
Role of p53 in Regulating Radiation Responses


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
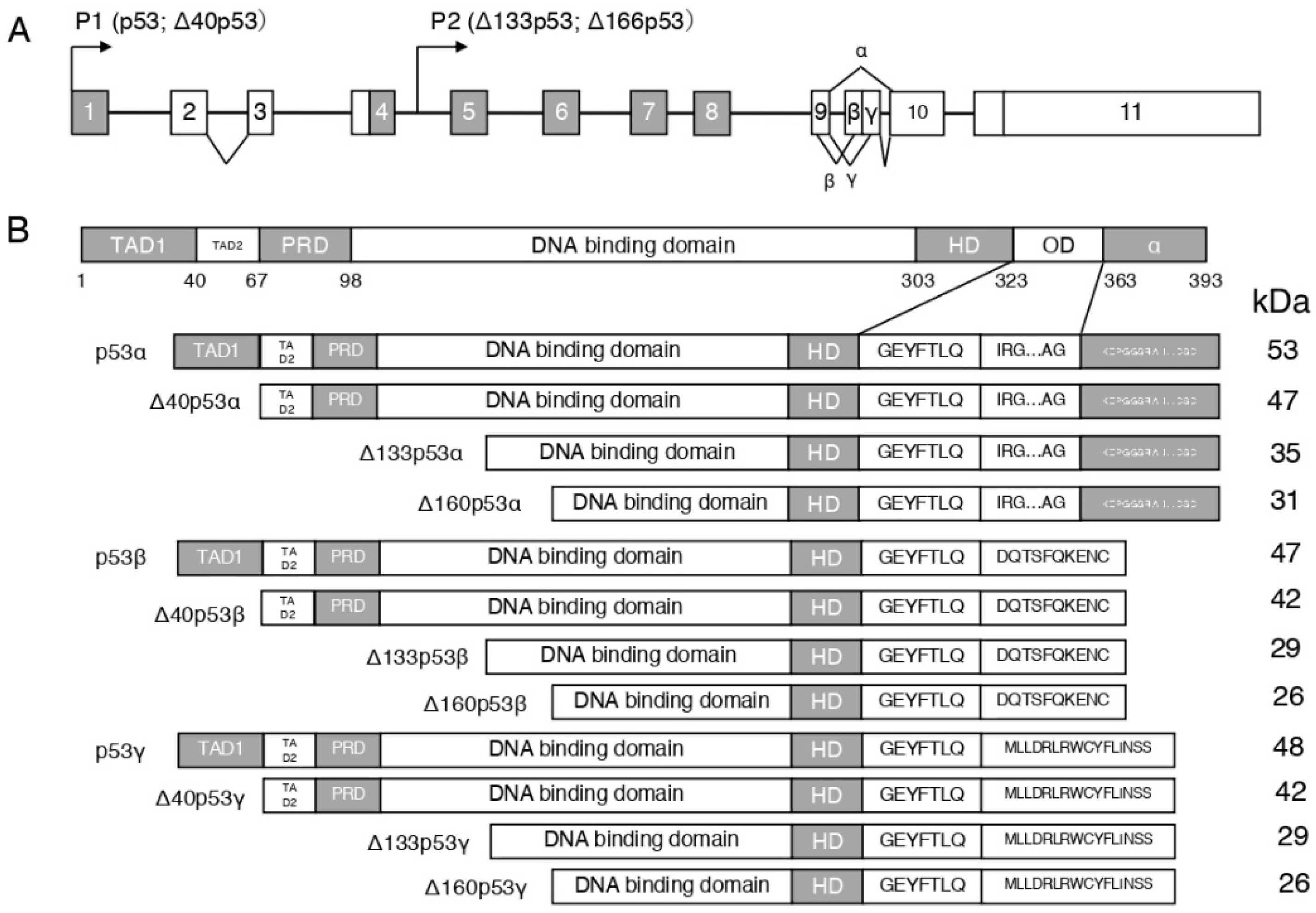
2. p53 Isoforms and Their Responses Response to Radiation
3. The Murine Double Minute 2 (MDM2)-p53 Pathway after Radiation
4. The p53 and Nuclear Factor-KappaB (NF-κB) Pathway after Radiation
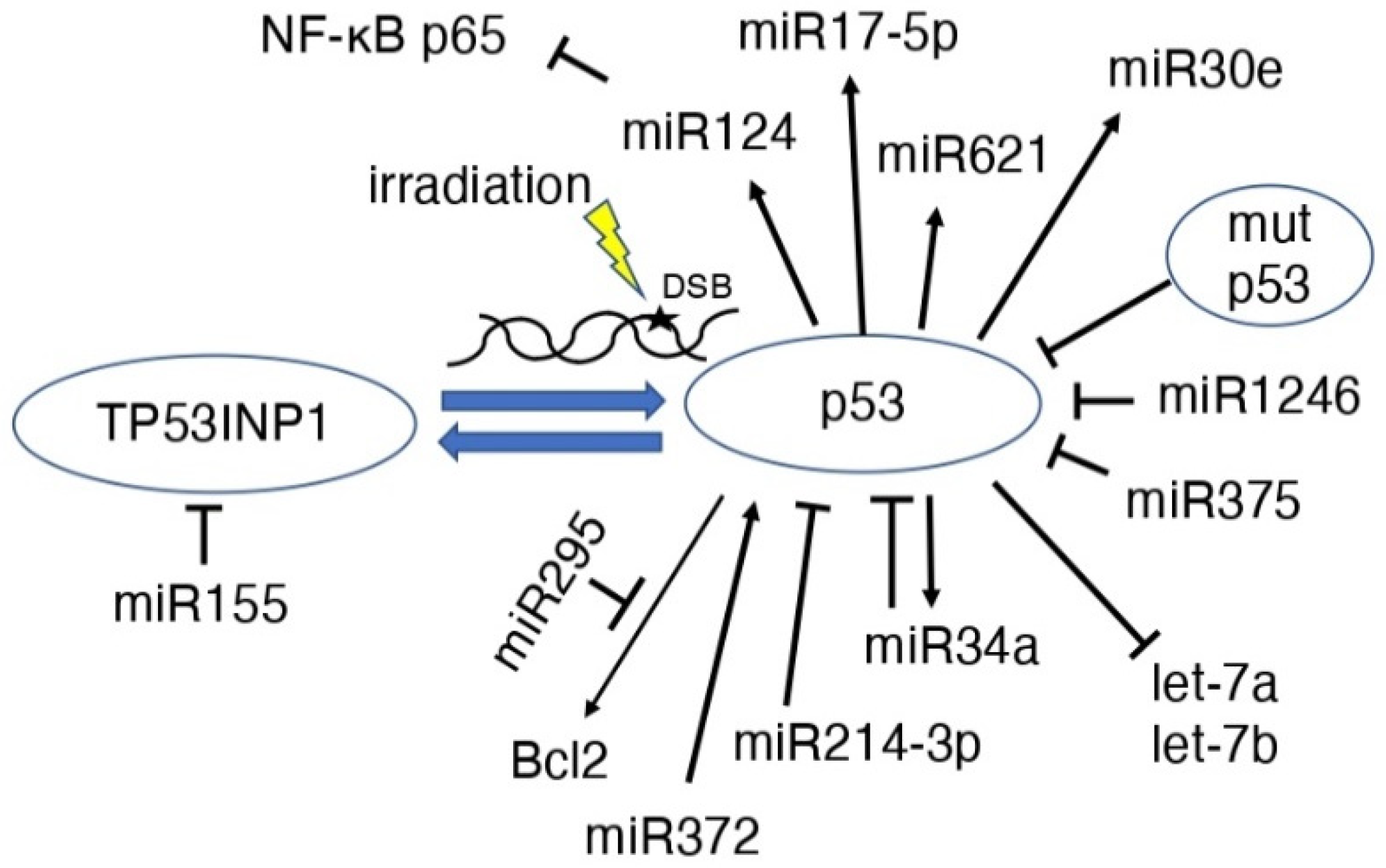
5. p53 and MicroRNAs (miRs) after Radiation
6. p53 and Inflammation Induced by Radiation
7. p53 and Radiation Induced Carcinogenesis
8. p53 and Senescence Induced by Radiation in Normal Cells
9. p53 and Senescence Induced by Radiation in Cancer Cells
10. p53 in the Radioadaptive Response (RAR)
11. p53 as a Biomarker after Irradiation
12. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Linzer, D.; Levine, A.J. Characterization of a 54K Dalton cellular SV40 tumor antigen present in SV40-transformed cells and uninfected embryonal carcinoma cells. Cell 1979, 17, 43–52. [Google Scholar] [CrossRef]
- Kress, M.; May, E.; Cassingena, R.; May, P. Simian virus 40-transformed cells express new species of proteins precipitable by anti-simian virus 40 tumor serum. J. Virol. 1979, 31, 472–483. [Google Scholar] [CrossRef] [PubMed]
- Lane, D.P.; Crawford, L.V. T antigen is bound to a host protein in SY40-transformed cells. Nature 1979, 278, 261–263. [Google Scholar] [CrossRef]
- Givol, D.; Oren, M.; Gruss, P.; Givol, D.; Oren, M. Participation of p53 cellular tumour antigen in transformation of normal embryonic cells. Nature 1984, 312, 646–649. [Google Scholar] [CrossRef]
- Rudge, K.; Jenkins, J.R.; Currie, G.A. Cellular immortalization by a cDNA clone encoding the transformation-associated phosphoprotein p53. Nature 1984, 312, 651–654. [Google Scholar] [CrossRef]
- Weinberg, R.A.; Rotter, V.; Parada, L.F.; Land, H.; Wolf, D. Cooperation between gene encoding p53 tumour antigen and ras in cellular transformation. Nature 1984, 312, 649–651. [Google Scholar] [CrossRef]
- Wolf, D.; Harris, N.; Rotter, V. Reconstitution of p53 expression in a nonproducer Ab-MuLV-transformed cell line by transfection of a functional p53 gene. Cell 1984, 38, 119–126. [Google Scholar] [CrossRef]
- Finlay, C.A.; Hinds, P.W.; Levine, A.J. The p53 proto-oncogene can act as a suppressor of transformation. Cell 1989, 57, 1083–1093. [Google Scholar] [CrossRef]
- Baker, S.J.; Fearon, E.R.; Nigro, J.M.; Hamilton, S.R.; Preisinger, A.C.; Jessup, J.M.; van Tuinen, P.; Ledbetter, D.H.; Barker, D.F.; Nakamura, Y.; et al. Chromosome 17 deletions and p53 gene mutations in colorectal carcinomas. Science 1989, 244, 217–221. [Google Scholar] [CrossRef]
- Lane, D.P. Cancer. p53, guardian of the genome. Nature 1992, 358, 15–16. [Google Scholar] [CrossRef]
- Morgan, S.E.; Kastan, M.B. Foundations in Cancer Research p53 and ATM: Cell Cycle, Cell Death, and Cancer. Adv. Cancer Res. 1997, 71, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Westphal, C.H. Cell-cycle signaling: Atm displays its many talents. Curr. Biol. 1997, 7, R789–R792. [Google Scholar] [CrossRef]
- Rodier, F.; Campisi, J.; Bhaumik, D. Two faces of p53: Aging and tumor suppression. Nucleic Acids Res. 2007, 35, 7475–7484. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Chen, Y.; St Clair, D.K. ROS and p53: A versatile partnership. Free Radic Biol. Med. 2008, 44, 1529–1535. [Google Scholar] [CrossRef] [PubMed]
- Menendez, D.; Inga, A.; Resnick, M.A. The expanding universe of p53 targets. Nat. Rev. Cancer 2009, 9, 724–737. [Google Scholar] [CrossRef]
- Borrás, C.; Gómez-Cabrera, M.C.; Viña, J. The dual role of p53: DNA protection and antioxidant. Free Radic Res. 2011, 45, 643–652. [Google Scholar] [CrossRef]
