
Mulches Used in Highbush Blueberry and Entomopathogenic Nematodes Affect Mortality Rates of Third-Instar Popillia japonica


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Laboratory Experiment
2.2. Microplot Field Experiment
2.3. Statistical Analysis
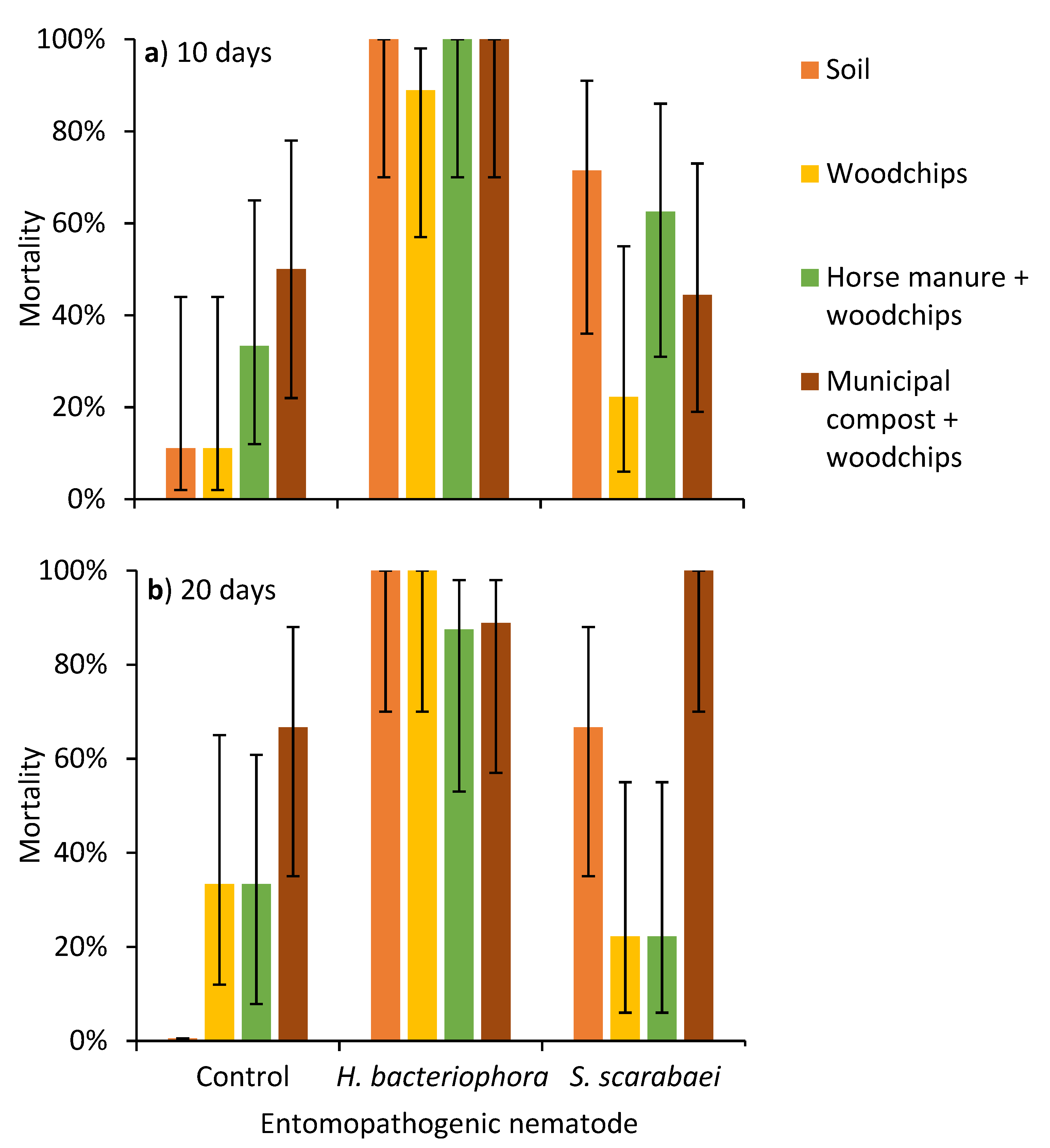
3. Results
3.1. Laboratory Experiment
3.2. Microplot Field Experiment
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Isaacs, R.; Szendrei, Z.; Wise, J.C. Evaluation of new approaches for management of Japanese beetles in highbush blueberries. Small Fruits Rev. 2004, 3, 349–360. [Google Scholar] [CrossRef]
- EPPO Global Database Popillia japonica (POPIJA). Available online: https://gd.eppo.int/taxon/POPIJA/distribution (accessed on 18 August 2021).
