Selection of Predatory Mites for the Biological Control of Potato Tuber Moth in Stored Potatoes



Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Material and Experimental Conditions
2.2. Acceptance Tests of P. operculella Eggs as Prey
2.3. Study of Predation Behaviour at Different Prey Densities: Functional Response
3. Results
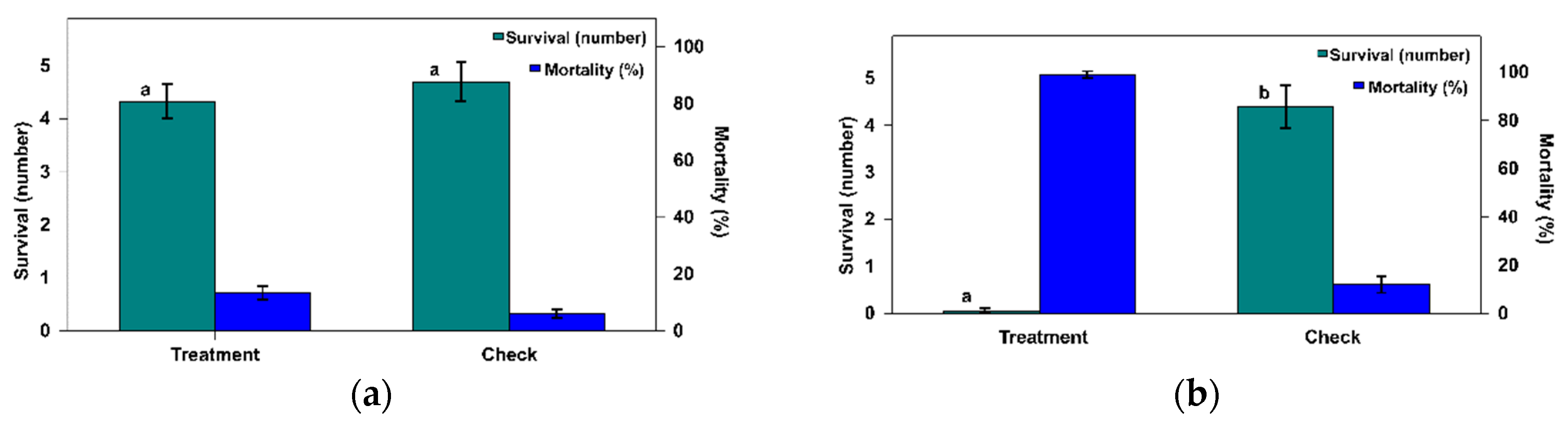
3.1. Acceptance Tests of P. operculella Eggs as Prey
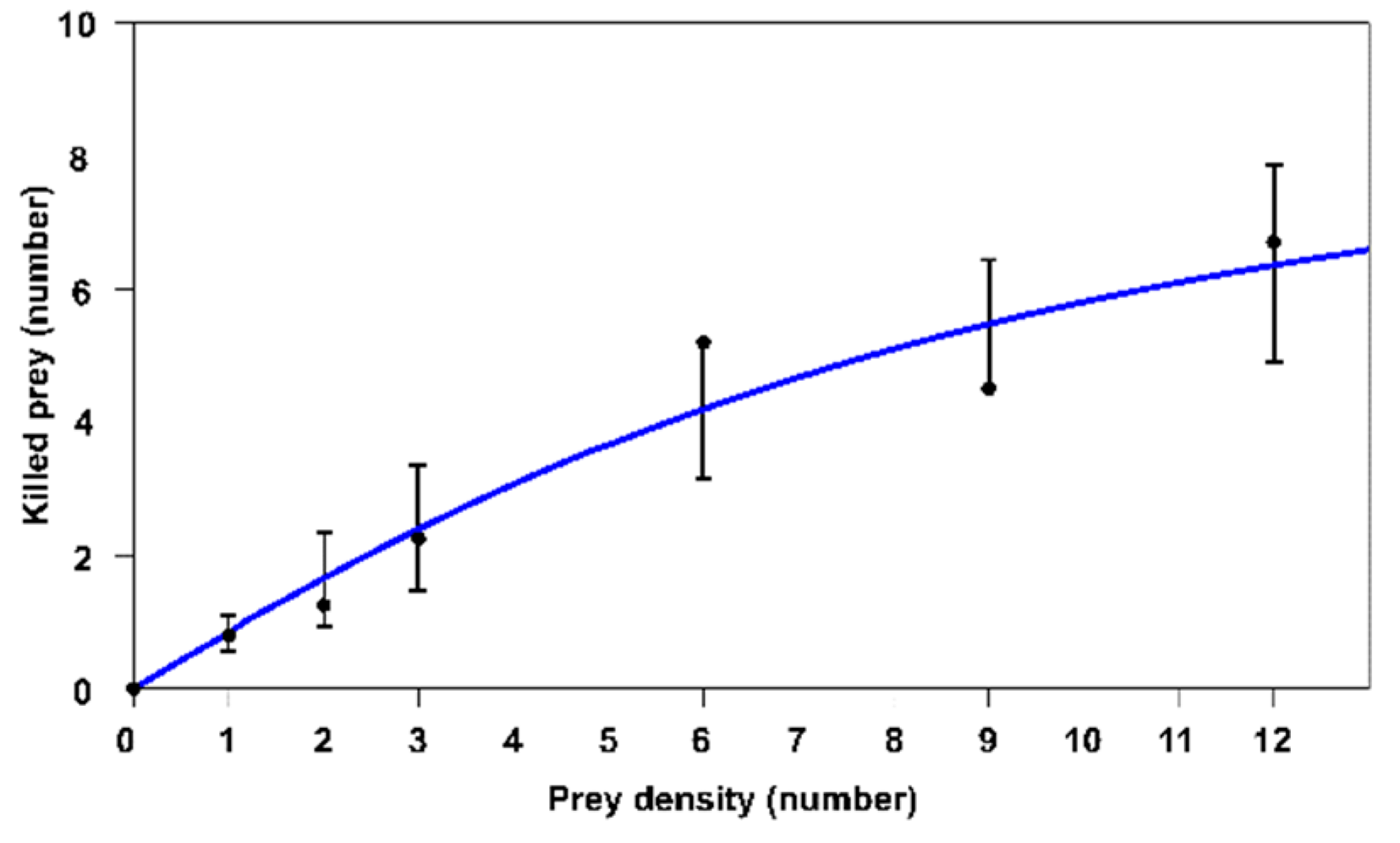
3.2. Study of Predation Behaviour at Different Prey Densities: Functional Response
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- FAOSTAT. Food and Agriculture Organization of the United Nations. FAO Statistics Division 2019. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 15 November 2019).
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Radcliffe, E.B. Insect pests of potato. Annu. Rev. Entomol. 1982, 27, 173–204. [Google Scholar] [CrossRef]
- Kroschel, J.; Schaub, B. Biology and ecology of potato. In Insect Pests of Potato: Global Perspectives on Biology and Management; Giordanengo, P., Vincent, C., Alyokhin, A., Eds.; Elsevier: Waltham, MA, USA, 2013; pp. 165–192. [Google Scholar]
- Das, G.P.; Raman, K.V. Alternate hosts of the potato tuber moth, Phthorimaea operculella. Crop Prot. 1994, 13, 83–86. [Google Scholar] [CrossRef]
- Rondon, S.I. The potato tuberworm: A literature review of its biology, ecology, and control. Am. J. Potato Res. 2010, 87, 149–166. [Google Scholar] [CrossRef]
- Trivedi, T.P.; Rajagopal, D. Distribution, biology, ecology and management of potato tuber moth, Phthorimaea operculella (Lep.: Gelechiidae): A review. Int. J. Pest Manag. 1992, 38, 279–285. [Google Scholar] [CrossRef]
- Hanafi, A. The canon of potato science: 17. Arthropods/insects. Potato Res. 2007, 50, 271–274. [Google Scholar] [CrossRef]
- Aryal, S.; Jung, C. IPM tactics of potato tuber moth, Phthorimaea operculella (Lep.: Gelechiidae): Literature study. Korean J. Soil Zool. 2015, 19, 42–51. [Google Scholar]
- EPPO. European and Mediterranean Plant Protection Organization. EPPO Global Database. Available online: https://gd.eppo.int/taxon/PHTOOP/distribution (accessed on 15 November 2019).
