
The Paramount Role of Drosophila melanogaster in the Study of Epigenetics: From Simple Phenotypes to Molecular Dissection and Higher-Order Genome Organization



{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
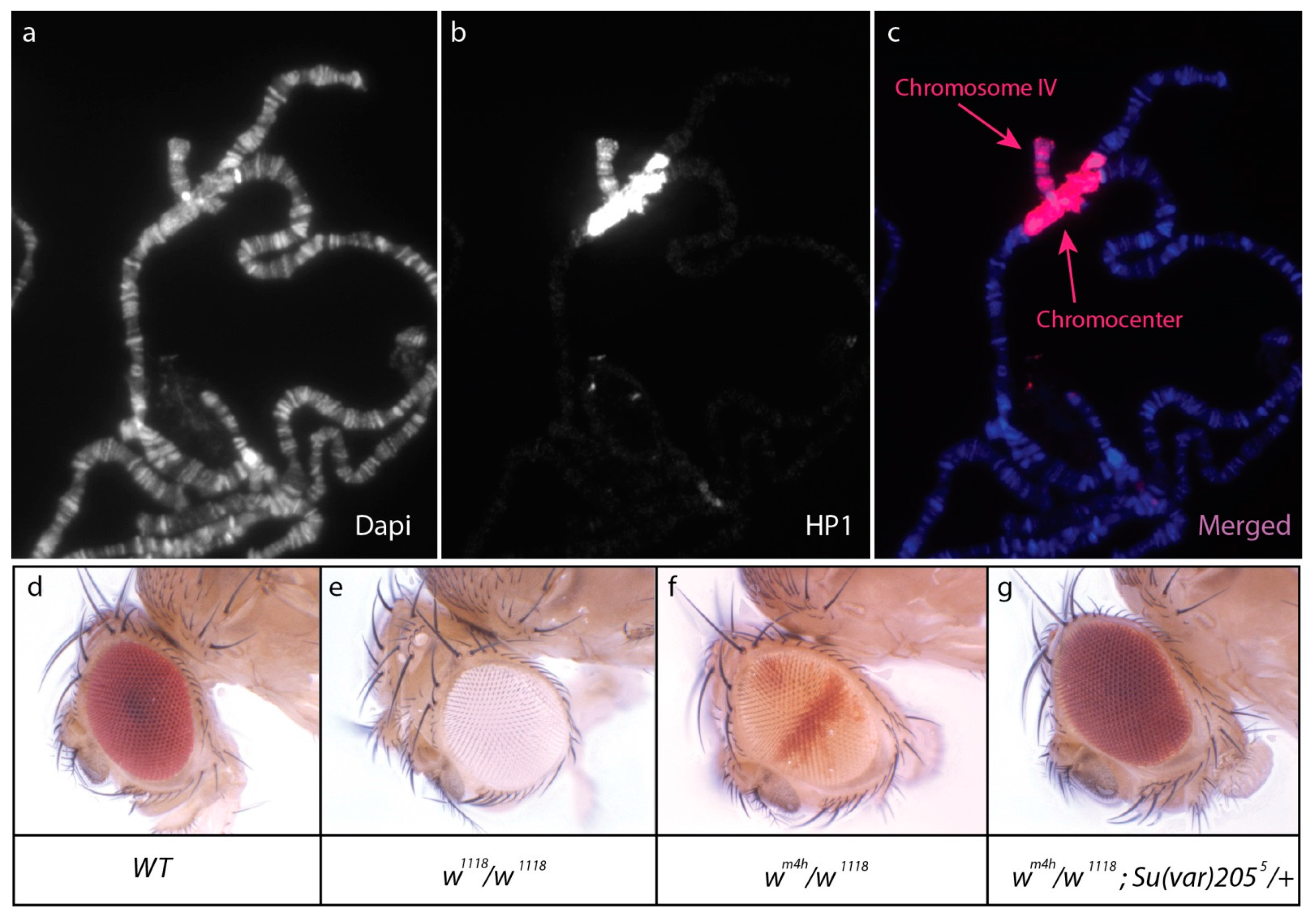
2. Formation and Maintenance of Heterochromatin
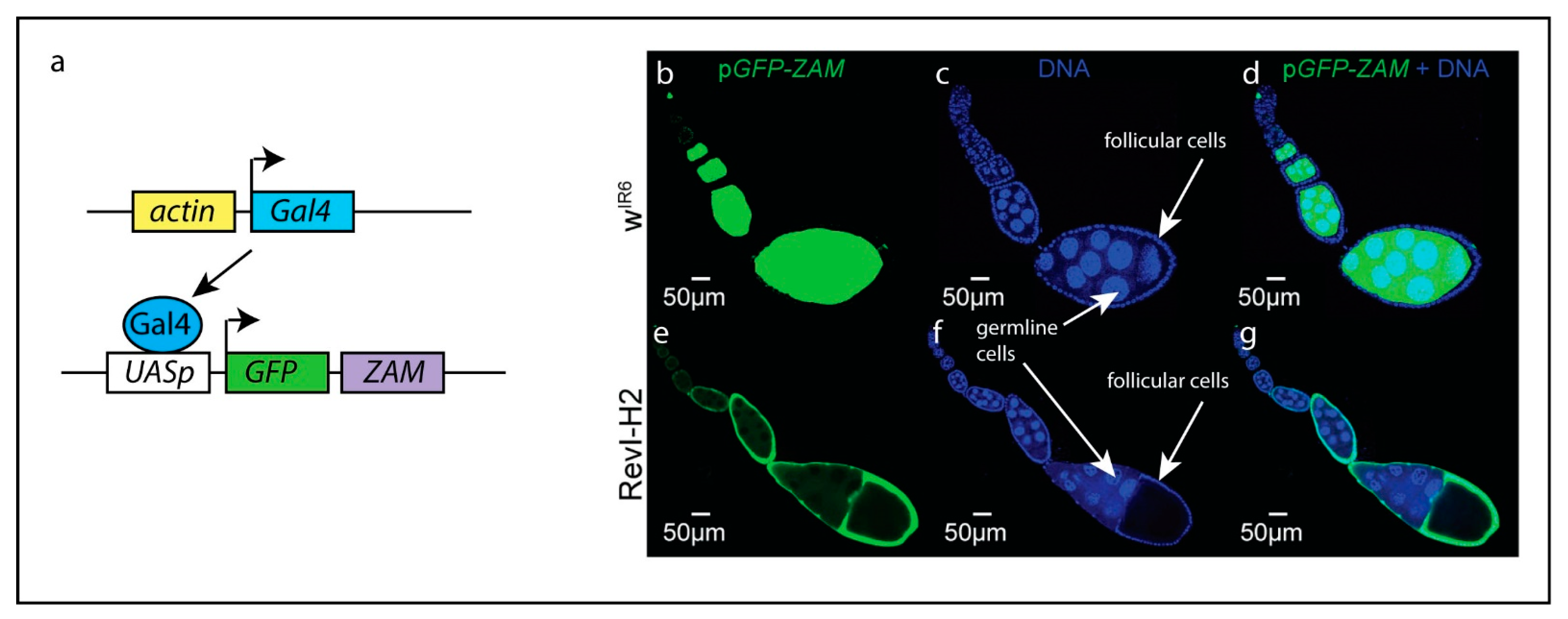
3. Regulation of Transposons by piRNA Clusters
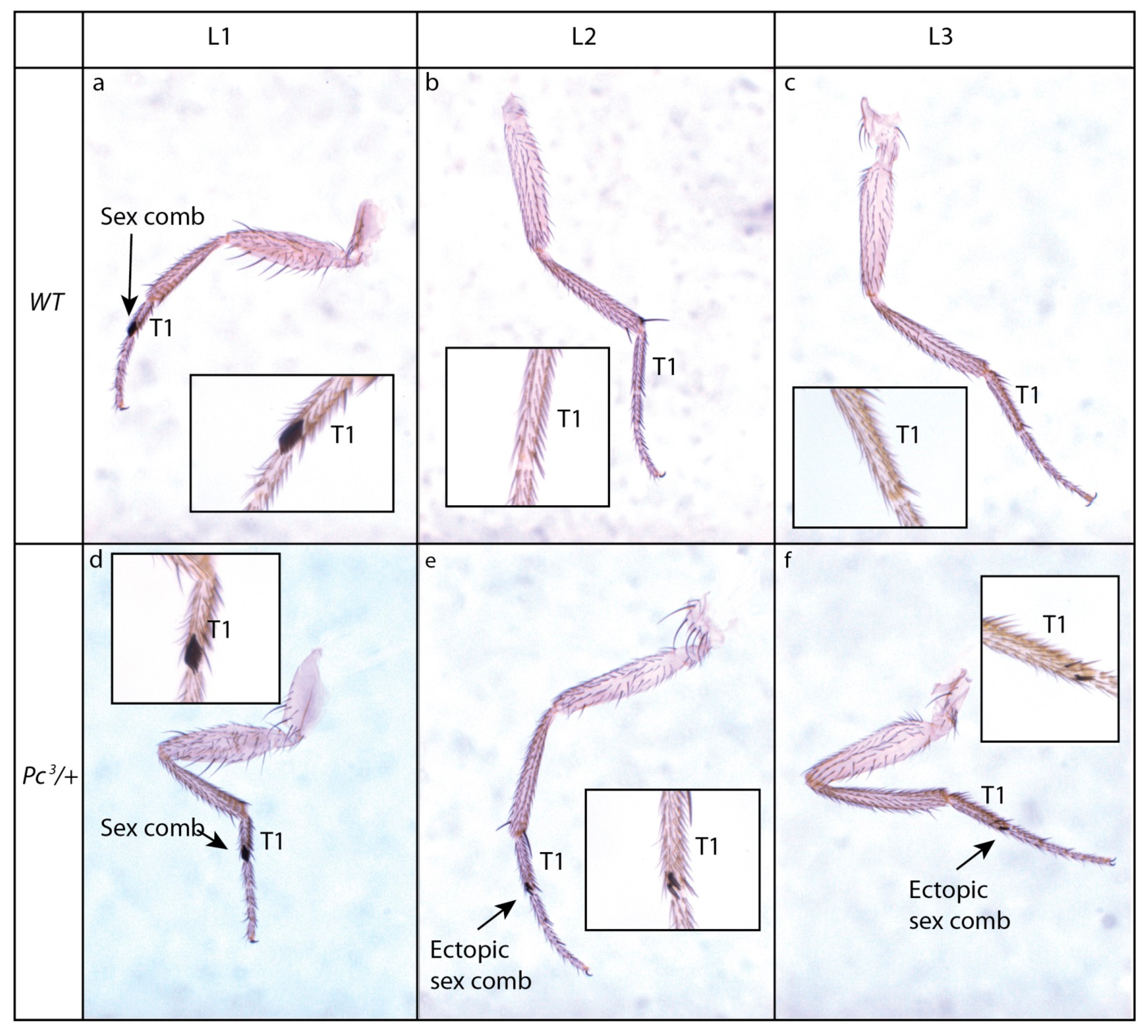
4. Polycomb and Trithorax Complexes and the Maintenance of Chromatin Conformation
5. Testing the Role of Histone Marks in Drosophila
6. Towards an Integrated Vision of Chromatin Domains
7. Drosophila Epigenetics and the Environment
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Waddington, C.H. The Epigenotype. Endeavor 1942, 1, 18–20. [Google Scholar] [CrossRef]
- Russo, V.; Martiensen, R.; Riggs, A. Introduction. In Epigenetic Mechanisms of Gene Regulation; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1996. [Google Scholar]
- Adams, M.D.; Celniker, S.E.; Holt, R.A.; Evans, C.A.; Gocayne, J.D.; Amanatides, P.G.; Scherer, S.E.; Li, P.W.; Hoskins, R.A.; Galle, R.F.; et al. The Genome Sequence of Drosophila melanogaster. Science 2000, 287, 2185–2195. [Google Scholar] [CrossRef]
- Schuettengruber, B.; Bourbon, H.-M.; Di Croce, L.; Cavalli, G. Genome Regulation by Polycomb and Trithorax: 70 Years and Counting. Cell 2017, 171, 34–57. [Google Scholar] [CrossRef]
- Sati, S.; Cavalli, G. Chromosome Conformation Capture Technologies and Their Impact in Understanding Genome Function. Chromosoma 2017, 126, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Janssen, A.; Colmenares, S.U.; Karpen, G.H. Heterochromatin: Guardian of the Genome. Annu. Rev. Cell Dev. Biol. 2018, 34, 265–288. [Google Scholar] [CrossRef]
- Elgin, S.C.R.; Reuter, G. Position-Effect Variegation, Heterochromatin Formation, and Gene Silencing in Drosophila. Cold Spring Harb. Perspect. Biol. 2013, 5, a017780. [Google Scholar] [CrossRef]
- Schotta, G.; Ebert, A.; Dorn, R.; Reuter, G. Position-Effect Variegation and the Genetic Dissection of Chromatin Regulation in Drosophila. Semin. Cell Dev. Biol. 2003, 14, 67–75. [Google Scholar] [CrossRef]
- Sun, F.L.; Cuaycong, M.H.; Craig, C.A.; Wallrath, L.L.; Locke, J.; Elgin, S.C. The Fourth Chromosome of Drosophila melanogaster: Interspersed Euchromatic and Heterochromatic Domains. Proc. Natl. Acad. Sci. USA 2000, 97, 5340–5345. [Google Scholar] [CrossRef]
- Reuter, G.; Wolff, I. Isolation of Dominant Suppressor Mutations for Position-Effect Variegation in Drosophila melanogaster. Mol. Gen. Genet. 1981, 182, 516–519. [Google Scholar] [CrossRef]
- James, T.C.; Elgin, S.C. Identification of a Nonhistone Chromosomal Protein Associated with Heterochromatin in Drosophila melanogaster and Its Gene. Mol. Cell. Biol. 1986, 6, 3862–3872. [Google Scholar] [CrossRef]
- Eissenberg, J.C.; James, T.C.; Foster-Hartnett, D.M.; Hartnett, T.; Ngan, V.; Elgin, S.C. Mutation in a Heterochromatin-Specific Chromosomal Protein Is Associated with Suppression of Position-Effect Variegation in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1990, 87, 9923–9927. [Google Scholar] [CrossRef]
- Fanti, L.; Berloco, M.; Piacentini, L.; Pimpinelli, S. Chromosomal Distribution of Heterochromatin Protein 1 (HP1) in Drosophila: A Cytological Map of Euchromatic HP1 Binding Sites. Genetica 2003, 117, 135–147. [Google Scholar] [CrossRef]
- Salvany, L.; Requena, D.; Azpiazu, N. Functional Association between Eyegone and HP1a Mediates Wingless Transcriptional Repression during Development. Mol. Cell. Biol. 2012, 32, 2407–2415. [Google Scholar] [CrossRef]
- Cryderman, D.E.; Grade, S.K.; Li, Y.; Fanti, L.; Pimpinelli, S.; Wallrath, L.L. Role of Drosophila HP1 in Euchromatic Gene Expression. Dev. Dyn. 2005, 232, 767–774. [Google Scholar] [CrossRef]