- Cheung, E.C.; Vousden, K.H. The role of p53 in glucose metabolism. Curr. Opin. Cell Biol. 2010, 22, 186–191. [Google Scholar] [CrossRef]
- Olovnikov, I.A.; Kravchenko, J.E.; Chumakov, P.M. Homeostatic functions of the p53 tumor suppressor: Regulation of energy metabolism and antioxidant defense. Semin. Cancer Biol. 2008, 19, 32–41. [Google Scholar] [CrossRef]
- Agoff, S.N.; Hou, J.; Linzer, D.I.; Wu, B. Regulation of the human hsp70 promoter by p53. Science 1993, 259, 84–87. [Google Scholar] [CrossRef]
- Hussain, S.P.; Amstad, P.; He, P.; Robles, A.; Lupold, S.; Kaneko, I.; Ichimiya, M.; Sengupta, S.; Mechanic, L.; Okamura, S.; et al. p53-Induced Up-Regulation of MnSOD and GPx but not Catalase Increases Oxidative Stress and Apoptosis. Cancer Res. 2004, 64, 2350–2356. [Google Scholar] [CrossRef]
- MacCallum, D.E.; Hall, P.A.; Wright, E.G. The Trp53 Pathway Is Induced In Vivo by Low Doses of Gamma Radiation. Radiat. Res. 2001, 156, 324–327. [Google Scholar] [CrossRef]
- Ghosh, J.C.; Suzuki, K.; Kodama, S.; Watanabe, M. Effects of Protein Kinase Inhibitors on the Accumulation Kinetics of p53 Protein in Normal Human Embryo Cells following X-irradiation. J. Radiat. Res. 1999, 40, 23–37. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Aloni-Grinstein, R.; Schwartz, D.; Rotter, V. Accumulation of wild-type p53 protein upon gamma-irradiation induces a G2 arrest-dependent immunoglobulin kappa light chain gene expression. EMBO J. 1995, 14, 1392–1401. [Google Scholar] [CrossRef]
- Fritsche, M.; Haessler, C.; Brandner, G. Induction of nuclear accumulation of the tumor-suppressor protein p53 by DNA-damaging agents. Oncogene 1993, 8, 307–318. [Google Scholar] [PubMed]
- Kim, S.; An, S.S. Role of p53 isoforms and aggregations in cancer. Medicine 2016, 95, e3993. [Google Scholar] [CrossRef] [PubMed]
- Joruiz, S.M.; Bourdon, J. p53 Isoforms: Key Regulators of the Cell Fate Decision. Cold Spring Harb. Perspect. Med. 2016, 6, a026039. [Google Scholar] [CrossRef] [PubMed]
- Joerger, A.C.; Fersht, A.R. Structural Biology of the Tumor Suppressor p53. Annu. Rev. Biochem. 2008, 77, 557–582. [Google Scholar] [CrossRef]
- Osada, M.; Ohba, M.; Kawahara, C.; Ishioka, C.; Kanamaru, R.; Katoh, I.; Ikawa, Y.; Nimura, Y.; Nakagawara, A.; Obinata, M.; et al. Cloning and functional analysis of human p51, which structurally and functionally resembles p53. Nat. Med. 1998, 4, 839–843. [Google Scholar] [CrossRef]
- Kaghad, M.; Bonnet, H.; Yang, A.; Creancier, L.; Biscan, J.C.; Valent, A.; Minty, A.; Chalon, P.; Lelias, J.M.; Dumont, X.; et al. Monoallelically Expressed Gene Related to p53 at 1p36, a Region Frequently Deleted in Neuroblastoma and Other Human Cancers. Cell 1997, 90, 809–819. [Google Scholar] [CrossRef]
- Jun, C.; Sok, M.N.; Changqing, C.; Zhenhai, Z.; Bourdon, J.C.; Lane, D.P.; Peng, J. p53 isoform Δ113p53 is a p53 target gene that antagonizes p53 apoptotic activity via BclxL activation in zebrafish. Genes Dev. 2009, 23, 278–290. [Google Scholar]
- Turnquist, C.; Beck, J.A.; Horikawa, I. Radiation-induced astrocyte senescence is rescued by Δ133p53. Neuro Oncol. 2019, 21, 474–485. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Nguyen, T.; Lee, E.M.; Castro-Aceituno, V.; Wagle, R.; Lee, K.S.; Choi, J.; Song, Y.H. Role of p53 isoforms in the DNA damage response during Drosophila oogenesis. Sci. Rep. 2019, 9, 11473. [Google Scholar] [CrossRef] [PubMed]