- Kroschel, J.; Canedo, V. Phthorimaea operculella (syn. Gnorimoschema operculella): Potato tuber moth. In Diseases, Pests and Disorders of Potatoes: A (Colour) Handbook; Wale, S., Platt, H.W., Cattlin, N., Eds.; Elsevier Publication: Berkeley, CA, USA, 2008; pp. 124–128. [Google Scholar]
- Alyokhin, A.; Chen, Y.H.; Udalov, M.; Benkovskaya, G.; Lindstrom, L. Evolutionary considerations in potato pest management. In Insect Pests of Potato: Global Perspectives on Biology and Management; Giordanengo, P., Vincent, C., Alyokhin, A., Eds.; Elsevier: Waltham, MA, USA, 2013; pp. 543–571. [Google Scholar]
- Fuglie, K.; Salah, H.B.; Essamet, M.; Temime, A.B.; Rahmouni, A. The development and adoption of integrated pest management of the potato tuber moth, Phthorimaea operculella, in Tunisia. Int. J. Trop. Insect Sci. 1993, 14, 501–509. [Google Scholar] [CrossRef]
- Dent, D. Insect Pest Management, 2nd ed.; CABI: Wallingford, UK, 2000. [Google Scholar]
- Gerson, U.; Smiley, R.L.; Ochoa, R. Mites (Acari) for Pest Control; Blackwell Science Ltd.: Oxford, UK, 2008. [Google Scholar]
- Vila, E.; Cabello, T. Biosystems engineering applied to greenhouse pest control. In Biosystems Engineering: Biofactories for Food Production in the XXI Century; Torres, I., Guevara, R., Eds.; Springer: Cham, Switzerland, 2014; pp. 99–128. [Google Scholar]
- Lefebvre, M.; Langrell, S.R.; Gomez-y-Paloma, S. Incentives and policies for integrated pest management in Europe: A review. Agron. Sustain. Dev. 2015, 35, 27–45. [Google Scholar] [CrossRef]
- Clark, B.; Hillocks, R. Integrated pest management for European agriculture. In Integrated Pest Management; Pimentel, D., Peshin, R., Eds.; Springer: Dordrecht, NL, USA, 2014; pp. 73–97. [Google Scholar]
- Das, G.P.; Magallona, E.D.; Raman, K.V.; Adalla, C.B. Effects of different components of IPM in the management of the potato tuber moth, in storage. Agric. Ecosyst. Environ. 1992, 41, 321–325. [Google Scholar] [CrossRef]
- Simoni, S.; Castagnoli, M. IPM strategies through specialist and generalist phytoseiids (Acari, Mesostigmata). In Integrated Management of Arthropod Pests and Insect Borne Diseases; Ciancio, A., Mukerji, K.G., Eds.; Springer: Dordrecht, NL, USA, 2010; pp. 311–325. [Google Scholar]
- Van Lenteren, J.C. The state of commercial augmentative biological control: Plenty of natural enemies, but a frustrating lack of uptake. BioControl 2012, 57, 1–20. [Google Scholar] [CrossRef]
- De Moraes, G.J.; Venancio, R.; dos Santos, V.L.; Paschoal, A.D. Potential of Ascidae, Blattisociidae and Melicharidae (Acari: Mesostigmata) as biological control agents of pest organisms. In Prospects for Biological Control of Plant Feeding Mites and other Harmful Organisms; Carrillo, D., de Moraes, G.J., Peña, J.E., Eds.; Springer: Cham, Switzerland, 2015; Volume 19, pp. 33–75. [Google Scholar]
- Haines, C.P. Laboratory studies on the role of an egg predator, Blattisocius tarsalis (Acari: Ascidae), in relation to the natural control of Ephestia cautella (Lep.: Pyralidae) in warehouses. Bull. Entomol. Res. 1981, 71, 557–574. [Google Scholar] [CrossRef]
- Nielsen, P.S. The impact of temperature on activity and consumption rate of moth eggs by Blattisocius tarsalis (Acari: Ascidae). Exp. Appl. Acarol. 1999, 23, 149–157. [Google Scholar] [CrossRef]
- Gallego, J.R.; Gamez, M.; Cabello, T. Potential of the Blattisocius mali Mite (Acari: Blattisociidae) as a biological control agent of potato tubermoth (Lep.: Gelechiidae) in stored potatoes. Potato Res. 2019, 1–11. [Google Scholar] [CrossRef]
- Thomas, H.Q.; Zalom, F.G.; Nicola, N.L. Laboratory studies of Blattisocius keegani (Acari: Ascidae) reared on eggs of navel orange worm: Potential for biological control. Bull. Entomol. Res. 2011, 11, 1–6. [Google Scholar] [CrossRef]
- Barker, P.S. Bionomics of Blattisocius keegani (Acarina: Ascidae), a predator on eggs of pests of stored grains. Can. J. Zool. 1967, 45, 1093–1099. [Google Scholar] [CrossRef]