- Piacentini, L.; Fanti, L.; Berloco, M.; Perrini, B.; Pimpinelli, S. Heterochromatin Protein 1 (HP1) Is Associated with Induced Gene Expression in Drosophila Euchromatin. J. Cell Biol. 2003, 161, 707–714. [Google Scholar] [CrossRef]
- Levine, M.T.; McCoy, C.; Vermaak, D.; Lee, Y.C.G.; Hiatt, M.A.; Matsen, F.A.; Malik, H.S. Phylogenomic Analysis Reveals Dynamic Evolutionary History of the Drosophila Heterochromatin Protein 1 (HP1) Gene Family. PLoS Genet. 2012, 8, e1002729. [Google Scholar] [CrossRef]
- Vermaak, D.; Henikoff, S.; Malik, H.S. Positive Selection Drives the Evolution of Rhino, a Member of the Heterochromatin Protein 1 Family in Drosophila. PLoS Genet. 2005, 1, 96–108. [Google Scholar] [CrossRef]
- Brower-Toland, B.; Riddle, N.C.; Jiang, H.; Huisinga, K.L.; Elgin, S.C. Multiple SET Methyltransferases Are Required to Maintain Normal Heterochromatin Domains in the Genome of Drosophila melanogaster. Genetics 2009, 181, 1303–1319. [Google Scholar] [CrossRef]
- Schotta, G.; Ebert, A.; Krauss, V.; Fischer, A.; Hoffmann, J.; Rea, S.; Jenuwein, T.; Dorn, R.; Reuter, G. Central Role of Drosophila SU(VAR)3-9 in Histone H3-K9 Methylation and Heterochromatic Gene Silencing. EMBO J. 2002, 21, 1121–1131. [Google Scholar] [CrossRef] [PubMed]
- Seum, C.; Reo, E.; Peng, H.; Rauscher, F.J., 3rd; Spierer, P.; Bontron, S. Drosophila SETDB1 Is Required for Chromosome 4 Silencing. PLoS Genet. 2007, 3, e76. [Google Scholar] [CrossRef] [PubMed]
- Tzeng, T.-Y.; Lee, C.-H.; Chan, L.-W.; Shen, C.-K.J. Epigenetic Regulation of the Drosophila Chromosome 4 by the Histone H3K9 Methyltransferase DSETDB1. Proc. Natl. Acad. Sci. USA 2007, 104, 12691–12696. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, M.L.A.; Philip, P.; Stenberg, P.; Larsson, J. HP1a Recruitment to Promoters Is Independent of H3K9 Methylation in Drosophila melanogaster. PLoS Genet. 2012, 8, e1003061. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.; Deng, H.; Johansen, J.; Girton, J.; Johansen, K.M. Loss-of-Function Alleles of the JIL-1 Histone H3S10 Kinase Enhance Position-Effect Variegation at Pericentric Sites in Drosophila Heterochromatin. Genetics 2007, 176, 1355–1358. [Google Scholar] [CrossRef]
- Zhang, W.; Deng, H.; Bao, X.; Lerach, S.; Girton, J.; Johansen, J.; Johansen, K.M. The JIL-1 Histone H3S10 Kinase Regulates Dimethyl H3K9 Modifications and Heterochromatic Spreading in Drosophila. Development 2006, 133, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Girton, J.; Johansen, J.; Johansen, K.M. A Balance between Euchromatic (JIL-1) and Heterochromatic [SU(Var)2-5 and SU(Var)3-9] Factors Regulates Position-Effect Variegation in Drosophila. Genetics 2011, 188, 745–748. [Google Scholar] [CrossRef]
- Swenson, J.M.; Colmenares, S.U.; Strom, A.R.; Costes, S.V.; Karpen, G.H. The Composition and Organization of Drosophila Heterochromatin Are Heterogeneous and Dynamic. eLife 2016, 5, e16096. [Google Scholar] [CrossRef]
- Strom, A.R.; Emelyanov, A.V.; Mir, M.; Fyodorov, D.V.; Darzacq, X.; Karpen, G.H. Phase Separation Drives Heterochromatin Domain Formation. Nature 2017, 547, 241–245. [Google Scholar] [CrossRef]
- Dernburg, A.F.; Broman, K.W.; Fung, J.C.; Marshall, W.F.; Philips, J.; Agard, D.A.; Sedat, J.W. Perturbation of Nuclear Architecture by Long-Distance Chromosome Interactions. Cell 1996, 85, 745–759. [Google Scholar] [CrossRef]
- Brennecke, J.; Aravin, A.A.; Stark, A.; Dus, M.; Kellis, M.; Sachidanandam, R.; Hannon, G.J. Discrete Small RNA-Generating Loci as Master Regulators of Transposon Activity in Drosophila. Cell 2007, 128, 1089–1103. [Google Scholar] [CrossRef]
- Saito, K.; Nishida, K.M.; Mori, T.; Kawamura, Y.; Miyoshi, K.; Nagami, T.; Siomi, H.; Siomi, M.C. Specific Association of Piwi with RasiRNAs Derived from Retrotransposon and Heterochromatic Regions in the Drosophila Genome. Genes Dev. 2006, 20, 2214–2222. [Google Scholar] [CrossRef]
