

The First Analysis of Synaptonemal Complexes in Jawless Vertebrates: Chromosome Synapsis and Transcription Reactivation at Meiotic Prophase I in the Lamprey Lampetra fluviatilis (Petromyzontiformes, Cyclostomata)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Lampreys
2.2. Meiotic Chromosome Studies: Immunostaining Procedure, Controls and Image Analysis
2.3. Electron Microscopy
3. Results
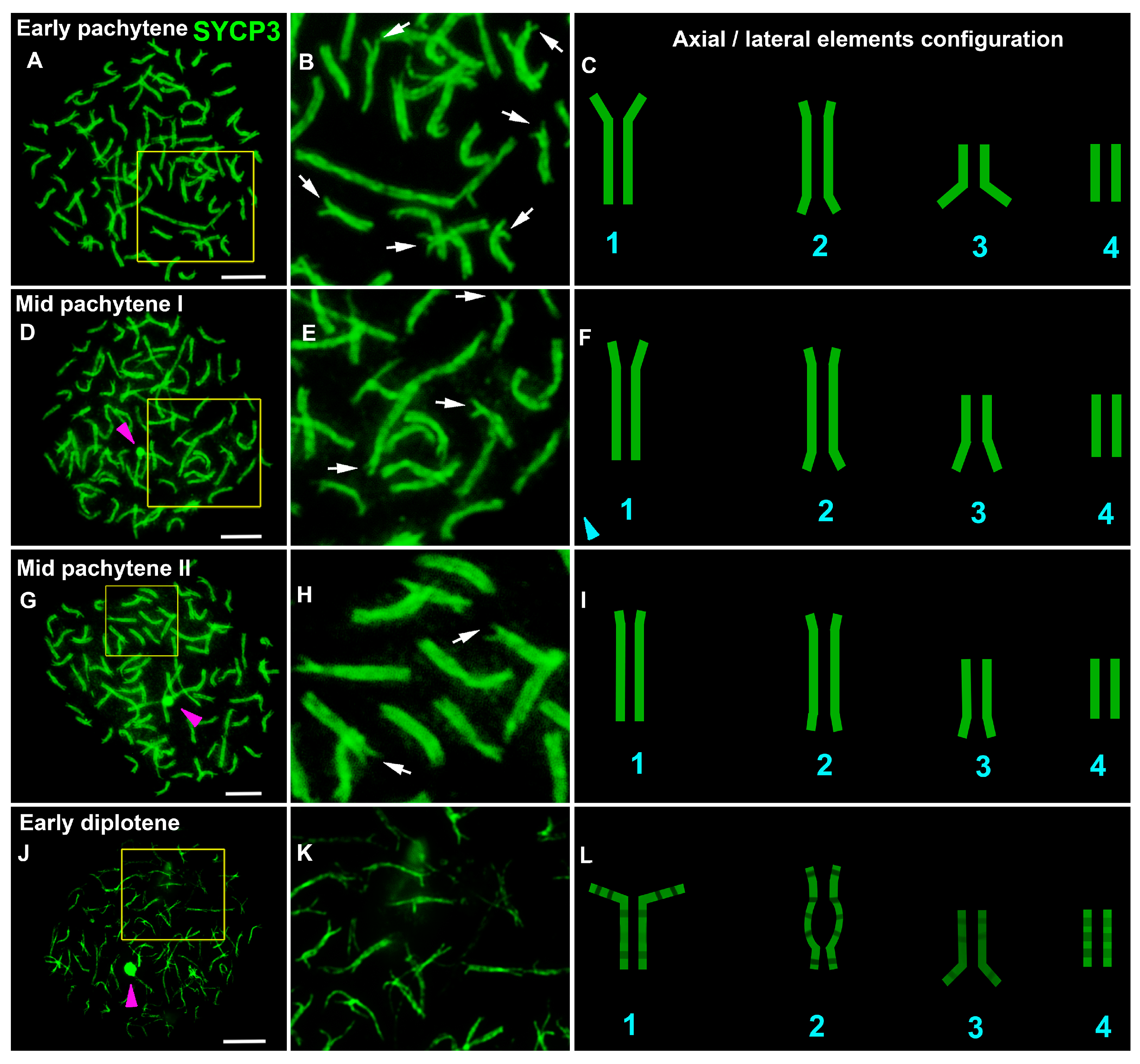
3.1. Lamprey Chromosome Synapsis
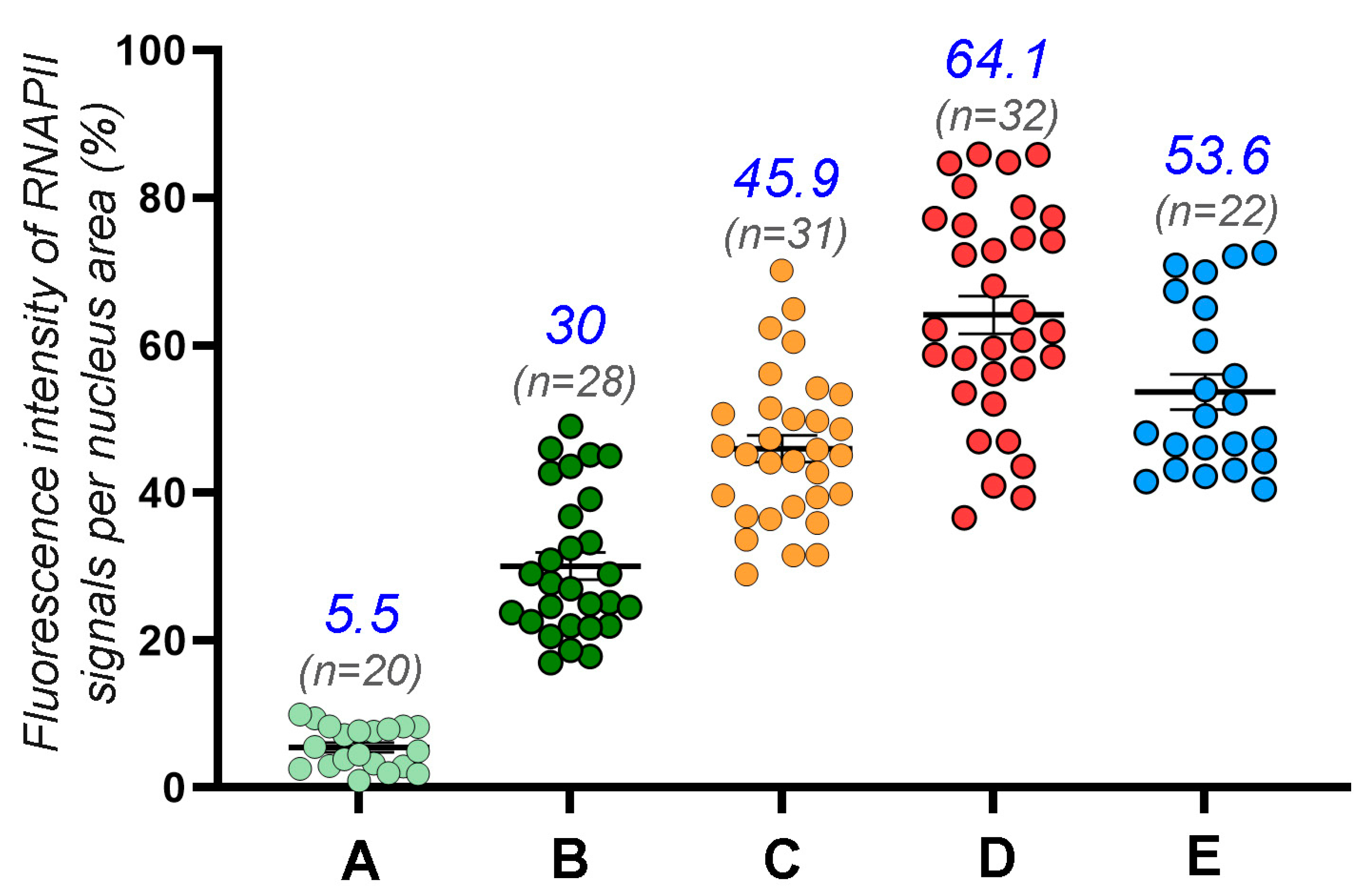
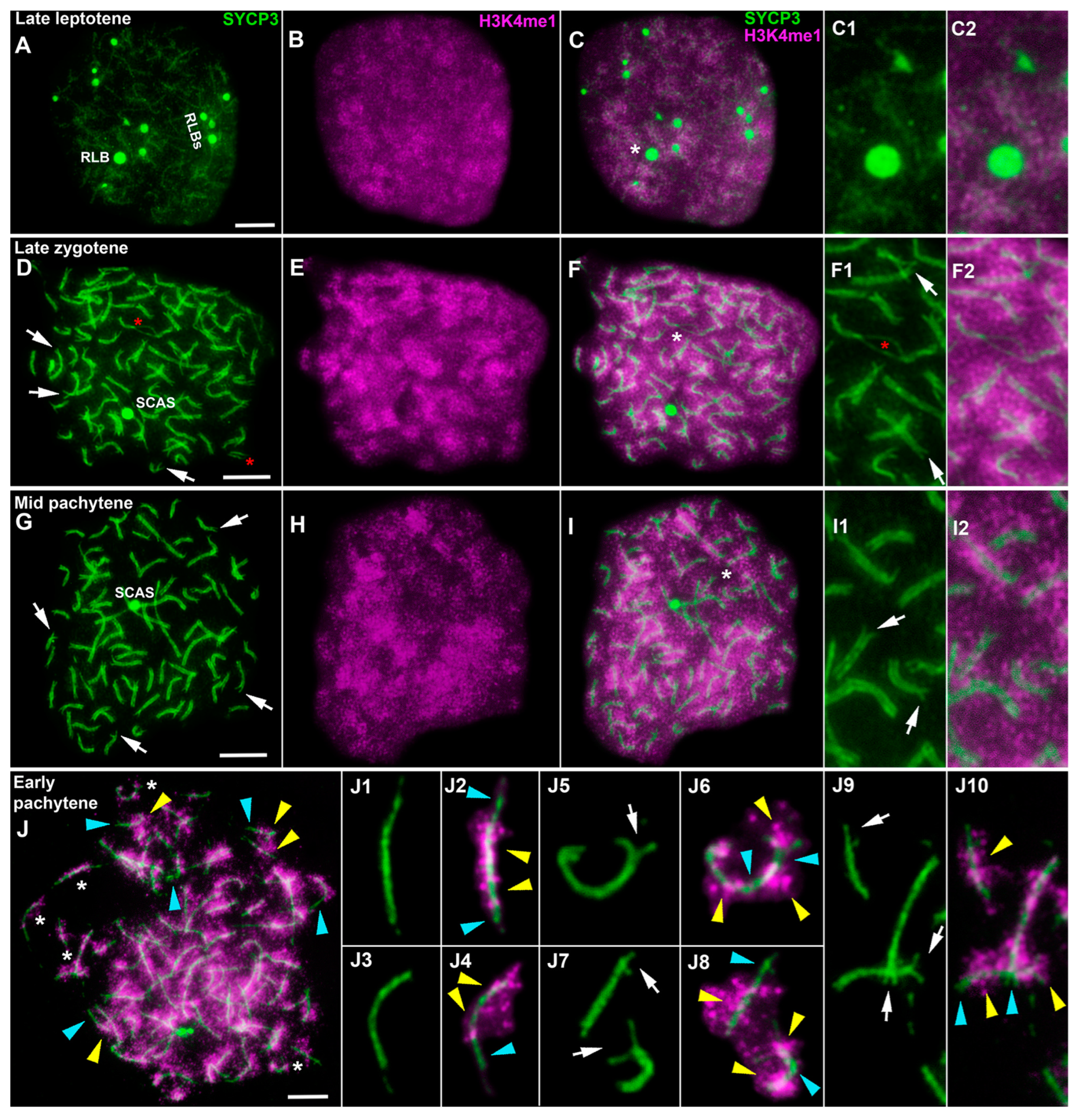
3.2. Transcription in the First Meiotic Prophase in the Lamprey
4. Discussion
4.1. Not All Chromosomes Show Completed Synapsis
4.2. Lamprey Prophase I Is Characterized by Some Unique Epigenetic Patterns
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bogdanov, Y.F.; Kolomiets, O.L. Synaptonemal Complex as an Indicator of the Dynamics of Meiosis and Chromosome Variation; KMK Press: Moscow, Russia, 2007; p. 359. [Google Scholar]
- Loidl, J. Conservation and variability of meiosis across the eukaryotes. Annu. Rev. Genet. 2016, 50, 293–316. [Google Scholar] [CrossRef] [PubMed]
- Alavattam, K.G.; Maezawa, S.; Sakashita, A.; Khoury, H.; Barski, A.; Kaplan, N.; Namekawa, S.H. Attenuated chromatin compartmentalization in meiosis and its maturation in sperm development. Nat. Struct. Mol. Biol. 2019, 26, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Berrios, S. Nuclear architecture of mouse spermatocytes: Chromosome topology, heterochromatin, and nucleolus. Cytogent. Genome Res. 2017, 151, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Zetka, M.; Paouneskou, D.; Jantsch, V. The nuclear envelope, a meiotic jack-of-all-trades. Cur. Opin. Cell Biol. 2020, 64, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Matveevsky, S.; Tretiakov, A.; Kashintsova, A.; Bakloushinskaya, I.; Kolomiets, O. Meiotic nuclear architecture in distinct mole vole hybrids with Robertsonian translocations: Chromosome chains, stretched centromeres, and distorted recombination. IJMS 2020, 21, 7630. [Google Scholar] [CrossRef] [PubMed]