- Coutandin, D.; Osterburg, C.; Srivastav, R.K.; Sumyk, M.; Kehrloesser, S.; Gebel, J.; Tuppi, M.; Hannewald, J.; Schäfer, B.; Salah, E.; et al. Quality control in oocytes by p63 is based on a spring-loaded activation mechanism on the molecular and cellular level. Elife 2016, 5, e13909. [Google Scholar] [CrossRef] [PubMed]
- Krolewski, B.; Little, J. Alterations of mdm2 gene in x-ray transformed mouse 10t1/2 cell clones. Int. J. Oncol. 1995, 6, 1123–1127. [Google Scholar] [CrossRef] [PubMed]
- Amundson, S.A.; Lee, R.A.; Koch-Paiz, C.A.; Bittner, M.L.; Meltzer, P.; Trent, J.M.; Fornace, A.J., Jr. Differential Responses of Stress Genes to Low Dose-Rate γ Irradiation. Mol. Cancer Res. 2003, 1, 445. [Google Scholar]
- Terzian, T.; Wang, Y.; Van Pelt, C.S.; Box, N.F.; Travis, E.L.; Lozano, G. Haploinsufficiency of Mdm2 and Mdm4 in Tumorigenesis and Development. Mol. Cell. Biol. 2007, 27, 5479–5485. [Google Scholar] [CrossRef]
- Jabbur, J.R.; Tabor, A.D.; Cheng, X.; Wang, H.; Uesugi, M.; Lozano, G.; Zhang, W. Mdm-2 binding and TAFII31 recruitment is regulated by hydrogen bond disruption between the p53 residues Thr18 and Asp21. Oncogene 2002, 21, 7100–7113. [Google Scholar] [CrossRef]
- Xiong, X.; Zhao, Y.; He, H.; Sun, Y. Ribosomal protein S27-like and S27 interplay with p53-MDM2 axis s a target, a substrate, and a regulator. Oncogene 2010, 30, 1798–1811. [Google Scholar] [CrossRef]
- Martínez, A.; Brethauer, U.; Borlando, J.; Spencer, M.L.; Rojas, I.G. Epithelial expression of p53, mdm-2 and p21 in normal lip and actinic cheilitis. Oral Oncol. 2007, 44, 878–883. [Google Scholar] [CrossRef]
- Beskow, C.; Skikuniene, J.; Holgersson, A.; Nilsson, B.; Lewensohn, R.; Kanter, L.; Viktorsson, K. Radioresistant cervical cancer shows upregulation of the NHEJ proteins DNA-PKcs, Ku70 and Ku86. Br. J. Cancer 2009, 101, 816–821. [Google Scholar] [CrossRef]
- Magné, N.; Toillon, R.A.; Bottero, V.; Didelot, C.; Houtte, P.V.; Gérard, J.P.; Peyron, J.F. NF-κB modulation and ionizing radiation: Mechanisms and future directions for cancer treatment. Cancer Lett. 2006, 231, 158–168. [Google Scholar] [CrossRef]
- Habraken, Y.; Piette, J. NF-κB activation by double-strand breaks. Biochem. Pharmacol. 2006, 72, 1132–1141. [Google Scholar] [CrossRef] [PubMed]
- Kawai, H.; Yamada, Y.; Tatsuka, M.; Niwa, O.; Yamamoto, K.; Suzuki, F. Down-regulation of nuclear factor kappaB is required for p53-dependent apoptosis in X-ray-irradiated mouse lymphoma cells and thymocytes. Cancer Res. 1999, 59, 6038–6041. [Google Scholar] [PubMed]
- Jonak, K.; Kurpas, M.; Szoltysek, K.; Janus, P.; Abramowicz, A.; Puszynski, K. A novel mathematical model of ATM/p53/NF- κB pathways points to the importance of the DDR switch-off mechanisms. BMC Syst. Biol. 2016, 10, 75. [Google Scholar] [CrossRef]
- Xufeng, C.; Binghui, S.; Liqun, X.; Khaletzkiy, A.; Chu, D.; Wong, J.Y.; Li, J.J. Activation of nuclear factor κb in radioresistance of TP53-inactive human keratinocytes. Cancer Res. 2002, 62, 1213–1221. [Google Scholar]
- Baust, H.; Schoke, A.; Brey, A.; Gern, U.; Los, M.; Schmid, R.M.; Röttinger, E.M.; Seufferlein, T. Evidence for radiosensitizing by gliotoxin in HL-60 cells: Implications for a role of NF-κB independent mechanisms. Oncogene 2003, 22, 8786–8796. [Google Scholar] [CrossRef][Green Version]
- van Hogerlinden, M.; Auer, G.; Toftgard, R. Inhibition of Rel/Nuclear Factor-[kappa]B signaling in skin results in defective DNA damage-induced cell cycle arrest and Ha-ras- and p53-independent tumor development. Oncogene 2002, 21, 4969. [Google Scholar] [CrossRef][Green Version]