- Vagin, V.V.; Sigova, A.; Li, C.; Seitz, H.; Gvozdev, V.; Zamore, P.D. A Distinct Small RNA Pathway Silences Selfish Genetic Elements in the Germline. Science 2006, 313, 320–324. [Google Scholar] [CrossRef]
- Coline, G.; Théron, E.; Brasset, E.; Vaury, C. History of the Discovery of a Master Locus Producing PiRNAs: The Flamenco/COM Locus in Drosophila melanogaster. Front. Genet. 2014, 5. [Google Scholar] [CrossRef]
- Rangan, P.; Malone, C.D.; Navarro, C.; Newbold, S.P.; Hayes, P.S.; Sachidanandam, R.; Hannon, G.J.; Lehmann, R. PiRNA Production Requires Heterochromatin Formation in Drosophila. Curr. Biol. 2011, 21, 1373–1379. [Google Scholar] [CrossRef]
- Goriaux, C.; Desset, S.; Renaud, Y.; Vaury, C.; Brasset, E. Transcriptional Properties and Splicing of the Flamenco PiRNA Cluster. EMBO Rep. 2014, 15, 411–418. [Google Scholar] [CrossRef]
- Malone, C.D.; Brennecke, J.; Dus, M.; Stark, A.; McCombie, W.R.; Sachidanandam, R.; Hannon, G.J. Specialized PiRNA Pathways Act in Germline and Somatic Tissues of the Drosophila Ovary. Cell 2009, 137, 522–535. [Google Scholar] [CrossRef] [PubMed]
- Klattenhoff, C.; Xi, H.; Li, C.; Lee, S.; Xu, J.; Khurana, J.S.; Zhang, F.; Schultz, N.; Koppetsch, B.S.; Nowosielska, A.; et al. The Drosophila HP1 Homolog Rhino Is Required for Transposon Silencing and PiRNA Production by Dual-Strand Clusters. Cell 2009, 138, 1137–1149. [Google Scholar] [CrossRef] [PubMed]
- Le Thomas, A.; Stuwe, E.; Li, S.; Du, J.; Marinov, G.; Rozhkov, N.; Chen, Y.-C.A.; Luo, Y.; Sachidanandam, R.; Toth, K.F.; et al. Transgenerationally Inherited PiRNAs Trigger PiRNA Biogenesis by Changing the Chromatin of PiRNA Clusters and Inducing Precursor Processing. Genes Dev. 2014, 28, 1667–1680. [Google Scholar] [CrossRef] [PubMed]
- Akkouche, A.; Mugat, B.; Barckmann, B.; Varela-Chavez, C.; Li, B.; Raffel, R.; Pélisson, A.; Chambeyron, S. Piwi Is Required during Drosophila Embryogenesis to License Dual-Strand PiRNA Clusters for Transposon Repression in Adult Ovaries. Mol. Cell. 2017, 66, 411-419.e4. [Google Scholar] [CrossRef] [PubMed]
- Mohn, F.; Sienski, G.; Handler, D.; Brennecke, J. The Rhino-Deadlock-Cutoff Complex Licenses Noncanonical Transcription of Dual-Strand PiRNA Clusters in Drosophila. Cell 2014, 157, 1364–1379. [Google Scholar] [CrossRef] [PubMed]
- Andersen, P.R.; Tirian, L.; Vunjak, M.; Brennecke, J. A Heterochromatin-Dependent Transcription Machinery Drives PiRNA Expression. Nature 2017, 549, 54–59. [Google Scholar] [CrossRef]
- Sato, K.; Siomi, M.C. The PiRNA Pathway in Drosophila Ovarian Germ and Somatic Cells. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2020, 96, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Sienski, G.; Batki, J.; Senti, K.-A.; Dönertas, D.; Tirian, L.; Meixner, K.; Brennecke, J. Silencio/CG9754 Connects the Piwi-PiRNA Complex to the Cellular Heterochromatin Machinery. Genes Dev. 2015, 29, 2258–2271. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, Y.W.; Murano, K.; Ishizu, H.; Shibuya, A.; Iyoda, Y.; Siomi, M.C.; Siomi, H.; Saito, K. Piwi Modulates Chromatin Accessibility by Regulating Multiple Factors Including Histone H1 to Repress Transposons. Mol. Cell 2016, 63, 408–419. [Google Scholar] [CrossRef]
- Duc, C.; Yoth, M.; Jensen, S.; Mouniée, N.; Bergman, C.M.; Vaury, C.; Brasset, E. Trapping a Somatic Endogenous Retrovirus into a Germline PiRNA Cluster Immunizes the Germline against Further Invasion. Genome Biol. 2019, 20, 127. [Google Scholar] [CrossRef]
- de Vanssay, A.; Bougé, A.-L.; Boivin, A.; Hermant, C.; Teysset, L.; Delmarre, V.; Antoniewski, C.; Ronsseray, S. Paramutation in Drosophila Linked to Emergence of a PiRNA-Producing Locus. Nature 2012, 490, 112–115. [Google Scholar] [CrossRef]
- Lewis, E.B. A Gene Complex Controlling Segmentation in Drosophila. Nature 1978, 276, 565–570. [Google Scholar] [CrossRef]
- Shearn, A.; Hersperger, E.; Hersperger, G. Genetic Studies of Mutations at Two Loci of Drosophila melanogaster Which Cause a Wide Variety of Homeotic Transformations. Rouxs Arch. Dev. Biol. 1987, 196, 231–242. [Google Scholar] [CrossRef]