- Potter, D.A.; Held, D.W. Biology and management of the Japanese beetle. Annu. Rev. Entomol. 2002, 47, 175–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smitley, D.R. Incidence of Popillia japonica (Coleoptera: Scarabaeidae) and other scarab larvae in nursery fields. J. Econ. Entomol. 1996, 89, 1262–1266. [Google Scholar] [CrossRef]
- [OMAFRA] Ontario Ministry of Agriculture and Rural Affairs. Notes on Blueberry Insects: White Grub Complex. Available online: http://www.omafra.gov.on.ca/english/crops/pub360/notes/bluewhgrub.htm (accessed on 18 August 2021).
- Cowles, R.S.; Alm, S.R.; Villani, M.G. Selective toxicity of halofenozide to exotic white grubs (Coleoptera: Scarabaeidae). J. Econ. Entomol. 1999, 92, 427–434. [Google Scholar] [CrossRef]
- Oliver, J.B.; Reding, M.E.; Youssef, N.N.; Klein, M.G.; Bishop, B.L.; Lewis, P.A. Surface-applied insecticide treatments for quarantine control of Japanese beetle, Popillia japonica Newman (Coleoptera: Scarabaeidae), larvae in field-grown nursery plants. Pest Manag. Sci. 2009, 65, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Georgis, R.; Koppenhöfer, A.M.; Lacey, L.A.; Bélair, G.; Duncan, L.W.; Grewal, P.S.; Samish, M.; Tan, L.; Torr, P.; van Tol, R.W.H.M. Successes and failures in the use of parasitic nematodes for pest control. Biol. Control. 2006, 38, 103–123. [Google Scholar] [CrossRef]
- Torrini, G.; Paoli, F.; Mazza, G.; Simoncini, S.; Benvenuti, C.; Strangi, A.; Tarasco, E.; Barzanti, G.P.; Bosio, G.; Cutino, I.; et al. Evaluation of indigenous entomopathogenic nematodes as potential biocontrol agents against Popillia japonica (Coleoptera: Scarabaeidae) in Northern Italy. Insects 2020, 11, 804. [Google Scholar] [CrossRef] [PubMed]
- Koppenhöfer, A.M.; Fuzy, E.M.; Crocker, R.L.; Gelernter, W.D.; Polavarapu, S. Pathogenicity of Heterorhabditis bacteriophora, Steinernema glaseri, and S. scarabaei (Rhabditida: Heterorhabditidae, Steinernematidae) against 12 white grub species (Coleoptera: Scarabaeidae). Biocontrol. Sci. Technol. 2004, 14, 87–92. [Google Scholar] [CrossRef]
- Koppenhöfer, A.M.; Grewal, P.S.; Fuzy, E.M. Virulence of the entomopathgenic nematodes Heterorhabditis bacteriophora, Heterorhabditis zealandica, Steinernema scarabaei against five white grub species (Coleoptera: Scarabaeidae) of economic importance in turfgrass in North America. Biol. Control. 2006, 38, 397–404. [Google Scholar] [CrossRef]
- Power, K.T.; An, R.; Grewal, P.S. Effectiveness of Heterorhabditis bacteriophora strain GPS11 applications targeted against difference instars of the Japanese beetle Popillia japonica. Biol. Control. 2009, 48, 232–236. [Google Scholar] [CrossRef]
- Grewal, P.S.; Power, K.T.; Grewal, S.K.; Suggars, A.; Haupricht, S. Enhanced consistency in biological control of white grubs (Coleoptera: Scarabaeidae) with new strains of entomopathogenic nematodes. Biol. Control. 2004, 30, 73–82. [Google Scholar] [CrossRef]
- Polavarapu, S.; Koppenhöfer, A.M.; Barry, J.D.; Holdcraft, R.J.; Fuzy, E.M. Entomopathogenic nematodes and neonicotinoids of remedial control of oriental beetle, Anomala orientalis (Coleoptera: Scarabaeidae), in highbush blueberry. Crop Prot. 2007, 26, 1266–1271. [Google Scholar] [CrossRef]
- Magee, J.B.; Spiers, J.M. Influence of mulching systems on yield and quality of southern highbush blueberries. J. Small Fruit Vitic. 1995, 3, 133–141. [Google Scholar] [CrossRef]
- Sciarappa, W.; Polavarapu, S.; Barry, J.; Oudemans, P.; Ehlenfeldt, M.; Pavlis, G.; Polk, D.; Holdcraft, R. Developing an organic production system for highbush blueberry. HortScience 2008, 43, 51–57. [Google Scholar] [CrossRef]
- Burkhard, N.E.; Lynch, D.H.; Percival, D.C.; Sharifi, M. Organic mulch impact on vegetation dynamics and productivity of highbush blueberry under organic production. HortScience 2009, 44, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Pate, E.; (Ontario Ministry of Agriculture, Food and Rural Affairs, Simcoe, ON, Canada). Personal communication, 2021.