- Wang, H.; Segaran, R.C.; Chan, L.Y.; Aladresi, A.A.M.; Chinnathambi, A.; Alharbi, S.A.; Sethi, G.; Tang, F.R. Gamma Radiation-Induced Disruption of Cellular Junctions in HUVECs Is Mediated through Affecting MAPK/NF-κB Inflammatory Pathways. Oxid. Med. Cell Longev. 2019, 2019, 1486232. [Google Scholar] [CrossRef]
- Janssens, S.; Tinel, A.; Lippens, S.; Tschopp, J. PIDD mediates NF-kappaB activation in response to DNA damage. Cell 2005, 123, 1079–1092. [Google Scholar] [CrossRef]
- Zając, G.; Marek, R.; Barbara, Ł.S.; Krzysztof, P.; Piotr, W. Activation of the atypical NF-κB pathway induced by ionizing radiation is not affected by the p53 status. Acta Biochim. Pol. 2022, 69, 205–210. [Google Scholar] [CrossRef]
- Zhu, Y.; Zuo, W.; Shen, X.; Liu, Y.; Zhao, Y.; Xiong, Y.; Cao, H.; Wang, Y.; Liang, Z. NF-κB is involved in the regulation of autophagy in mutant p53 cells in response to ionizing radiation. Cell Death. Discov. 2021, 7, 159. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Okazaki, R.; Yoshida, Y. Senescence-associated secretory phenotype and activation of NF-κB in splenocytes of old mice exposed to irradiation at a young age. Dev. Comp. Immunol. 2021, 122, 104124. [Google Scholar] [CrossRef] [PubMed]
- Wellenstein, M.D.; Coffelt, S.B.; Duits, D.E.M.; van Miltenburg, M.H.; Slagter, M.; de Rink, I.; Henneman, L.; Kas, S.M.; Prekovic, S.; Hau, C.S. Loss of p53 triggers WNT-dependent systemic inflammation to drive breast cancer metastasis. Nature 2019, 572, 538–542. [Google Scholar] [CrossRef]
- Gasparian, A.V.; Burkhart, C.A.; Purmal, A.A.; Brodsky, L.; Pal, M.; Saranadasa, M.; Bosykh, D.A.; Commane, M.; Guryanova, O.A.; Pal, S.; et al. Curaxins: Anticancer compounds that simultaneously suppress NF-κB and activate p53 by targeting FACT. Sci. Transl. Med. 2011, 3, 95ra74. [Google Scholar] [CrossRef] [PubMed]
- Gurova, K.V.; Hill, J.E.; Guo, C.; Prokvolit, A.; Burdelya, L.G.; Samoylova, E.; Khodyakova, A.V.; Ganapathi, R.; Ganapathi, M.; Tararova, N.D.; et al. Small Molecules That Reactivate p53 in Renal Cell Carcinoma Reveal a NF-κB-Dependent Mechanism of p53 Suppression in Tumors. Proc. Natl. Acad. Sci. USA 2005, 102, 17448–17453. [Google Scholar] [CrossRef] [PubMed]
- Gudkov, A.V.; Komarova, E.A. p53 and the Carcinogenicity of Chronic Inflammation. Cold Spring Harb. Perspect. Med. 2016, 6, a026161. [Google Scholar] [CrossRef]
- Puszynski, K.; Bertolusso, R.; Lipniacki, T. Crosstalk between p53 and nuclear factor-B systems: Pro- and anti-apoptotic functions of NF-B. IET Syst. Biol. 2009, 3, 356–367. [Google Scholar] [CrossRef]
- Adams, B.D.; Parsons, C.; Walker, L.; Zhang, W.C.; Slack, F.J. Targeting noncoding RNAs in disease. J. Clin. Investig. 2017, 127, 761–771. [Google Scholar] [CrossRef]
- Bajan, S.; Hutvagner, G. Regulation of miRNA Processing and miRNA Mediated Gene Repression in Cancer. Microrna 2014, 3, 10–17. [Google Scholar] [CrossRef]
- Bajan, S.; Hutvagner, G. RNA-Based Therapeutics: From Antisense Oligonucleotides to miRNAs. Cells 2020, 9, 137. [Google Scholar] [CrossRef]
- Zenz, T.; Mohr, J.; Eldering, E.; Kater, A.P.; Bühler, A.; Kienle, D.; Winkler, D.; Dürig, J.; van Oers, M.H.; Mertens, D.; et al. miR-34a as part of the resistance network in chronic lymphocytic leukemia. Blood 2009, 113, 3801–3808. [Google Scholar] [CrossRef] [PubMed]
- Hermeking, H. p53 Enters the MicroRNA World. Cancer Cell 2007, 12, 414–418. [Google Scholar] [CrossRef] [PubMed]
- Balca-Silva, J.; Sousa Neves, S.; Goncalves, C.; Abrantes, A.M.; Casalta-Lopes, J.; Botelho, M.F.; Sarmento-Ribeiro, A.B.; Silva, H.C. Effect of miR-34b Overexpression on the Radiosensitivity of Non-small Cell Lung Cancer Cell Lines. Anticancer Res. 2012, 32, 1603–1609. [Google Scholar] [PubMed]