- Kassis, J.A.; Kennison, J.A.; Tamkun, J.W. Polycomb and Trithorax Group Genes in Drosophila. Genetics 2017, 206, 1699–1725. [Google Scholar] [CrossRef]
- Pattatucci, A.M.; Kaufman, T.C. The Homeotic Gene Sex Combs Reduced of Drosophila melanogaster Is Differentially Regulated in the Embryonic and Imaginal Stages of Development. Genetics 1991, 129, 443–461. [Google Scholar] [CrossRef]
- Paro, R.; Hogness, D.S. The Polycomb Protein Shares a Homologous Domain with a Heterochromatin-Associated Protein of Drosophila. Proc. Natl. Acad. Sci. USA 1991, 88, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Messmer, S.; Franke, A.; Paro, R. Analysis of the Functional Role of the Polycomb Chromo Domain in Drosophila melanogaster. Genes Dev. 1992, 6, 1241–1254. [Google Scholar] [CrossRef][Green Version]
- Saurin, A.J.; Shao, Z.; Erdjument-Bromage, H.; Tempst, P.; Kingston, R.E. A Drosophila Polycomb Group Complex Includes Zeste and DTAFII Proteins. Nature 2001, 412, 655–660. [Google Scholar] [CrossRef]
- Lagarou, A.; Mohd-Sarip, A.; Moshkin, Y.M.; Chalkley, G.E.; Bezstarosti, K.; Demmers, J.A.A.; Verrijzer, C.P. DKDM2 Couples Histone H2A Ubiquitylation to Histone H3 Demethylation during Polycomb Group Silencing. Genes Dev. 2008, 22, 2799–2810. [Google Scholar] [CrossRef]
- Tie, F.; Prasad-Sinha, J.; Birve, A.; Rasmuson-Lestander, A.; Harte, P.J. A 1-Megadalton ESC/E(Z) Complex from Drosophila That Contains Polycomblike and RPD3. Mol. Cell. Biol. 2003, 23, 3352–3362. [Google Scholar] [CrossRef]
- Tie, F.; Furuyama, T.; Prasad-Sinha, J.; Jane, E.; Harte, P.J. The Drosophila Polycomb Group Proteins ESC and E(Z) Are Present in a Complex Containing the Histone-Binding Protein P55 and the Histone Deacetylase RPD3. Development 2001, 128, 275–286. [Google Scholar] [CrossRef]
- Klymenko, T.; Papp, B.; Fischle, W.; Kocher, T.; Schelder, M.; Fritsch, C.; Wild, B.; Wilm, M.; Muller, J. A Polycomb Group Protein Complex with Sequence-Specific DNA-Binding and Selective Methyl-Lysine-Binding Activities. Genes Dev. 2006, 20, 1110–1122. [Google Scholar] [CrossRef] [PubMed]
- Scheuermann, J.C.; de Ayala Alonso, A.G.; Oktaba, K.; Ly-Hartig, N.; McGinty, R.K.; Fraterman, S.; Wilm, M.; Muir, T.W.; Muller, J. Histone H2A Deubiquitinase Activity of the Polycomb Repressive Complex PR-DUB. Nature 2010, 465, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Nekrasov, M.; Klymenko, T.; Fraterman, S.; Papp, B.; Oktaba, K.; Köcher, T.; Cohen, A.; Stunnenberg, H.G.; Wilm, M.; Müller, J. Pcl-PRC2 Is Needed to Generate High Levels of H3-K27 Trimethylation at Polycomb Target Genes. EMBO J. 2007, 26, 4078–4088. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, L.; Oktaba, K.; Scheuermann, J.C.; Gambetta, M.C.; Ly-Hartig, N.; Müller, J. The Role of the Histone H2A Ubiquitinase Sce in Polycomb Repression. Development 2012, 139, 117–127. [Google Scholar] [CrossRef]
- Muller, J.; Hart, C.M.; Francis, N.J.; Vargas, M.L.; Sengupta, A.; Wild, B.; Miller, E.L.; O’Connor, M.B.; Kingston, R.E.; Simon, J.A. Histone Methyltransferase Activity of a Drosophila Polycomb Group Repressor Complex. Cell 2002, 111, 197–208. [Google Scholar] [CrossRef]
- Zenk, F.; Loeser, E.; Schiavo, R.; Kilpert, F.; Bogdanović, O.; Iovino, N. Germ Line-Inherited H3K27me3 Restricts Enhancer Function during Maternal-to-Zygotic Transition. Science 2017, 357, 212–216. [Google Scholar] [CrossRef]
- Francis, N.J.; Kingston, R.E.; Woodcock, C.L. Chromatin Compaction by a Polycomb Group Protein Complex. Science (N. Y.) 2004, 306, 1574–1577. [Google Scholar] [CrossRef]
