T.; Bowen, P.; Torres, Ú.J.R.; Hart, M. Cover crops to increase soil microbial diversity and mitigate decline in perennial agriculture. A review. Agron. Sustain. Dev. 2016, 36, 48. [Google Scholar] [CrossRef]
- Hartwig, N.L.; Ammon, H.U. Cover crops and living mulches. Weed Sci. 2002, 50, 688–699. [Google Scholar] [CrossRef]
- Bainard, L.D.; Bainard, J.D.; Hamel, C.; Gan, Y. Spatial and temporal structuring of arbuscular mycorrhizal communities is differentially influenced by abiotic factors and host crop in a semi-arid prairie agroecosystem. FEMS Microbiol. Ecol. 2014, 88, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Benitez, M.S.; Taheri, W.I.; Lehman, R.M. Selection of fungi by candidate cover crops. Appl. Soil Ecol. 2016, 103, 72–82. [Google Scholar] [CrossRef]
- Hontoria, C.; González, G.I.; Quemada, M.; Roldán, A.; Alguacil, M.M. The cover crop determines the AMF community composition in soil and in roots of maize after a ten-year continuous crop rotation. Sci. Total Environ. 2019, 660, 913–922. [Google Scholar] [CrossRef]
- Cloutier, M.L.; Murrell, E.; Barbercheck, M.; Kaye, J.; Finney, D.; González, G.I.; Bruns, M.A. Fungal community shifts in soils with varied cover crop treatments and edaphic properties. Sci. Rep. 2020, 10, 6198. [Google Scholar] [CrossRef] [PubMed]
- Lundgren, J.G.; Fergen, J.K. The effects of a winter cover crop on Diabrotica virgifera (Coleoptera: Chrysomelidae) populations and beneficial arthropod communities in no-till maize. Environ. Entomol. 2010, 39, 1816–1828. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Douglas, A.E. Symbiotic microorganisms: Untapped resources for insect pest control. Trends Biotechnol. 2007, 25, 338–342. [Google Scholar] [CrossRef]
- Dematheis, F.; Kurtz, B.; Vidal, S.; Smalla, K. Microbial communities associated with the larval gut and eggs of the western corn rootworm. PLoS ONE 2012, 7, 4685. [Google Scholar] [CrossRef]
- Ludwick, D.C.; Ericsson, A.C.; Meihls, L.N.; Gregory, M.L.J.; Finke, D.L.; Coudron, T.A.; Hibbard, B.E.; Shelby, K.S. Survey of bacteria associated with western corn rootworm life stages reveals no difference between insects reared in different soils. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.C.; Spencer, J.L.; Curzi, M.J.; Zavala, J.A.; Seufferheld, M.J. Gut bacteria facilitate adaptation to crop rotation in the western corn rootworm. Proc. Natl. Acad. Sci. USA 2013, 110, 11917–11922. [Google Scholar] [CrossRef] [PubMed]
- Perlatti, B.; Luiz, A.L.; Prieto, E.L.; Fernandes, J.B.; da Silva, M.F.d.G.F.; Ferreira, D.; Costa, E.N.; Júnior, B.A.L.; Forim, M.R. MALDI-TOF MS identification of microbiota associated with pest insect Diabrotica speciosa. Agric. For. Entomol. 2017, 19, 408–417. [Google Scholar] [CrossRef]
- Robert, C.A.M.; Frank, D.L.; Leach, K.A.; Turlings, T.C.J.; Hibbard, B.E.; Erb, M. Direct and indirect plant defenses are not suppressed by endosymbionts of a specialist root herbivore. J. Chem. Ecol. 2013, 39, 507–515. [Google Scholar] [CrossRef]
- Clark, T.L.; Meinke, L.J.; Skoda, S.R.; Foster, J.E. Occurrence of Wolbachia in selected Diabroticite (Coleoptera: Chrysomelidae) beetles. Ann. Entomol. Soc. Am. 2001, 94, 877–885. [Google Scholar] [CrossRef][Green Version]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master manipulators of invertebrate biology. Nat. Rev. Microbiol. 2008, 6, 741–751. [Google Scholar] [CrossRef]
- Werren, J.H. Wolbachia and Speciation. In Endless Forms: Species and Speciation; Berlocher, S.H., Ed.; Oxford University Press: New York, NY, USA, 1998; pp. 245–260. [Google Scholar]