- Liu, C.; Zhou, C.; Gao, F.; Cai, S.; Zhang, C.; Zhao, L.; Zhao, F.; Cao, F.; Lin, J.; Yang, Y.; et al. MiR-34a in Age and Tissue Related Radio-Sensitivity and Serum miR-34a as a Novel Indicator of Radiation Injury. Int. J. Biol. Sci. 2011, 7, 221–233. [Google Scholar] [CrossRef]
- Mert, U.; Ozgür, E.; Tiryakioglu, D.; Dalay, N.; Gezer, U. Induction of p53-inducible microRNA miR-34 by gamma radiation and bleomycin are different. Front. Genet. 2012, 3, 220. [Google Scholar] [CrossRef]
- Sasaki, A.; Udaka, Y.; Tsunoda, Y.; Yamamoto, G.; Tsuji, M.; Oyamada, H.; Oguchi, K.; Mizutani, T. Analysis of p53 and miRNA Expression after Irradiation of Glioblastoma Cell Lines. Anticancer Res. 2012, 32, 4709–4713. [Google Scholar]
- Wang, B.; Li, D.; Kovalchuk, O. p53 Ser15 phosphorylation and histone modifications contribute to IR-induced miR-34a transcription in mammary epithelial cells. Cell Cycle 2013, 12, 2073–2083. [Google Scholar] [CrossRef]
- Saleh, A.D.; Savage, J.E.; Cao, L.; Soule, B.P.; Ly, D.; DeGraff, W.; Harris, C.C.; Mitchell, J.B.; Simone, N.L. Cellular stress induced alterations in microRNA let-7a and let-7b expression are dependent on p53. PLoS ONE 2011, 6, e24429. [Google Scholar] [CrossRef]
- John-Aryankalayil, M.; Palayoor, S.T.; Makinde, A.Y.; Cerna, D.; Simone, C.B.; Falduto, M.T.; Magnuson, S.R.; Coleman, C.N. Fractionated Radiation Alters Oncomir and Tumor Suppressor miRNAs in Human Prostate Cancer Cells. Radiat. Res. 2012, 178, 105–117. [Google Scholar] [CrossRef]
- Zheng, G.X.; Ravi, A.; Calabrese, J.M.; Medeiros, L.A.; Kirak, O.; Dennis, L.M.; Jaenisch, R.; Burge, C.B.; Sharp, P.A. A Latent Pro-Survival Function for the Mir-290-295 Cluster in Mouse Embryonic Stem Cells. PLoS Genet. 2011, 7, e1002054. [Google Scholar] [CrossRef]
- Wang, Y.; Scheiber, M.N.; Neumann, C.; Calin, G.A.; Zhou, D. MicroRNA Regulation of Ionizing Radiation-Induced Premature Senescence. Int. J. Radiat. Oncol. Biol. Phys. 2011, 81, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xing, R.; Zhang, X.; Dong, W.; Zhang, J.; Yan, Z.; Li, W.; Cui, J.; Lu, Y. miR-375 targets the p53 gene to regulate cellular response to ionizing radiation and etoposide in gastric cancer cells. DNA Repair. 2013, 12, 741–750. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.; Kim, J.; Nam, J.; Sun, H.; Lee, Y.H.; Lee, T.J.; Aguiar, R.C.; Kim, S.W. MicroRNA-124 links p53 to the NF-κB pathway in B-cell lymphomas. Leukemia 2015, 29, 1868–1874. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.Y.; Wu, A.T.; Liu, S. MicroRNA-17-5p regulated apoptosis-related protein expression and radiosensitivity in oral squamous cell carcinoma caused by betel nut chewing. Oncotarget 2016, 7, 51482–51493. [Google Scholar] [CrossRef]
- Wang, Z.; Mao, J.; Liu, G.; Wang, F.G.; Ju, Z.S.; Zhou, D.; Wang, R.Y. MicroRNA-372 enhances radiosensitivity while inhibiting cell invasion and metastasis in nasopharyngeal carcinoma through activating the PBK-dependent p53 signaling pathway. Cancer Med. 2019, 8, 712–728. [Google Scholar] [CrossRef]
- Shao, Y.; Song, X.; Jiang, W.; Chen, Y.; Ning, Z.; Gu, W.; Jiang, J. MicroRNA-621 Acts as a Tumor Radiosensitizer by Directly Targeting SETDB1 in Hepatocellular Carcinoma. Mol. Ther. 2019, 27, 355–364. [Google Scholar] [CrossRef]
- Xu, R.; Li, H.; Wu, S.; Qu, J.; Yuan, H.; Zhou, Y.; Lu, Q. MicroRNA-1246 regulates the radio-sensitizing effect of curcumin in bladder cancer cells via activating P53. Int. Urol. Nephrol. 2019, 51, 1771–1779. [Google Scholar] [CrossRef]