- Fallahi, M.; Getun, I.V.; Wu, Z.K.; Bois, P.R. A global expression switch marks pachytene initiation during mouse male meiosis. Genes 2010, 1, 469–483. [Google Scholar] [CrossRef]
- Ball, R.L.; Fujiwara, Y.; Sun, F.; Hu, J.; Hibbs, M.A.; Handel, M.A.; Carter, G.W. Regulatory complexity revealed by integrated cytological and RNA-seq analyses of meiotic substages in mouse spermatocytes. BMC Genom. 2016, 17, 628. [Google Scholar] [CrossRef]
- Da Cruz, I.; Rodríguez-Casuriaga, R.; Santiñaque, F.F.; Farías., J.; Curti, G.; Capoano, C.A.; Folle, G.A.; Benavente, R.; Sotelo-Silveira, J.R.; Geisinger, A. Transcriptome analysis of highly purified mouse spermatogenic cell populations: Gene expression signatures switch from meiotic-to postmeiotic-related processes at pachytene stage. BMC Genom. 2016, 17, 294. [Google Scholar] [CrossRef]
- Fine, A.D.; Ball, R.L.; Fujiwara, Y.; Handel, M.A.; Carter, G.W. Uncoupling of transcriptomic and cytological differentiation in mouse spermatocytes with impaired meiosis. Mol. Biol. Cell 2019, 30, 717–728. [Google Scholar] [CrossRef]
- Khalil, A.M.; Driscoll, D.J. Epigenetic regulation of pericentromeric heterochromatin during mammalian meiosis. Cytogent. Genome Res. 2010, 129, 280–289. [Google Scholar] [CrossRef]
- Page, J.; De La Fuente, R.; Manterola, M.; Parra, M.T.; Viera, A.; Berríos, S.; Fernández-Donoso, R.; Rufas, J.S. Inactivation or non-reactivation: What accounts better for the silence of sex chromosomes during mammalian male meiosis? Chromosoma 2012, 121, 307–326. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente, R.; Pratto, F.; Hernández-Hernández, A.; Manterola, M.; López-Jiménez, P.; Gómez, R.; Page, J. Epigenetic Dysregulation of Mammalian Male Meiosis Caused by Interference of Recombination and Synapsis. Cells 2021, 10, 2311. [Google Scholar] [CrossRef]
- Wang, L.; Xu, Z.; Khawar, M.B.; Liu, C.; Li, W. The histone codes for meiosis. Reproduction 2017, 154, R65–R79. [Google Scholar] [CrossRef]