- Gibert, J.M.; Karch, F. The Polycomb Group Protein CRAMPED Is Involved with TRF2 in the Activation of the Histone H1 Gene. Chromosoma 2011, 120, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Landais, S.; D’Alterio, C.; Jones, D.L. Persistent Replicative Stress Alters Polycomb Phenotypes and Tissue Homeostasis in Drosophila melanogaster. Cell Rep. 2014, 7, 859–870. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Sabath, I.; Kunduru, L.; van Wijnen, A.J.; Marzluff, W.F.; Dominski, Z. A Conserved Interaction That Is Essential for the Biogenesis of Histone Locus Bodies. J. Biol. Chem. 2014, 289, 33767–33782. [Google Scholar] [CrossRef] [PubMed]
- Tatomer, D.C.; Terzo, E.; Curry, K.P.; Salzler, H.; Sabath, I.; Zapotoczny, G.; McKay, D.J.; Dominski, Z.; Marzluff, W.F.; Duronio, R.J. Concentrating Pre-MRNA Processing Factors in the Histone Locus Body Facilitates Efficient Histone MRNA Biogenesis. J. Cell Biol. 2016, 213, 557–570. [Google Scholar] [CrossRef]
- Gambetta, M.C.; Oktaba, K.; Muller, J. Essential Role of the Glycosyltransferase Sxc/Ogt in Polycomb Repression. Science (N. Y.) 2009, 325, 93–96. [Google Scholar] [CrossRef]
- Mohan, M.; Herz, H.-M.; Smith, E.R.; Zhang, Y.; Jackson, J.; Washburn, M.P.; Florens, L.; Eissenberg, J.C.; Shilatifard, A. The COMPASS Family of H3K4 Methylases in Drosophila. Mol. Cell. Biol. 2011, 31, 4310–4318. [Google Scholar] [CrossRef]
- Schmähling, S.; Meiler, A.; Lee, Y.; Mohammed, A.; Finkl, K.; Tauscher, K.; Israel, L.; Wirth, M.; Philippou-Massier, J.; Blum, H.; et al. Regulation and Function of H3K36 Di-Methylation by the Trithorax-Group Protein Complex AMC. Development 2018, 145. [Google Scholar] [CrossRef]
- Mohan, M.; Herz, H.-M.; Takahashi, Y.-H.; Lin, C.; Lai, K.C.; Zhang, Y.; Washburn, M.P.; Florens, L.; Shilatifard, A. Linking H3K79 Trimethylation to Wnt Signaling through a Novel Dot1-Containing Complex (DotCom). Genes Dev. 2010, 24, 574–589. [Google Scholar] [CrossRef]
- Smith, S.T.; Petruk, S.; Sedkov, Y.; Cho, E.; Tillib, S.; Canaani, E.; Mazo, A. Modulation of Heat Shock Gene Expression by the TAC1 Chromatin-Modifying Complex. Nat. Cell Biol. 2004, 6, 162–167. [Google Scholar] [CrossRef]
- Petruk, S.; Sedkov, Y.; Smith, S.; Tillib, S.; Kraevski, V.; Nakamura, T.; Canaani, E.; Croce, C.M.; Mazo, A. Trithorax and DCBP Acting in a Complex to Maintain Expression of a Homeotic Gene. Science 2001, 294, 1331–1334. [Google Scholar] [CrossRef]
- Mohrmann, L.; Langenberg, K.; Krijgsveld, J.; Kal, A.J.; Heck, A.J.; Verrijzer, C.P. Differential Targeting of Two Distinct SWI/SNF-Related Drosophila Chromatin-Remodeling Complexes. Mol. Cell. Biol. 2004, 24, 3077–3088. [Google Scholar] [CrossRef]
- Schuettengruber, B.; Ganapathi, M.; Leblanc, B.; Portoso, M.; Jaschek, R.; Tolhuis, B.; van Lohuizen, M.; Tanay, A.; Cavalli, G. Functional Anatomy of Polycomb and Trithorax Chromatin Landscapes in Drosophila Embryos. PLoS Biol. 2009, 7, e1000013. [Google Scholar] [CrossRef]
- Finogenova, K.; Bonnet, J.; Poepsel, S.; Schäfer, I.B.; Finkl, K.; Schmid, K.; Litz, C.; Strauss, M.; Benda, C.; Müller, J. Structural Basis for PRC2 Decoding of Active Histone Methylation Marks H3K36me2/3. eLife 2020, 9. [Google Scholar] [CrossRef]
- Tie, F.; Banerjee, R.; Stratton, C.A.; Prasad-Sinha, J.; Stepanik, V.; Zlobin, A.; Diaz, M.O.; Scacheri, P.C.; Harte, P.J. CBP-Mediated Acetylation of Histone H3 Lysine 27 Antagonizes Drosophila Polycomb Silencing. Development 2009, 136, 3131–3141. [Google Scholar] [CrossRef]
- Tie, F.; Banerjee, R.; Fu, C.; Stratton, C.A.; Fang, M.; Harte, P.J. Polycomb Inhibits Histone Acetylation by CBP by Binding Directly to Its Catalytic Domain. Proc. Natl. Acad. Sci. USA 2016, 113, E744–E753. [Google Scholar] [CrossRef]
- Francis, N.J.; Saurin, A.J.; Shao, Z.; Kingston, R.E. Reconstitution of a Functional Core Polycomb Repressive Complex. Mol. Cell 2001, 8, 545–556. [Google Scholar] [CrossRef]