- Sohn, D.; Peters, D.; Piekorz, R.P.; Budach, W.; Jänicke, R.U. miR-30e controls DNA damage-induced stress responses by modulating expression of the CDK inhibitor p21WAF1/CIP1 and caspase-3. Oncotarget 2016, 7, 15915–15929. [Google Scholar] [CrossRef]
- Lei, X.; He, N.; Zhu, L.; Zhou, M.; Zhang, K.; Wang, C.; Huang, H.; Chen, S.; Li, Y.; Liu, Q.; et al. Mesenchymal Stem Cell-Derived Extracellular Vesicles Attenuate Radiation-Induced Lung Injury via miRNA-214-3p. Antioxid. Redox Signal. 2021, 35, 849–862. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, C.; Zhao, Y.; Feng, Z. MicroRNA Control of p53. J. Cell. Biochem. 2017, 118, 7–14. [Google Scholar] [CrossRef]
- Candéias, S.M.; Testard, I. The many interactions between the innate immune system and the response to radiation. Cancer Lett. 2015, 368, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, T.; Takaoka, A.; Hayakawa, S.; Yanai, H.; Stoiber, D.; Negishi, H.; Kikuchi, H.; Sasaki, S.; Imai, K.; Shibue, T.; et al. Integration of interferon-α/β signalling to p53 responses in tumour suppression and antiviral defence. Nature 2003, 424, 516–523. [Google Scholar] [CrossRef]
- Muñoz-Fontela, C.; Macip, S.; Martínez-Sobrido, L.; Brown, L.; Ashour, J.; García-Sastre, A.; Lee, S.W.; Aaronson, S.A. Transcriptional role of p53 in interferon-mediated antiviral immunity. J. Exp. Med. 2008, 205, 1929–1938. [Google Scholar] [CrossRef] [PubMed]
- Dharel, N.; Kato, N.; Muroyama, R.; Taniguchi, H.; Otsuka, M.; Wang, Y.; Jazag, A.; Shao, R.X.; Chang, J.H.; Adler, M.K.; et al. Potential contribution of tumor suppressor p53 in the host defense against hepatitis C virus. Hepatology 2008, 47, 1136–1149. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, D.; Coates, P.J.; Lorimore, S.A.; Wright, E.G. The in vivo expression of radiation-induced chromosomal instability has an inflammatory mechanism. Radiat. Res. 2012, 177, 18–24. [Google Scholar] [CrossRef]
- Coates, P.; Robinson, J.; Lorimore, S.A.; Wright, E.G. Ongoing activation of p53 pathway responses is a long-term consequence of radiation exposure in vivo and associates with altered macrophage activities. J. Pathol. 2008, 214, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Coates, P.J.; Rundle, J.K.; Lorimore, S.A.; Wright, E.G. Indirect Macrophage Responses to Ionizing Radiation: Implications for Genotype-Dependent Bystander Signaling. Cancer Res. 2008, 68, 450–456. [Google Scholar] [CrossRef]
- Lorimore, S.A.; Chrystal, J.A.; Robinson, J.I.; Coates, P.J.; Wright, E.G. Chromosomal Instability in Unirradiated Hemaopoietic Cells Induced by Macrophages Exposed In vivo to Ionizing Radiation. Cancer Res. 2008, 68, 8122–8126. [Google Scholar] [CrossRef]
- Lorimore, S.A.; Mukherjee, D.; Robinson, J.I.; Chrystal, J.A.; Wright, E.G. Long-lived inflammatory signaling in irradiated bone marrow is genome dependent. Cancer Res. 2011, 71, 6485–6491. [Google Scholar] [CrossRef]
- McAllister, K.A.; Lorimore, S.A.; Wright, E.G.; Coates, P.J. In Vivo Interactions between Ionizing Radiation, Inflammation and Chemical Carcinogens Identified by Increased DNA Damage Responses. Radiat. Res. 2012, 177, 584–593. [Google Scholar] [CrossRef]
- Futaki, N.; Takahashi, S.; Yokoyama, M.; Arai, I.; Higuchi, S.; Otomo, S. NS-398, a new anti-inflammatory agent, selectively inhibits prostaglandin G/H synthase/cyclooxygenase (COX-2) activity in vitro. Prostaglandins 1994, 47, 55–59. [Google Scholar] [CrossRef]