- Bordenstein, S.R. Symbiosis and the origin of species. In Insect Symbiosis; CRC Press: Boca Raton, FL, USA, 2003; pp. 283–304. [Google Scholar]
- Giordano, R.; Jackson, J.J.; Robertson, H.M. The role of Wolbachia bacteria in reproductive incompatibilities and hybrid zones of Diabrotica beetles and Gryllus crickets. Proc. Natl. Acad. Sci. USA 1997, 94, 11439–11444. [Google Scholar] [CrossRef]
- Lombaert, E.; Ciosi, M.; Miller, N.J.; Sappington, T.W.; Blin, A.; Guillemaud, T. Colonization history of the western corn rootworm (Diabrotica virgifera virgifera) in North America: Insights from random forest ABC using microsatellite data. Biol. Invasions 2018, 20, 665–677. [Google Scholar] [CrossRef]
- Ye, Y.H.; Seleznev, A.; Flores, H.A.; Woolfit, M.; McGraw, E.A. Gut microbiota in Drosophila melanogaster interacts with Wolbachia but does not contribute to Wolbachia-mediated antiviral protection. J. Invertebr. Pathol. 2017, 143, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Hedges, L.M.; Brownlie, J.C.; O’Neill, S.L.; Johnson, K.N. Wolbachia and virus protection in insects. Science 2008, 322, 702. [Google Scholar] [CrossRef]
- Barr, K.L.; Hearne, L.B.; Briesacher, S.; Clark, T.L.; Davis, G.E. Microbial symbionts in insects influence down-regulation of defense genes in maize. PLoS ONE 2010, 5, e11339. [Google Scholar] [CrossRef] [PubMed]
- Lance, D.R. Odors influence choice of oviposition sites by Diabrotica virgifera virgifera (Coleoptera: Chrysomelidae). J. Chem. Ecol. 1992, 18, 1227–1237. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.P.; Beran, F. Gut microbiota degrades toxic isothiocyanates in a flea beetle pest. Mol. Ecol. 2020, 29, 4692–4705. [Google Scholar] [CrossRef]
- Itoh, H.; Tago, K.; Hayatsu, M.; Kikuchi, Y. Detoxifying symbiosis: Microbe-mediated detoxification of phytotoxins and pesticides in insects. Nat. Prod. Rep. 2018, 35, 434–454. [Google Scholar] [CrossRef]
- van den Bosch, T.J.M.; Welte, C.U. Detoxifying symbionts in agriculturally important pest insects. Microb. Biotechnol. 2017, 10, 531–540. [Google Scholar] [CrossRef]
- Hammer, T.J.; Bowers, M.D. Gut microbes may facilitate insect herbivory of chemically defended plants. Oecologia 2015, 179, 1–14. [Google Scholar] [CrossRef]
- Steinhaus, E. Microbial Control-The emergence of an idea. Hilgardia 1956, 26, 107–160. [Google Scholar] [CrossRef][Green Version]
- Lacey, L.A.; Grzywacz, D.; Ilan, S.D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef] [PubMed]
- Vega, F.; Kaya, H. (Eds.) Insect Pathology; Academic Press: San Diego, CA, USA, 2012; ISBN 9780123849847. [Google Scholar]
- Knowles, B.H.; Ellar, D.J. Colloid-osmotic lysis is a general feature of the mechanism of action of Bacillus thuringiensis δ-endotoxins with different insect specificity. BBA Gen. Subj. 1987, 924, 509–518. [Google Scholar] [CrossRef]
- Yin, Y.; Flasinski, S.; Moar, W.; Bowen, D.; Chay, C.; Milligan, J.; Kouadio, J.L.; Pan, A.; Werner, B.; Buckman, K.; et al. A new Bacillus thuringiensis protein for western corn rootworm control. PLoS ONE 2020, 15, e0242791. [Google Scholar] [CrossRef]
- Bowen, D.; Yin, Y.; Flasinski, S.; Chay, C.; Bean, G.; Milligan, J.; Moar, W.; Pan, A.; Werner, B.; Buckman, K.; et al. Cry75Aa (Mpp75Aa) insecticidal proteins for controlling the western corn rootworm, Diabrotica virgifera virgifera, (Coleoptera: Chrysomelidae), isolated from the insect pathogenic bacteria Brevibacillus laterosporus. Appl. Environ. Microbiol. 2020. [Google Scholar] [CrossRef]