- Renkema, J.M.; Lynch, D.H.; Cutler, G.C.; MacKenzie, K.; Walde, S.J. Emergence of blueberry maggot flies (Diptera: Tephritidae) from mulches and soil at various depths. Environ. Entomol. 2012, 41, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Renkema, J.M.; Cutler, G.C.; Lynch, D.H.; MacKenzie, K.; Walde, S.J. Organic mulches in highbush blueberries alter beetle (Coleoptera) community composition and improve functional group abundance and diversity. Agric. For. Entomol. 2016, 18, 119–127. [Google Scholar] [CrossRef]
- Khumalo, N.N.; Lephoto, T.E.; Gray, V.M. The effect of organic compost and soil texture on the survival and infectivity of entomopathogenic nematode species. Arch. Phytopathol. Pflanzenschutz 2021. [Google Scholar] [CrossRef]
- Herren, G.L.; Binnemans, I.; Joos, L.; Viane, N.; Ehlers, R.-U.; Vandecasteele, B.; Bert, W.; Steel, H. Compost as a carrier medium entomopathogenic nematodes—The influence of compost maturity on their virulence and survival. Biol. Control. 2018, 125, 29–38. [Google Scholar] [CrossRef]
- SAS. JMP® Version 15.0.0; SAS Institute Inc.: Cary, NC, USA, 2019. [Google Scholar]
- Koppenhöfer, A.M.; Fuzy, E.M. Soil moisture effects on infectivity and persistence of the entomopathogenic nematodes Steinernema scarabaei, S. glaseri, Heterorhabditis zealandica, and H. bacteriophora. Appl. Soil. Ecol. 2007, 35, 128–139. [Google Scholar] [CrossRef]
- Koppenhöfer, A.M.; Fuzy, E.M. Effect of soil type on infectivity and persistence of the entomopathogenic nematodes Steinernema scarabaei, Steinernema glaseri, Heterorhabditis zealandica and Heterorhabditis bacteriophora. J. Invert. Pathol. 2006, 92, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Khathwayo, Z.; Ramakuwela, T.; Hatting, J.; Shapiro-Ilan, D.I.; Cochrane, N. Quantification of pH tolerance levels among entomopathogenic nematodes. J. Nematol. 2021, 53, 1–12. [Google Scholar] [CrossRef]
- Litterick, A.M.; Harrier, L.; Wallace, P.; Watson, C.A.; Wood, M. The Role of Uncomposted Materials, Composts, Manures, and Compost Extracts in Reducing Pest and Disease Incidence and Severity in Sustainable Temperate Agricultural and Horticultural Crop Production—A Review. Crit. Rev. Plant Sci. 2004, 23, 453–479. [Google Scholar] [CrossRef]
- Mori, N.; Santoiemma, G.; Glazer, I.; Gilioli, G.; Ciampitti, M.; Cavagna, B.; Battisti, A. Management of Popillia japonica in container-grown nursery stock in Italy. Phytoparasitica 2021, 1–7. [Google Scholar] [CrossRef]
- Simões, N.; Laumond, C.; Bonifassi, E. Effectiveness of Steinernema spp. and Heterorhabditis bacteriophora against Popillia japonica in the Azores. J. Nematol. 1993, 25, 480–485. [Google Scholar] [PubMed]