- Mukherjee, D.; Coates, P.J.; Rastogi, S.; Lorimore, S.A.; Wright, E.G. Radiation-induced bone marrow apoptosis, inflammatory bystander-type signaling and tissue cytotoxicity. Int. J. Radiat. Biol. 2013, 89, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Frank, A.K.; Leu, J.J.; Zhou, Y.; Devarajan, K.; Nedelko, T.; Klein-Szanto, A.; Hollstein, M.; Murphy, M.E. The Codon 72 Polymorphism of p53 Regulates Interaction with NF-κB and Transactivation of Genes Involved in Immunity and Inflammation. Mol. Cell. Biol. 2011, 31, 1201–1213. [Google Scholar] [CrossRef]
- Lorimore, S.A.; Coates, P.J.; Scobie, G.E.; Milne, G.; Wright, E.G. Inflammatory-type responses after exposure to ionizing radiation in vivo: A mechanism for radiation-induced bystander effects. Oncogene 2001, 20, 7085–7095. [Google Scholar] [CrossRef]
- Merched, A.J.; Chan, L. Absence of p21Waf1/Cip1/Sdi1 Modulates Macrophage Differentiation and Inflammatory Response and Protects Against Atherosclerosis. Circulation 2004, 110, 3830–3841. [Google Scholar] [CrossRef] [PubMed]
- Derradji, H.; Bekaert, S.; De Meyer, T.; Jacquet, P.; Abou-El-Ardat, K.; Ghardi, M.; Arlette, M.; Baatout, S. Ionizing radiation-induced gene modulations, cytokine content changes and telomere shortening in mouse fetuses exhibiting forelimb defects. Dev. Biol. 2008, 322, 302–313. [Google Scholar] [CrossRef]
- Toutounchian, J.J.; Steinle, J.J.; Makena, P.S.; Waters, C.M.; Wilson, M.W.; Haik, B.G.; Miller, D.D.; Yates, C.R. Modulation of radiation injury response in retinal endothelial cells by quinic acid derivative KZ-41 involves p38 MAPK. PLoS ONE 2014, 9, e100210. [Google Scholar] [CrossRef]
- Dong, X.; Luo, M.; Huang, G.; Zhang, J.; Tong, F.; Cheng, Y.; Cai, Q.; Dong, J.; Wu, G.; Cheng, J. Relationship between irradiation-induced neuro-inflammatory environments and impaired cognitive function in the developing brain of mice. Int. J. Radiat. Biol. 2015, 91, 224–239. [Google Scholar] [CrossRef]
- Huang, B.; Zhang, Q.; Yuan, Y.; Xin, N.; He, K.; Huang, Y.; Tang, H.; Gong, P. Sema3a inhibits the differentiation of Raw264.7 cells to osteoclasts under 2Gy radiation by reducing inflammation. PLoS ONE 2018, 13, e0200000. [Google Scholar] [CrossRef]
- Ham, S.W.; Jeon, H.Y.; Jin, X.; Kim, E.J.; Kim, J.K.; Shin, Y.J.; Lee, Y.; Kim, S.H.; Lee, S.Y.; Seo, S.; et al. TP53 gain-of-function mutation promotes inflammation in glioblastoma. Cell Death Differ. 2019, 26, 409–425. [Google Scholar] [CrossRef]
- Madhugiri, V.S.; Moiyadi, A.V.; Shetty, P.; Gupta, T.; Epari, S.; Jalali, R.; Subeikshanan, V.; Dutt, A.; Sasidharan, G.M.; Roopesh Kumar, V.R.; et al. Analysis of Factors Associated with Long-Term Survival in Patients with Glioblastoma. World. Neurosurg. 2021, 149, e758–e765. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, L.; Bi, N.; Ji, W.; Tan, W.; Zhao, L.; Yu, D.; Wu, C.; Wang, L.; Lin, D. Association of P53 and ATM polymorphisms with risk of radiation-Induced pneumonitis in lung cancer patients treated with radiotherapy. Int. J. Radiat. Oncol. Biol. Phys. 2011, 79, 1402–1407. [Google Scholar] [CrossRef]
- Rogers, C.J.; Kyubwa, E.M.; Lukaszewicz, A.I.; Yamada-Hanff, J.; Starbird, M.A.; Miller, T.A.; Phelps, A.A.; Wallack, S.; Mahendra, S.; Thrall, K.; et al. Identification of miRNA Associated with Reduced Survival after Whole-Thorax Lung Irradiation in Non-Human Primates. Radiat. Res. 2021, 196, 510–522. [Google Scholar] [CrossRef] [PubMed]