- Busturia, A.; Lloyd, A.; Bejarano, F.; Zavortink, M.; Xin, H.; Sakonju, S. The MCP Silencer of the Drosophila Abd-B Gene Requires Both Pleiohomeotic and GAGA Factor for the Maintenance of Repression. Development 2001, 128, 2163–2173. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.K.; Mihaly, J.; Barges, S.; Spierer, A.; Karch, F.; Hagstrom, K.; Schweinsberg, S.E.; Schedl, P. The Iab-7 Polycomb Response Element Maps to a Nucleosome-Free Region of Chromatin and Requires Both GAGA and Pleiohomeotic for Silencing Activity. Mol. Cell. Biol. 2001, 21, 1311–1318. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.H.; Chang, Y.L.; Yang, C.C.; Pan, I.C.; King, B. Pipsqueak Encodes a Factor Essential for Sequence-Specific Targeting of a Polycomb Group Protein Complex. Mol. Cell. Biol. 2002, 22, 6261–6271. [Google Scholar] [CrossRef]
- Blastyak, A.; Mishra, R.K.; Karch, F.; Gyurkovics, H. Efficient and Specific Targeting of Polycomb Group Proteins Requires Cooperative Interaction between Grainyhead and Pleiohomeotic. Mol. Cell. Biol. 2006, 26, 1434–1444. [Google Scholar] [CrossRef]
- Déjardin, J.; Rappailles, A.; Cuvier, O.; Grimaud, C.; Decoville, M.; Locker, D.; Cavalli, G. Recruitment of Drosophila Polycomb Group Proteins to Chromatin by DSP1. Nature 2005, 434, 533–538. [Google Scholar] [CrossRef]
- Enderle, D.; Beisel, C.; Stadler, M.B.; Gerstung, M.; Athri, P.; Paro, R. Polycomb Preferentially Targets Stalled Promoters of Coding and Noncoding Transcripts. Genome Res. 2011, 21, 216–226. [Google Scholar] [CrossRef]
- Gindhart, J.G.; Kaufman, T.C. Identification of Polycomb and Trithorax Group Responsive Elements in the Regulatory Region of the Drosophila Homeotic Gene Sex Combs Reduced. Genetics 1995, 139, 797–814. [Google Scholar] [CrossRef] [PubMed]
- Cavalli, G.; Paro, R. The Drosophila Fab-7 Chromosomal Element Conveys Epigenetic Inheritance during Mitosis and Meiosis. Cell 1998, 93, 505–518. [Google Scholar] [CrossRef]
- Hansen, K.H.; Helin, K. Epigenetic Inheritance through Self-Recruitment of the Polycomb Repressive Complex 2. Epigenetics 2009, 4, 133–138. [Google Scholar] [CrossRef][Green Version]
- Petruk, S.; Sedkov, Y.; Johnston, D.M.; Hodgson, J.W.; Black, K.L.; Kovermann, S.K.; Beck, S.; Canaani, E.; Brock, H.W.; Mazo, A. TrxG and PcG Proteins but Not Methylated Histones Remain Associated with DNA through Replication. Cell 2012, 150, 922–933. [Google Scholar] [CrossRef] [PubMed]
- Lo, S.M.; Follmer, N.E.; Lengsfeld, B.M.; Madamba, E.V.; Seong, S.; Grau, D.J.; Francis, N.J. A Bridging Model for Persistence of a Polycomb Group Protein Complex through DNA Replication in Vitro. Mol. Cell 2012, 46, 784–796. [Google Scholar] [CrossRef] [PubMed]
- Laprell, F.; Finkl, K.; Müller, J. Propagation of Polycomb-Repressed Chromatin Requires Sequence-Specific Recruitment to DNA. Science 2017, 356, 85–88. [Google Scholar] [CrossRef]
- Coleman, R.T.; Struhl, G. Causal Role for Inheritance of H3K27me3 in Maintaining the OFF State of a Drosophila HOX Gene. Science 2017, 356, eaai8236. [Google Scholar] [CrossRef]
- Jenuwein, T.; Allis, C.D. Translating the Histone Code. Science 2001, 293, 1074–1080. [Google Scholar] [CrossRef]
- Hödl, M.; Basler, K. Transcription in the Absence of Histone H3.2 and H3K4 Methylation. Curr. Biol. 2012, 22, 2253–2257. [Google Scholar] [CrossRef]
- Pengelly, A.R.; Copur, Ö.; Jäckle, H.; Herzig, A.; Müller, J. A Histone Mutant Reproduces the Phenotype Caused by Loss of Histone-Modifying Factor Polycomb. Science 2013, 339, 698–699. [Google Scholar] [CrossRef]
- Dorafshan, E.; Kahn, T.G.; Glotov, A.; Savitsky, M.; Walther, M.; Reuter, G.; Schwartz, Y.B. Ash1 Counteracts Polycomb Repression Independent of Histone H3 Lysine 36 Methylation. EMBO Rep. 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Penke, T.J.R.; McKay, D.J.; Strahl, B.D.; Matera, A.G.; Duronio, R.J. Direct Interrogation of the Role of H3K9 in Metazoan Heterochromatin Function. Genes Dev. 2016, 30, 1866–1880. [Google Scholar] [CrossRef]