- Turner, J.; Mahadevaiah, S.K.; Fernandez-Capetillo, O.; Nussenzweig, A.; Xu, X.; Deng, C.X.; Burgoyne, P.S. Silencing of unsynapsed meiotic chromosomes in the mouse. Nat. Genet. 2005, 37, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Rogakou, E.P.; Pilch, D.R.; Orr, A.H.; Ivanova, V.S.; Bonner, W.M. DNA double-stranded breaks induce histone H2AX phosphorylation on serine 139. J. Biol. Chem. 1998, 273, 5858–5868. [Google Scholar] [CrossRef] [PubMed]
- Mahadevaiah, S.K.; Turner, J.; Baudat, F.; Rogakou, E.P.; de Boer, P.; Blanco-Rodríguez, J.; Jasin, M.; Keeney, S.; Bonner, W.M.; Burgoyne, P.S. Recombinational DNA double-strand breaks in mice precede synapsis. Nat. Genet. 2001, 27, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Capetillo, O.; Lee, A.; Nussenzweig, M.; Nussenzweig, A. H2AX: The histone guardian of the genome. DNA Repair 2004, 3, 959–967. [Google Scholar] [CrossRef] [PubMed]
- Manterola, M.; Brown, T.M.; Oh, M.Y.; Garyn, C.; Gonzalez, B.J.; Wolgemuth, D.J. BRDT is an essential epigenetic regulator for proper chromatin organization, silencing of sex chromosomes and crossover formation in male meiosis. PLoS Genet. 2018, 14, e1007209. [Google Scholar] [CrossRef] [PubMed]
- Quintella, B.R.; Clemens, B.J.; Sutton, T.M.; Lança, M.J.; Madenjian, C.P.; Happel, A.; Harvey, C.J. At-sea feeding ecology of parasitic lampreys. J. Great Lakes Res. 2021, 47, S72–S89. [Google Scholar] [CrossRef]
- Janvier, P. Early Vertebrates; Clarendon Press: Oxford, UK, 1996; p. 393. [Google Scholar]
- Hedges, S.B. Molecular evidence for the early history of living vertebrates. In Major Events in Early Vertebrate Evolution: Paleontology, Phylogeny, Genetics and Development; Ahlberg, P.E., Ed.; Taylor and Francis: London, UK; New York, NY, USA, 2001; pp. 119–134. [Google Scholar]
- Janvier, P. Modern look for ancient lamprey. Nature 2006, 443, 921–923. [Google Scholar] [CrossRef]
- Gess, R.W.; Coates, M.I.; Rubidge, B.S. A lamprey from the Devonian period of South Africa. Nature 2006, 443, 981–984. [Google Scholar] [CrossRef]
- Bayramov, A.V.; Ermakova, G.V.; Kucheryavyy, A.V.; Meintser, I.V.; Zaraisky, A.G. Lamprey as Laboratory Model for Study of Molecular Bases of Ontogenesis and Evolutionary History of Vertebrata. J. Ichthyol. 2022, 62, 1213–1229. [Google Scholar] [CrossRef]