- Sampson, K.; Zaitseva, J.; Stauffer, M.; Berg, V.B.; Guo, R.; Tomso, D.; McNulty, B.; Desai, N.; Balasubramanian, D. Discovery of a novel insecticidal protein from Chromobacterium piscinae, with activity against western corn rootworm, Diabrotica virgifera virgifera. J. Invertebr. Pathol. 2017, 142, 34–43. [Google Scholar] [CrossRef]
- Wei, J.Z.; Rear, O.J.; Schellenberger, U.; Rosen, B.A.; Park, Y.J.; McDonald, M.J.; Zhu, G.; Xie, W.; Kassa, A.; Procyk, L.; et al. A selective insecticidal protein from Pseudomonas mosselii for corn rootworm control. Plant Biotechnol. J. 2018, 16, 649–659. [Google Scholar] [CrossRef]
- Yalpani, N.; Altier, D.; Barry, J.; Kassa, A.; Nowatzki, T.M.; Sethi, A.; Zhao, J.Z.; Diehn, S.; Crane, V.; Sandahl, G.; et al. An Alcaligenes strain emulates Bacillus thuringiensis producing a binary protein that kills corn rootworm through a mechanism similar to Cry34Ab1/Cry35Ab1. Sci. Rep. 2017, 7, 3063. [Google Scholar] [CrossRef]
- Bowling, A.J.; Pence, H.E.; Li, H.; Tan, S.Y.; Evans, S.L.; Narva, K.E. Histopathological effects of Bt and TcdA insecticidal proteins on the midgut epithelium of western corn rootworm larvae (Diabrotica virgifera virgifera). Toxins (Basel) 2017, 9, 156. [Google Scholar] [CrossRef] [PubMed]
- Panevska, A.; Hodnik, V.; Skočaj, M.; Novak, M.; Modic, Š.; Pavlic, I.; Podržaj, S.; Zarić, M.; Resnik, N.; Maček, P.; et al. Pore-forming protein complexes from Pleurotus mushrooms kill western corn rootworm and Colorado potato beetle through targeting membrane ceramide phosphoethanolamine. Sci. Rep. 2019, 9, 5073. [Google Scholar] [CrossRef]
- Hurst, M.R.H.; van Koten, C.; Jackson, T.A. Pathology of Yersinia entomophaga MH96 towards Costelytra zealandica (Coleoptera; Scarabaeidae) larvae. J. Invertebr. Pathol. 2014, 115, 102–107. [Google Scholar] [CrossRef]
- Ruiu, L.; Satta, A.; Floris, I. Emerging entomopathogenic bacteria for insect pest management. Bull. Insectology 2013, 66, 181–186. [Google Scholar]
- Jackson, T.A.; Pearson, J.F.; Callaghan, O.M.; Mahanty, H.K.; Willocks, M.J. Pathogen to product - development of Serratia entomophila (Enterobacteriaceae) as a commercial biological control agent for the New Zealand grass grub (Costelytra zealandica). In Use of Pathogens in Scarab Management; Jackson, T.A., Glare, T.R., Eds.; Intercept Ltd.: Andover, MA, USA, 1992; pp. 191–198. [Google Scholar]
- Liu, S.; Chen, Y.; Sappington, T.W.; Bonning, B.C. Genome sequence of a novel positive-sense, single-stranded RNA virus isolated from western corn rootworm, Diabrotica virgifera virgifera LeConte. Genome Announc. 2017, 5, 17. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Chen, Y.; Sappington, T.W.; Bonning, B.C. Genome sequence of the first coleopteran iflavirus isolated from western corn rootworm, Diabrotica virgifera virgifera LeConte. Genome Announc. 2017, 5, 16. [Google Scholar] [CrossRef]
- Liu, S.; Chen, Y.; Sappington, T.W.; Bonning, B.C. Genome sequence of Diabrotica virgifera virgifera virus 2, a novel small RNA virus of the western corn rootworm, Diabrotica virgifera virgifera LeConte. Genome Announc. 2017, 5, 17. [Google Scholar] [CrossRef] [PubMed]
- Huger, A.M. The Oryctes virus: Its detection, identification, and implementation in biological control of the coconut palm rhinoceros beetle, Oryctes rhinoceros (Coleoptera: Scarabaeidae). J Invertebr. Pathol. 2005, 89, 78–84. [Google Scholar] [CrossRef]