- Krantz, G.W. Review reflections on the biology, morphology and ecology of the Macrochelidae. Exp. Appl. Acarol. 1998, 22, 125–137. [Google Scholar] [CrossRef]
- Azevedo, L.H.; Emberson, R.M.; Esteca, F.C.N.; de Moraes, G.J. Macrochelid mites (Mesostigmata: Macrochelidae) as biological control agents. In Prospects for Biological Control of Plant Feeding Mites and Other Harmful Organisms; Carrillo, D., de Moraes, G.J., Peña, J.E., Eds.; Springer: Cham, Switzerland, 2015; Volume 19, pp. 103–132. [Google Scholar]
- Nesbitt, H.J. A taxonomic study of the Phytoseiinae (family Laelaptidae) predaceous upon Tetranychidae of economic importance. Zool. Verh. Leiden 1951, 12, 1–64. [Google Scholar]
- Haines, C.P. A revision of the genus Blattisocius (Mesostigmata: Ascidae) with especial reference to B. tarsalis and the description of a new species. Acarologia 1978, 20, 19–38. [Google Scholar]
- Greenspan, L. Humidity fixed points of binary saturated aqueous solutions. J. Res. Natl. Bur. Stand. 1977, 81, 89–96. [Google Scholar] [CrossRef]
- Fenemore, P.G. Oviposition of potato tuber moth, Phthorimaea operculella (Lep.: Gelechiidae); fecundity in relation to mated state, age, and pupal weight. N. Z. J. Zool. 1977, 4, 187–191. [Google Scholar] [CrossRef]
- Robertson, J.L.; Preisler, H.K. Pesticide Bioassays with Arthropods; CRC Press: Boca Raton, FL, USA, 1992. [Google Scholar]
- Nielsen, P.S. Predation by Blattisocius tarsalis (Acari: Ascidae) on eggs of Ephestia kuehniella (Lep.: Pyralidae). J. Stored Prod. Res. 2003, 39, 395–400. [Google Scholar] [CrossRef]
- Garcia-Martin, M.; Gamez, M.; Torres-Ruiz, A.; Cabello, T. Functional response of Chelonus oculator (Hym.: Braconidae) to temperature, and its consequences to parasitism. Comm. Ecol. 2008, 9, 45–51. [Google Scholar] [CrossRef]
- Juliano, S. Nonlinear curve fitting: Predation and functional response curves. In Design and Analysis of Ecological Experiments; Scheiner, S., Gurevitch, J., Eds.; Oxford University Press: Oxford, UK, 2001; pp. 178–196. [Google Scholar]
- Hassell, M.P. Arthropod Predator-Prey Systems; Princeton University Press: Princeton, NJ, USA, 1978. [Google Scholar]
- Cabello, T.; Gamez, M.; Varga, Z. An improvement of the Holling type III functional response in entomophagous species model. J. Biol. Syst. 2007, 15, 515–524. [Google Scholar] [CrossRef]
- Azevedo, L.H.; Ferreira, M.P.; de Campos Castilho, R.; Cançado, P.H.D.; de Moraes, G.J. Potential of Macrocheles species (Acari: Mesostigmata: Macrochelidae) as control agents of harmful flies (Dip.) and biology of Macrocheles embersoni on Stomoxys calcitrans and Musca domestica (Dip.: Muscidae). Biol. Control 2018, 123, 1–8. [Google Scholar] [CrossRef]
- Messelink, G.; van Holstein-Saj, R. Improving thrips control by the soil-dwelling predatory mite Macrocheles robustulus. IOBC WPRS Bull. 2008, 32, 135–138. [Google Scholar]
- Pozzebon, A.; Boaria, A.; Duso, C. Single and combined releases of biological control agents against canopy-and soil-dwelling stages of Frankliniella occidentalis in cyclamen. BioControl 2015, 60, 341–350. [Google Scholar] [CrossRef]
- Filipponi, A. The feasibility of mass producing macrochelid mites for field trials against houseflies. Bull. World Health Organ. 1964, 31, 499–501. [Google Scholar]
- Hassan, M.F.; Ali, F.S.; Hussein, A.M.; Mahgoub, M.H. Biological studies on Macrocheles muscaedomesticae fed on different stages of potato tuber moth, Phthorimaea operculella. Egypt. J. Biol. Pest Control 2002, 12, 43–46. [Google Scholar]
- Axtell, R.C. Role of mesostigmatid mites in integrated fly control. Mod. Acarol. 1991, 2, 639–649. [Google Scholar]
- Trivedi, T.P.; Rajagopal, D.; Tandon, P.L. Life table for establishment of potato tubermoth Phthorimaea operculella. J. Indian Potato Assoc. 1994, 21, 97–105. [Google Scholar]
- CABI. Centre for Agricultural Bioscience International. Phthorimaea operculella (potato tuber moth). Available online: http://www.cabi.org/isc/datasheet/40686 (accessed on 15 November 2019).