- Timoshevskiy, V.A.; Lampman, R.T.; Hess, J.E.; Porter, L.L.; Smith, J.J. Deep ancestry of programmed genome rearrangement in lampreys. Develop. Biol. 2017, 429, 31–34. [Google Scholar] [CrossRef]
- Timoshevskiy, V.A.; Timoshevskaya, N.Y.; Smith, J.J. Germline-specific repetitive elements in programmatically eliminated chromosomes of the sea lamprey (Petromyzon marinus). Genes 2019, 10, 832. [Google Scholar] [CrossRef] [PubMed]
- Nagao, K.; Otsuzumi, T.; Chinone, H.; Sasaki, T.; Yoshimoto, J.; Matsuda, M.; Kubota, S.; Goto, Y. Novel selectively amplified DNA sequences in the germline genome of the Japanese hagfish, Eptatretus burgeri. Sci. Rep. 2022, 12, 21373. [Google Scholar] [CrossRef] [PubMed]
- Peters, A.H.F.M.; Plug, A.W.; van Vugt, M.J.; de Boer, P. A drying-down technique for the spreading of mammalian meiocytes from the male and female germ line. Chromosome Res. 1997, 5, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Page, J.; Berríos, S.; Rufas, J.S.; Parra, M.T.; Suja, J.Á.; Heyting, C.; Fernández-Donoso, R. The pairing of X and Y chromosomes during meiotic prophase in the marsupial species Thylamys elegans is maintained by a dense plate developed from their axial elements. J. Cell Sci. 2003, 116, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Kolomiets, O.; Matveevsky, S.; Bakloushinskaya, I. Sexual dimorphism in prophase I of meiosis in the Northern mole vole (Ellobius talpinus Pallas, 1770) with isomorphic (XX) chromosomes in males and females. Comp. Cytogenet. 2010, 4, 55. [Google Scholar] [CrossRef]
- Matveevsky, S.; Bakloushinskaya, I.; Kolomiets, O. Unique sex chromosome systems in Ellobius: How do male XX chromosomes recombine and undergo pachytene chromatin inactivation? Sci. Rep. 2016, 6, 29949. [Google Scholar] [CrossRef]
- Matveevsky, S.; Chassovnikarova, T.; Grishaeva, T.; Atsaeva, M.; Malygin, V.; Bakloushinskaya, I.; Kolomiets, O. Kinase CDK2 in mammalian meiotic prophase I: Screening for hetero-and homomorphic sex chromosomes. Int. J. Mol. Sci. 2021, 22, 1969. [Google Scholar] [CrossRef]