- Pilz, C.; Wegensteiner, R.; Keller, S. Selection of entomopathogenic fungi for the control of the western corn rootworm Diabrotica virgifera virgifera. J. Appl. Entomol. 2007, 131, 426–431. [Google Scholar] [CrossRef]
- Pilz, C.; Keller, S.; Kuhlmann, U.; Toepfer, S. Comparative efficacy assessment of fungi, nematodes and insecticides to control western corn rootworm larvae in maize. BioControl 2009, 54, 671–684. [Google Scholar] [CrossRef]
- Meyling, N.V.; Eilenberg, J. Ecology of the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae in temperate agroecosystems: Potential for conservation biological control. Biol. Control 2007, 43, 145–155. [Google Scholar] [CrossRef]
- Toepfer, S.; Gueldenzoph, C.; Ehlers, R.U.; Kuhlmann, U. Screening of entomopathogenic nematodes for virulence against the invasive western corn rootworm, Diabrotica virgifera virgifera (Coleoptera: Chrysomelidae) in Europe. Bull. Entomol. Res. 2005, 95, 473–482. [Google Scholar] [CrossRef]
- Kurtz, B.; Toepfer, S.; Ehlers, R.U.; Kuhlmann, U. Assessment of establishment and persistence of entomopathogenic nematodes for biological control of western corn rootworm. J. Appl. Entomol. 2007, 131, 420–425. [Google Scholar] [CrossRef]
- Kurtz, B.; Hiltpold, I.; Turlings, T.C.J.; Kuhlmann, U.; Toepfer, S. Comparative susceptibility of larval instars and pupae of the western corn rootworm to infection by three entomopathogenic nematodes. BioControl 2009, 54, 255. [Google Scholar] [CrossRef]
- Ilan, S.D.I.; Gouge, D.H.; Piggott, S.J.; Fife, J.P. Application technology and environmental considerations for use of entomopathogenic nematodes in biological control. Biol. Control 2006, 38, 124–133. [Google Scholar] [CrossRef]
- Hiltpold, I.; Jaffuel, G.; Turlings, T.C.J. The dual effects of root-cap exudates on nematodes: From quiescence in plant-parasitic nematodes to frenzy in entomopathogenic nematodes. J. Exp. Bot. 2015, 66, 603–611. [Google Scholar] [CrossRef]
- Jaffuel, G.; Hiltpold, I.; Turlings, T.C.J. Highly potent extracts from pea (Pisum sativum) and maize (Zea mays) roots can be used to induce quiescence in entomopathogenic nematodes. J. Chem. Ecol. 2015, 41, 793–800. [Google Scholar] [CrossRef]
- Shields, E.J.; Testa, A.M.; Neil, O.W.J. Long-term Persistence of Native New York Entomopathogenic Nematode Isolates Across Crop Rotation. J. Econ. Entomol. 2018, 111, 2592–2598. [Google Scholar] [CrossRef]
- Ehlers Ralf, U.; Hiltpold, I.; Kulhmann, U.; Toepfer, S. Field results on the use of Heterorhabditis bacteriophage against the invasive maize pest Diabrotica virgifera virgeifera. Insect Pathog. Insect Parasit Nematodes 2008, 31, 332–335. [Google Scholar]
- Toepfer, S.; Peters, A.; Ehlers, R.U.; Kuhlmann, U. Comparative assessment of the efficacy of entomopathogenic nematode species at reducing western corn rootworm larvae and root damage in maize. J. Appl. Entomol. 2008, 132, 337–348. [Google Scholar] [CrossRef]
- Jaffuel, G.; Sbaiti, I.; Turlings, T.C.J. Encapsulated entomopathogenic nematodes can protect maize plants from Diabrotica balteata larvae. Insects 2020, 11, 27. [Google Scholar] [CrossRef]
- Griffin, C.T. Perspectives on the behavior of entomopathogenic nematodes from dispersal to reproduction: Traits contributing to nematode fitness and biocontrol efficacy. J Nematol. 2012, 44, 177–184. [Google Scholar]
- Dillman, A.R.; Guillermin, M.L.; Lee, J.H.; Kim, B.; Sternberg, P.W.; Hallem, E.A. Olfaction shapes host-parasite interactions in parasitic nematodes. Proc. Natl. Acad. Sci. USA 2012, 109, E2324–E2333. [Google Scholar] [CrossRef] [PubMed]