- Filion, G.J.; van Bemmel, J.G.; Braunschweig, U.; Talhout, W.; Kind, J.; Ward, L.D.; Brugman, W.; de Castro, I.J.; Kerkhoven, R.M.; Bussemaker, H.J.; et al. Systematic Protein Location Mapping Reveals Five Principal Chromatin Types in Drosophila Cells. Cell 2010, 143, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Kharchenko, P.V.; Alekseyenko, A.A.; Schwartz, Y.B.; Minoda, A.; Riddle, N.C.; Ernst, J.; Sabo, P.J.; Larschan, E.; Gorchakov, A.A.; Gu, T.; et al. Comprehensive Analysis of the Chromatin Landscape in Drosophila melanogaster. Nature 2011, 471, 480–485. [Google Scholar] [CrossRef]
- Marshall, O.J.; Brand, A.H. Chromatin State Changes during Neural Development Revealed by in Vivo Cell-Type Specific Profiling. Nat. Commun. 2017, 8, 2271. [Google Scholar] [CrossRef] [PubMed]
- Bantignies, F.; Grimaud, C.; Lavrov, S.; Gabut, M.; Cavalli, G. Inheritance of Polycomb-Dependent Chromosomal Interactions in Drosophila. Genes Dev. 2003, 17, 2406–2420. [Google Scholar] [CrossRef] [PubMed]
- Lanzuolo, C.; Roure, V.; Dekker, J.; Bantignies, F.; Orlando, V. Polycomb Response Elements Mediate the Formation of Chromosome Higher-Order Structures in the Bithorax Complex. Nat. Cell Biol. 2007, 9, 1167–1174. [Google Scholar] [CrossRef]
- Cheutin, T.; Cavalli, G. Progressive Polycomb Assembly on H3K27me3 Compartments Generates Polycomb Bodies with Developmentally Regulated Motion. PLoS Genet. 2012, 8, e1002465. [Google Scholar] [CrossRef]
- Sexton, T.; Cavalli, G. The Role of Chromosome Domains in Shaping the Functional Genome. Cell 2015, 160, 1049–1059. [Google Scholar] [CrossRef]
- Ogiyama, Y.; Schuettengruber, B.; Papadopoulos, G.L.; Chang, J.-M.; Cavalli, G. Polycomb-Dependent Chromatin Looping Contributes to Gene Silencing during Drosophila Development. Mol. Cell 2018, 71, 73–88. [Google Scholar] [CrossRef]
- Cheutin, T.; Cavalli, G. Loss of PRC1 Induces Higher-Order Opening of Hox Loci Independently of Transcription during Drosophila Embryogenesis. Nat. Commun. 2018, 9, 3898. [Google Scholar] [CrossRef] [PubMed]
- Ciabrelli, F.; Comoglio, F.; Fellous, S.; Bonev, B.; Ninova, M.; Szabo, Q.; Xuéreb, A.; Klopp, C.; Aravin, A.; Paro, R.; et al. Stable Polycomb-Dependent Transgenerational Inheritance of Chromatin States in Drosophila. Nat. Genet. 2017, 49, 876–886. [Google Scholar] [CrossRef] [PubMed]
- Fauvarque, M.O.; Dura, J.M. Polyhomeotic Regulatory Sequences Induce Developmental Regulator-Dependent Variegation and Targeted P-Element Insertions in Drosophila. Genes Dev. 1993, 7, 1508–1520. [Google Scholar] [CrossRef] [PubMed]
- Pigliucci, M. Phenotypic Plasticity, Beyond Nature and Nurture; The Johns Hopkins University Press: Baltimore, MD, USA; London, UK, 2001. [Google Scholar]
- Gibert, P.; Moreteau, B.; David, J.R. Developmental Constraints on an Adaptive Plasticity: Reaction Norms of Pigmentation in Adult Segments of Drosophila melanogaster. Evol. Dev. 2000, 2, 249–260. [Google Scholar] [CrossRef]
- Gibert, J.-M.; Mouchel-Vielh, E.; De Castro, S.; Peronnet, F. Phenotypic Plasticity through Transcriptional Regulation of the Evolutionary Hotspot Gene Tan in Drosophila melanogaster. PLoS Genet. 2016, 12, e1006218. [Google Scholar] [CrossRef]
- Casier, K.; Delmarre, V.; Gueguen, N.; Hermant, C.; Viodé, E.; Vaury, C.; Ronsseray, S.; Brasset, E.; Teysset, L.; Boivin, A. Environmentally-Induced Epigenetic Conversion of a PiRNA Cluster. eLife 2019, 8, e39842. [Google Scholar] [CrossRef]
- Harr, B.; Kauer, M.; Schlotterer, C. Hitchhiking Mapping: A Population-Based Fine-Mapping Strategy for Adaptive Mutations in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2002, 99, 12949–12954. [Google Scholar] [CrossRef]
- Levine, M.T.; Begun, D.J. Evidence of Spatially Varying Selection Acting on Four Chromatin-Remodeling Loci in Drosophila melanogaster. Genetics 2008, 179, 475–485. [Google Scholar] [CrossRef][Green Version]