- Dresser, M.E.; Moses, M.J. Synaptonemal complex karyotyping in spermatocytes of the Chinese hamster (Cricetulus griseus). Chromosoma 1980, 76, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Van Eenennaam, A.L.; Murray, J.D.; Medrano, J.F. Synaptonemal complex analysis in spermatocytes of white sturgeon, Acipenser transmontanus Richardson (Pisces, Acipenseridae), a fish with a very high chromosome number. Genome 1998, 41, 51–61. [Google Scholar] [CrossRef]
- Cheng, J.; Blum, R.; Bowman, C.; Hu, D.; Shilatifard, A.; Shen, S.; Dynlacht, B.D. A role for H3K4 monomethylation in gene repression and partitioning of chromatin readers. Mol. Cell 2014, 53, 979–992. [Google Scholar] [CrossRef] [PubMed]
- Robinson, E.S.; Potter, I.C. Meiotic chromosomes of Mordacia praecox and a discussion of chromosome numbers in lampreys. Copeia 1969, 4, 824–828. [Google Scholar] [CrossRef]
- Robinson, E.S.; Potter, I.C. The chromosomes of the southern hemispheric lamprey, Geotria australis Gray. Experientia 1981, 37, 239–240. [Google Scholar] [CrossRef]
- Covelo-Soto, L.; Morán, P.; Pasantes, J.J.; Pérez-García, C. Cytogenetic evidences of genome rearrangement and differential epigenetic chromatin modification in the sea lamprey (Petromyzon marinus). Genetica 2014, 142, 545–554. [Google Scholar] [CrossRef]
- Roeder, G.S.; Bailis, J.M. The pachytene checkpoint. Trend. Genet. 2000, 16, 395–403. [Google Scholar] [CrossRef]
- Burgoyne, P.S.; Mahadevaiah, S.K.; Turner, J.M.A. The consequences of asynapsis for mammalian meiosis. Nat. Rev. Genet. 2009, 10, 207–216. [Google Scholar] [CrossRef]
- Bhattacharyya, T.; Gregorova, S.; Mihola, O.; Anger, M.; Sebestova, J.; Denny, P.; Simecek, P.; Forejt, J. Mechanistic basis of infertility of mouse intersubspecific hybrids. PNAS 2013, 110, E468–E477. [Google Scholar] [CrossRef]
- Jan, S.Z.; Jongejan, A.; Korver, C.M.; van Daalen, S.K.; van Pelt, A.M.; Repping, S.; Hamer, G. Distinct prophase arrest mechanisms in human male meiosis. Development 2018, 145. [Google Scholar] [CrossRef]