- Rasmann, S.; Köllner, T.G.; Degenhardt, J.; Hiltpold, I.; Toepfer, S.; Kuhlmann, U.; Gershenzon, J.; Turlings, T.C.J. Recruitment of entomopathogenic nematodes by insect-damaged maize roots. Nature 2005, 434, 732–737. [Google Scholar] [CrossRef] [PubMed]
- Anbesse, S.; Ehlers, R.U. Attraction of Heterorhabditis sp. toward synthetic (E)-beta-cariophyllene, a plant SOS signal emitted by maize on feeding by larvae of Diabrotica virgifera virgifera. Commun. Agric. Appl. Biol. Sci. 2010, 75, 455–458. [Google Scholar]
- Hiltpold, I.; Baroni, M.; Toepfer, S.; Kuhlmann, U.; Turlings, T.C.J. Selection of entomopathogenic nematodes for enhanced responsiveness to a volatile root signal helps to control a major root pest. J. Exp. Biol. 2010, 213, 2417–2423. [Google Scholar] [CrossRef]
- Zhang, X.; Machado, R.A.R.; Van Doan, C.; Arce, C.C.M.; Hu, L.; Robert, C.A.M. Entomopathogenic nematodes increase predation success by inducing cadaver volatiles that attract healthy herbivores. eLife 2019, 8, 6668. [Google Scholar] [CrossRef] [PubMed]
- Baiocchi, T.; Lee, G.; Choe, D.H.; Dillman, A.R. Host seeking parasitic nematodes use specific odors to assess host resources. Sci. Rep. 2017, 7, 6270. [Google Scholar] [CrossRef]
- Jagdale, G.B.; Kamoun, S.; Grewal, P.S. Entomopathogenic nematodes induce components of systemic resistance in plants: Biochemical and molecular evidence. Biol. Control 2009, 51, 102–109. [Google Scholar] [CrossRef]
- Helms, A.M.; Ray, S.; Matulis, N.L.; Kuzemchak, M.C.; Grisales, W.; Tooker, J.F.; Ali, J.G. Chemical cues linked to risk: Cues from below-ground natural enemies enhance plant defences and influence herbivore behaviour and performance. Funct. Ecol. 2019, 33, 798–808. [Google Scholar] [CrossRef]
- Robert, C.A.M.; Zhang, X.; Machado, R.A.R.; Schirmer, S.; Lori, M.; Mateo, P.; Erb, M.; Gershenzon, J. Sequestration and activation of plant toxins protect the western corn rootworm from enemies at multiple trophic levels. eLife 2017, 6, 9307. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Van Doan, C.; Arce, C.C.M.; Hu, L.; Gruenig, S.; Parisod, C.; Hibbard, B.E.; Hervé, M.R.; Nielson, C.; Robert, C.A.M.; et al. Plant defense resistance in natural enemies of a specialist insect herbivore. Proc. Natl. Acad. Sci. USA 2019, 116, 23174–23181. [Google Scholar] [CrossRef] [PubMed]
- Bruno, P.; Machado, R.A.R.; Glauser, G.; Köhler, A.; Herrera, C.R.; Bernal, J.; Toepfer, S.; Erb, M.; Robert, C.A.M.; Arce, C.C.M.; et al. Entomopathogenic nematodes from Mexico that can overcome the resistance mechanisms of the western corn rootworm. Sci. Rep. 2020, 10, 8257. [Google Scholar] [CrossRef] [PubMed]
- Machado, R.A.R.; Thönen, L.; Arce, C.C.M.; Theepan, V.; Prada, F.; Wüthrich, D.; Robert, C.A.M.; Vogiatzaki, E.; Shi, Y.M.; Schaeren, O.P.; et al. Engineering bacterial symbionts of nematodes improves their biocontrol potential to counter the western corn rootworm. Nat. Biotechnol. 2020, 38, 600–608. [Google Scholar] [CrossRef]
- Degenhardt, J.; Hiltpold, I.; Köllner, T.G.; Frey, M.; Gierl, A.; Gershenzon, J.; Hibbard, B.E.; Ellersieck, M.R.; Turlings, T.C.J. Restoring a maize root signal that attracts insect-killing nematodes to control a major pest. Proc. Natl. Acad. Sci. USA 2009, 106, 13213–13218. [Google Scholar] [CrossRef]
- Robert, C.A.M.; Erb, M.; Hiltpold, I.; Hibbard, B.E.; Gaillard, M.D.P.; Bilat, J.; Degenhardt, J.; Cambet, P.J.X.; Turlings, T.C.J.; Zwahlen, C. Genetically engineered maize plants reveal distinct costs and benefits of constitutive volatile emissions in the field. Plant Biotechnol. J. 2013, 11, 628–639. [Google Scholar] [CrossRef] [PubMed]