- Voigt, S.; Laurent, S.; Litovchenko, M.; Stephan, W. Positive Selection at the Polyhomeotic Locus Led to Decreased Thermosensitivity of Gene Expression in Temperate Drosophila melanogaster. Genetics 2015, 200, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Gibert, J.M.; Karch, F.; Schlotterer, C. Segregating Variation in the Polycomb Group Gene Cramped Alters the Effect of Temperature on Multiple Traits. PLoS Genet. 2011, 7, e1001280. [Google Scholar] [CrossRef] [PubMed]
- Osborne, A.J.; Dearden, P.K. A “Phenotypic Hangover”: The Predictive Adaptive Response and Multigenerational Effects of Altered Nutrition on the Transcriptome of Drosophila melanogaster. Environ. Epigenet. 2017, 3, dvx019. [Google Scholar] [CrossRef] [PubMed]
- Stern, S.; Fridmann-Sirkis, Y.; Braun, E.; Soen, Y. Epigenetically Heritable Alteration of Fly Development in Response to Toxic Challenge. Cell Rep. 2012, 1, 528–542. [Google Scholar] [CrossRef]
- Siebold, A.P.; Banerjee, R.; Tie, F.; Kiss, D.L.; Moskowitz, J.; Harte, P.J. Polycomb Repressive Complex 2 and Trithorax Modulate Drosophila Longevity and Stress Resistance. Proc. Natl. Acad. Sci. USA 2010, 107, 169–174. [Google Scholar] [CrossRef]
- Duncan, E.J.; Leask, M.P.; Dearden, P.K. Genome Architecture Facilitates Phenotypic Plasticity in the Honeybee (Apis mellifera). Mol. Biol. Evol. 2020, 37, 1964–1978. [Google Scholar] [CrossRef]
- Li, Z.; Mon, H.; Mitsunobu, H.; Zhu, L.; Xu, J.; Lee, J.M.; Kusakabe, T. Dynamics of Polycomb Proteins-Mediated Histone Modifications during UV Irradiation-Induced DNA Damage. Insect. Biochem. Mol. Biol. 2014, 55, 9–18. [Google Scholar] [CrossRef]
- Mitsunobu, H.; Izumi, M.; Mon, H.; Tatsuke, T.; Lee, J.M.; Kusakabe, T. Molecular Characterization of Heterochromatin Proteins 1a and 1b from the Silkworm, Bombyx mori. Insect Mol. Biol. 2012, 21, 9–20. [Google Scholar] [CrossRef]
- Simola, D.F.; Graham, R.J.; Brady, C.M.; Enzmann, B.L.; Desplan, C.; Ray, A.; Zwiebel, L.J.; Bonasio, R.; Reinberg, D.; Liebig, J.; et al. Epigenetic (Re)Programming of Caste-Specific Behavior in the Ant Camponotus floridanus. Science 2016, 351. [Google Scholar] [CrossRef] [PubMed]
- Piunti, A.; Shilatifard, A. The Roles of Polycomb Repressive Complexes in Mammalian Development and Cancer. Nat. Rev. Mol. Cell Biol. 2021, 22, 326–345. [Google Scholar] [CrossRef] [PubMed]
- Coustham, V.; Li, P.; Strange, A.; Lister, C.; Song, J.; Dean, C. Quantitative Modulation of Polycomb Silencing Underlies Natural Variation in Vernalization. Science 2012, 337, 584–587. [Google Scholar] [CrossRef]
- De Lucia, F.; Crevillen, P.; Jones, A.M.E.; Greb, T.; Dean, C. A PHD-Polycomb Repressive Complex 2 Triggers the Epigenetic Silencing of FLC during Vernalization. Proc. Natl. Acad. Sci. USA 2008, 105, 16831–16836. [Google Scholar] [CrossRef] [PubMed]
- Ernst, C.; Odom, D.T.; Kutter, C. The Emergence of PiRNAs against Transposon Invasion to Preserve Mammalian Genome Integrity. Nat. Commun. 2017, 8, 1411. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gibert, J.-M.; Peronnet, F. The Paramount Role of Drosophila melanogaster in the Study of Epigenetics: From Simple Phenotypes to Molecular Dissection and Higher-Order Genome Organization. Insects 2021, 12, 884. https://doi.org/10.3390/insects12100884
Gibert J-M, Peronnet F. The Paramount Role of Drosophila melanogaster in the Study of Epigenetics: From Simple Phenotypes to Molecular Dissection and Higher-Order Genome Organization. Insects. 2021; 12(10):884. https://doi.org/10.3390/insects12100884
Chicago/Turabian StyleGibert, Jean-Michel, and Frédérique Peronnet. 2021. "The Paramount Role of Drosophila melanogaster in the Study of Epigenetics: From Simple Phenotypes to Molecular Dissection and Higher-Order Genome Organization" Insects 12, no. 10: 884. https://doi.org/10.3390/insects12100884
APA StyleGibert, J.-M., & Peronnet, F. (2021). The Paramount Role of Drosophila melanogaster in the Study of Epigenetics: From Simple Phenotypes to Molecular Dissection and Higher-Order Genome Organization. Insects, 12(10), 884. https://doi.org/10.3390/insects12100884