- Solari, A.J. Sex chromosome pairing and fertility in the heterogametic sex of mammals and birds. In Fertility and Chromosome Pairing: Recent Studies in Plants and Animals; Gillies, C.B., Ed.; CRC Press: Boca Raton, FL, USA, 1989; pp. 77–107. [Google Scholar]
- Spangenberg, V.; Arakelyan, M.; Galoyan, E.; Matveevsky, S.; Petrosyan, R.; Bogdanov, Y.; Danielyan, F.; Kolomiets, O. Reticulate evolution of the rock lizards: Meiotic chromosome dynamics and spermatogenesis in diploid and triploid males of the genus Darevskia. Genes 2017, 8, 149. [Google Scholar] [CrossRef] [PubMed]
- Li, X.C.; Barringer, B.C.; Barbash, D.A. The pachytene checkpoint and its relationship to evolutionary patterns of polyploidization and hybrid sterility. Heredity 2009, 102, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Timoshevskiy, V.A.; Herdy, J.R.; Keinath, M.C.; Smith, J.J. Cellular and molecular features of developmentally programmed genome rearrangement in a vertebrate (sea lamprey: Petromyzon marinus). PLoS Genet. 2016, 12, e1006103. [Google Scholar] [CrossRef] [PubMed]
- Schimenti, J. Synapsis or silence. Nat. Genet. 2005, 37, 11–13. [Google Scholar] [CrossRef] [PubMed]
- Selth, L.A.; Sigurdsson, S.; Svejstrup, J.Q. Transcript elongation by RNA polymerase II. An. Rev. Biochem. 2010, 79, 271–293. [Google Scholar] [CrossRef]
- Eick, D.; Geyer, M. The RNA polymerase II carboxy-terminal domain (CTD) code. Chem. Rev. 2013, 113, 8456–8490. [Google Scholar] [CrossRef]
- Corden, J.L. RNA polymerase II C-terminal domain: Tethering transcription to transcript and template. Chem. Rev. 2013, 113, 8423–8455. [Google Scholar] [CrossRef]
- Matveevsky, S.N.; Pavlova, S.V.; Atsaeva, M.M.; Searle, J.B.; Kolomiets, O.L. Dual mechanism of chromatin remodeling in the common shrew sex trivalent (XY1Y2). Comp. Cytogenet. 2017, 11, 727. [Google Scholar] [CrossRef]
- Gil-Fernández, A.; Matveevsky, S.; Martín-Ruiz, M.; Ribagorda, M.; Parra, M.T.; Viera, A.; Rufas, J.S.; Kolomiets, O.; Bakloushinskaya, I.; Page, J. Sex differences in the meiotic behavior of an XX sex chromosome pair in males and females of the mole vole Ellobius tancrei: Turning an X into a Y chromosome? Chromosoma 2021, 130, 113–131. [Google Scholar] [CrossRef]