- Lewis, E.E.; Ilan, S.D.I. Host cadavers protect entomopathogenic nematodes during freezing. J. Invertebr. Pathol. 2002, 81, 25–32. [Google Scholar] [CrossRef]
- Ilan, S.D.I.; Brown, I. Earthworms as phoretic hosts for Steinernema carpocapsae and Beauveria bassiana: Implications for enhanced biological control. Biol. Control 2013, 66, 41–48. [Google Scholar] [CrossRef]
- Ilan, S.D.I.; Lewis, E.E.; Son, Y.; Tedders, W.L. Superior efficacy observed in entomopathogenic nematodes applied in infected-host cadavers compared with application in aqueous suspension. J. Invertebr. Pathol. 2003, 83, 270–272. [Google Scholar] [CrossRef]
- Kaya, H.K.; Nelsen, C.E. Encapsulation of Steinernematid and Heterorhabditid nematodes with calcium alginate: A new approach for insect control and other applications. Environ. Entomol. 1985, 14, 572–574. [Google Scholar] [CrossRef]
- Hiltpold, I.; Hibbard, B.E.; French, B.W.; Turlings, T.C.J. Capsules containing entomopathogenic nematodes as a Trojan horse approach to control the western corn rootworm. Plant Soil 2012, 358, 11–25. [Google Scholar] [CrossRef]
- Wu, S.; Kaplan, F.; Lewis, E.; Alborn, H.T.; Ilan, S.D.I. Infected host macerate enhances entomopathogenic nematode movement towards hosts and infectivity in a soil profile. J. Invertebr. Pathol. 2018, 159, 141–144. [Google Scholar] [CrossRef]
- Ilan, S.D.I.; Kaplan, F.; Hofman, O.C.; Schliekelman, P.; Alborn, H.T.; Lewis, E.E. Conspecific pheromone extracts enhance entomopathogenic infectivity. J. Nematol. 2019, 51, 1–5. [Google Scholar] [CrossRef]
- Hofman, O.C.; Kaplan, F.; Stevens, G.; Lewis, E.; Wu, S.; Alborn, H.T.; Gentil, P.A.; Ilan, S.D.I. Pheromone extracts act as boosters for entomopathogenic nematodes efficacy. J. Invertebr. Pathol. 2019, 164, 38–42. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Years | Location | References |
|---|---|---|
| 1935–1945 | Illinois Natural History Survey | [61,62] |
| 1970–2007 | Iowa State University | [63,64,73,84,95,106,108,109,110,111,112] |
| 1963–2010 | USDA-ARS, Brookings, South Dakota | [65,66,67,68,69,70,71,72,74,75,76,77,78,79,80,81,113] |
| 1990–1997 | University of Ottawa | [82,83,85,86,87,88,89] |
| 1992–present | USDA-ARS, Columbia, MO | [90,91,92,93,94,96,97,98,99,100] |
| 1995–present | University of J. J. Strossmayer | [101,102,103,104,114] |
| 2002–present | University of Illinois-Champaign | [105,107] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paddock, K.J.; Robert, C.A.M.; Erb, M.; Hibbard, B.E. Western Corn Rootworm, Plant and Microbe Interactions: A Review and Prospects for New Management Tools. Insects 2021, 12, 171. https://doi.org/10.3390/insects12020171
Paddock KJ, Robert CAM, Erb M, Hibbard BE. Western Corn Rootworm, Plant and Microbe Interactions: A Review and Prospects for New Management Tools. Insects. 2021; 12(2):171. https://doi.org/10.3390/insects12020171
Chicago/Turabian StylePaddock, Kyle J., Christelle A. M. Robert, Matthias Erb, and Bruce E. Hibbard. 2021. "Western Corn Rootworm, Plant and Microbe Interactions: A Review and Prospects for New Management Tools" Insects 12, no. 2: 171. https://doi.org/10.3390/insects12020171
APA StylePaddock, K. J., Robert, C. A. M., Erb, M., & Hibbard, B. E. (2021). Western Corn Rootworm, Plant and Microbe Interactions: A Review and Prospects for New Management Tools. Insects, 12(2), 171. https://doi.org/10.3390/insects12020171