- Darst, P.H.; King, E.W. Biology of Melichares tarsalis in association with Plodia interpunctella. Ann. Entomol. Soc. Am. 1969, 62, 747–749. [Google Scholar] [CrossRef]
- Riudavets, J.; Maya, M.; Monserrat, M. Predation by Blattisocius tarsalis (Acari: Ascidae) on stored product pests. IOBC WPRS Bull. 2002, 25, 121–126. [Google Scholar]
- Lindquist, E.E.; Krantz, G.W.; Walter, D.E. Order Mesostigmata. In A Manual of Acarology, 3rd ed.; Krantz, G.W., Walter, D.E., Eds.; Texas Tech University Press: Lubbock, TX, USA, 2009; pp. 124–232. [Google Scholar]
- Treat, A.E. Association of the mite Blattisocius tarsalis with the moth Epizeuxis aemula. In Proceedings of the 3rd International Congress of Acarology, Prague, CZ, USA, 31 August–6 September 1971; Daniel, M., Rosicky, B., Eds.; Springer: Dordrecht, NL, USA, 1973; pp. 171–175. [Google Scholar]
- Nielsen, P.S. Developmental time of Blattisocius tarsalis (Acari: Ascidae) at different temperatures. Exp. Appl. Acarol. 2001, 25, 605–608. [Google Scholar] [CrossRef]
- Thind, B.B.; Ford, H.L. Laboratory studies on the use of two new arenas to evaluate the impact of the predatory mites Blattisocius tarsalis and Cheyletus eruditus on residual populations of the stored product mite Acarus siro. Exp. Appl. Acarol. 2006, 38, 167–180. [Google Scholar] [CrossRef]
- Hajek, A.E.; Eilenberg, J. Natural Enemies: An Introduction to Biological Control; Cambridge University Press: Cambridge, UK, 2018. [Google Scholar]



{kind=link}
{kind=link}
| Parameter | Value | SE | 95% Confidence Level | F.R. Type | |
|---|---|---|---|---|---|
| P0 (Intercept) | 1.3021 | 0.4021 | 0.5139 | 2.0902 | II |
| P1 (Linear) | −0.2499 | 0.1187 | −0.4825 | −0.0073 | |
| Type | Fit Curve Parameters (±SE) | Statistical Parameters | ||||
|---|---|---|---|---|---|---|
| a’ (day−1) | Th (day) | α | d.f. | R2 | AICC | |
| I | 0.8907 (±0.1175) | ̶ | ̶ | 6 | 0.8996 | 0.3989 |
| II | 2.1258 (±1.2096) | 0.1014 (± 0.0419) | ̶ | 5 | 0.9408 | 0.3106* |
| III | ̶ | 0.1678 (± 0.0193) | 0.6339 (± 0.2725) | 5 | 0.9393 | 0.3933 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallego, J.R.; Caicedo, O.; Gamez, M.; Hernandez, J.; Cabello, T. Selection of Predatory Mites for the Biological Control of Potato Tuber Moth in Stored Potatoes. Insects 2020, 11, 196. https://doi.org/10.3390/insects11030196
Gallego JR, Caicedo O, Gamez M, Hernandez J, Cabello T. Selection of Predatory Mites for the Biological Control of Potato Tuber Moth in Stored Potatoes. Insects. 2020; 11(3):196. https://doi.org/10.3390/insects11030196
Chicago/Turabian StyleGallego, Juan R., Otto Caicedo, Manuel Gamez, Joaquin Hernandez, and Tomas Cabello. 2020. "Selection of Predatory Mites for the Biological Control of Potato Tuber Moth in Stored Potatoes" Insects 11, no. 3: 196. https://doi.org/10.3390/insects11030196
APA StyleGallego, J. R., Caicedo, O., Gamez, M., Hernandez, J., & Cabello, T. (2020). Selection of Predatory Mites for the Biological Control of Potato Tuber Moth in Stored Potatoes. Insects, 11(3), 196. https://doi.org/10.3390/insects11030196