- Marianelli, L.; Paoli, F.; Torrini, G.; Mazza, C.; Benvenuti, F.; Binazzi, G.; Sabbatini Peverieri, G.; Bosio, D.; Venanzio, E.; Giacometto, S.; et al. Entomopathogenic nematodes as potential biological control agents of Popillia japonica (Coleoptera, Scarabaeidae) in Piedmont Region (Italy). J. Appl. Entomol. 2017, 142, 311–318. [Google Scholar] [CrossRef]
- Koppenhöfer, A.M.; Fuzy, E.M. Ecological characterization of Steinernema scarabaei, a scarab-adapted entomopathogenic nematode from New Jersey. J. Invert. Pathol. 2003, 83, 139–148. [Google Scholar] [CrossRef]
- Paoli, F.; Marianelli, L.; Torrini, G.; Mazza, G.; Benvenuti, C.; Bosio, G.; Venanzio, D.; Tarasco, E.; Klein, M.; Roversi, P.F. Differential susceptibility of Popillia japonica 3rd instars to Heterorhabditis bacteriophora (Italian strain) at three different seasons. Biocontrol. Sci. Technol. 2017, 27, 439–444. [Google Scholar] [CrossRef]
- Villani, M.G.; Nyrop, J.P. Age-depdendent movement patterns of Japanese beetle and European chafer (Coleoptera: Scarabaeidae) grubs in soil—Turfgrass microcosms. Environ. Entomol. 1991, 20, 241–251. [Google Scholar] [CrossRef] [Green Version]
- Szendrei, Z.; Mallampalli, N.; Isaacs, R. Effect of tillage on abundance of Japanese beetle, Popillia japonica Newman (Col., Scarabaeidae), larvae and adults in highbush blueberry fields. J. Appl. Entomol. 2005, 129, 258–264. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mulch Type | Morning (°C) | Afternoon (°C) | Maximum (°C) | Minimum (°C) |
|---|---|---|---|---|
| Sand and peat | 08.7 ± 0.41 b | 13.7 ± 0.43 ab | 18.1 ± 0.62 (21 October) | 6.0 ± 0.10 (29 October) |
| Soil | 08.8 ± 0.39 b | 13.3 ± 0.35 ab | 15.9 ± 0.27 (21 October) | 6.7 ± 0.18 (29 October) |
| Woodchips 1 | 09.2 ± 0.38 b | 12.7 ± 0.33 b | 15.9 ± 0.52 (21 October) | 6.6 ± 0.19 (29 October) |
| Woodchips 2 | 08.9 ± 0.39 b | 12.4 ± 0.36 b | 15.3 ± 0.43 (21 October) | 7.1 ± 0.09 (18 October) |
| Municipal compost + woodchips 1 | 10.9 ± 0.41 a | 14.6 ± 0.32 a | 17.4 ± 0.10 (22 October) | 8.8 ± 0.09 (18 October) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Renkema, J.M.; Parent, J.-P. Mulches Used in Highbush Blueberry and Entomopathogenic Nematodes Affect Mortality Rates of Third-Instar Popillia japonica. Insects 2021, 12, 907. https://doi.org/10.3390/insects12100907
Renkema JM, Parent J-P. Mulches Used in Highbush Blueberry and Entomopathogenic Nematodes Affect Mortality Rates of Third-Instar Popillia japonica. Insects. 2021; 12(10):907. https://doi.org/10.3390/insects12100907
Chicago/Turabian StyleRenkema, Justin M., and Jean-Philippe Parent. 2021. "Mulches Used in Highbush Blueberry and Entomopathogenic Nematodes Affect Mortality Rates of Third-Instar Popillia japonica" Insects 12, no. 10: 907. https://doi.org/10.3390/insects12100907
APA StyleRenkema, J. M., & Parent, J.-P. (2021). Mulches Used in Highbush Blueberry and Entomopathogenic Nematodes Affect Mortality Rates of Third-Instar Popillia japonica. Insects, 12(10), 907. https://doi.org/10.3390/insects12100907