- Viera, A.; Parra, M.T.; Rufas, J.S.; Page, J. Transcription reactivation during the first meiotic prophase in bugs is not dependent on synapsis. Chromosoma 2017, 126, 179–194. [Google Scholar] [CrossRef]
- Bae, S.; Lesch, B.J. H3K4me1 distribution predicts transcription state and poising at promoters. Front. Cell Dev. Biol. 2020, 8, 289. [Google Scholar] [CrossRef] [PubMed]
- Prakash, K.; Fournier, D.; Redl, S.; Best, G.; Borsos, M.; Tiwari, V.K.; Tachibana-Konwalski, K.; Ketting, R.F.; Parekh, S.H.; Cremer, C.; et al. Superresolution imaging reveals structurally distinct periodic patterns of chromatin along pachytene chromosomes. Proc. Nat. Acad. Sci. USA 2015, 112, 14635–14640. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matveevsky, S.; Tropin, N.; Kucheryavyy, A.; Kolomiets, O. The First Analysis of Synaptonemal Complexes in Jawless Vertebrates: Chromosome Synapsis and Transcription Reactivation at Meiotic Prophase I in the Lamprey Lampetra fluviatilis (Petromyzontiformes, Cyclostomata). Life 2023, 13, 501. https://doi.org/10.3390/life13020501
Matveevsky S, Tropin N, Kucheryavyy A, Kolomiets O. The First Analysis of Synaptonemal Complexes in Jawless Vertebrates: Chromosome Synapsis and Transcription Reactivation at Meiotic Prophase I in the Lamprey Lampetra fluviatilis (Petromyzontiformes, Cyclostomata). Life. 2023; 13(2):501. https://doi.org/10.3390/life13020501
Chicago/Turabian StyleMatveevsky, Sergey, Nikolay Tropin, Aleksandr Kucheryavyy, and Oxana Kolomiets. 2023. "The First Analysis of Synaptonemal Complexes in Jawless Vertebrates: Chromosome Synapsis and Transcription Reactivation at Meiotic Prophase I in the Lamprey Lampetra fluviatilis (Petromyzontiformes, Cyclostomata)" Life 13, no. 2: 501. https://doi.org/10.3390/life13020501
APA StyleMatveevsky, S., Tropin, N., Kucheryavyy, A., & Kolomiets, O. (2023). The First Analysis of Synaptonemal Complexes in Jawless Vertebrates: Chromosome Synapsis and Transcription Reactivation at Meiotic Prophase I in the Lamprey Lampetra fluviatilis (Petromyzontiformes, Cyclostomata). Life, 13(2), 501. https://doi.org/10.3390/life13020501