- Ju, Z.; Shen, L.; Zhou, M.; Luo, J.; Yu, Z.; Qu, C.; Lei, R.; Lei, M.; Huang, R. Helicobacter pylori and Alzheimer’s Disease-Related Metabolic Dysfunction: Activation of TLR4/Myd88 Inflammation Pathway from p53 Perspective and a Case Study of Low-Dose Radiation Intervention. ACS. Chem. Neurosci. 2022, 13, 1065–1081. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, N. Reexamining the role of tissue inflammation in radiation carcinogenesis: A hypothesis to explain an earlier onset of cancer. Int. J. Radiat. Biol. 2021, 97, 1341–1351. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.W.; Karin, M.A. cytokine-mediated link between innate immunity, inflammation, and cancer. J. Clin. Investig. 2007, 117, 1175–1183. [Google Scholar] [CrossRef]
- Coussens, L.M.; de Visser, K.E.; Eichten, A. Paradoxical roles of the immune system during cancer development. Nat. Rev. Cancer 2006, 6, 24–37. [Google Scholar] [CrossRef]
- Balkwill, F.; Coussens, L.M. Cancer: An inflammatory link. Nature 2004, 431, 405–406. [Google Scholar] [CrossRef]
- Coussens, L.M.; Zena, W. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef]
- Nguyen, D.H.; Fredlund, E.; Zhao, W.; Perou, C.M.; Balmain, A.; Mao, J.H.; Barcellos-Hoff, M.H. Murine microenvironment metaprofiles associate with human cancer etiology and intrinsic subtypes. Clin. Cancer Res. 2013, 19, 1353–1362. [Google Scholar] [CrossRef]
- Morioka, T.; Miyoshi-Imamura, T.; Blyth, B.J.; Kaminishi, M.; Kokubo, T.; Nishimura, M.; Kito, S.; Tokairin, Y.; Tani, S.; Murakami-Murofushi, K.; et al. Ionizing radiation, inflammation, and their interactions in colon carcinogenesis in Mlh1-deficient mice. Cancer Sci. 2015, 106, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Manna, K.; Das, U.; Das, D.; Kaminishi, M.; Kokubo, T.; Nishimura, M.; Kito, S.; Tokairin, Y.; Tani, S.; Murakami-Murofushi, K.; et al. Naringin inhibits gamma radiation-induced oxidative DNA damage and inflammation, by modulating p53 and NF-κB signaling pathways in murine splenocytes. Cancer Sci. 2015, 49, 422–439. [Google Scholar] [CrossRef] [PubMed]
- Keam, S.P.; Caramia, F.; Gamell, C.; Paul, P.J.; Arnau, G.M.; Neeson, P.J.; Williams, S.G.; Haupt, Y. The Transcriptional Landscape of Radiation-Treated Human Prostate Cancer: Analysis of a Prospective Tissue Cohort. Int. J. Radiat. Oncol. Biol. Phys. 2018, 100, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Kobatake, K.; Ikeda, K.; Nakata, Y.; Yamasaki, N.; Ueda, T.; Kanai, A.; Sentani, K.; Sera, Y.; Hayashi, T.; Koizumi, M.; et al. Kdm6a Deficiency Activates Inflammatory Pathways, Promotes M2 Macrophage Polarization, and Causes Bladder Cancer in Cooperation with p53 Dysfunction. Clin. Cancer Res. 2020, 26, 2065–2079. [Google Scholar] [CrossRef]
- Brachman, D.G.; Hallahan, D.E.; Beckett, M.A.; Yandell, D.W.; Weichselbaum, R.R. p53 gene mutations and abnormal retinoblastoma protein in radiation-induced human sarcomas. Cancer Res. 1991, 51, 6393–6396. [Google Scholar]
- Lee, J.M.; Abrahamson, J.L.; Kandel, R.; Donehower, L.A.; Bernstein, A. Susceptibility to radiation-carcinogenesis and accumulation of chromosomal breakage in p53 deficient mice. Oncogene 1994, 9, 3731–3736. [Google Scholar]
- Iwamoto, K.S.; Fujii, S.; Kurata, A.; Suzuki, M.; Hayashi, T.; Ohtsuki, Y.; Okada, Y.; Narita, M.; Takahashi, M.; Hosobe, S.; et al. p53 mutations in tumor and non-tumor tissues of Thorotrast recipients: A model for cellular selection during radiation carcinogenesis in the liver. Carcinogenesis 1999, 20, 1283–1291. [Google Scholar] [CrossRef][Green Version]
- Wiese, C.; Gauny, S.S.; Liu, W.C.; Cherbonnel-Lasserre, C.L.; Kronenberg, A. Different Mechanisms of Radiation-induced Loss of Heterozygosity in Two Human Lymphoid Cell Lines from a Single Donor. Cancer Res. 2001, 61, 1129–1137. [Google Scholar]
- Gonin-Laurent, N.; Gibaud, A.; Huygue, M.; Lefèvre, S.H.; Le Bras, M.; Chauveinc, L.; Sastre-Garau, X.; Doz, F.; Lumbroso, L.; Chevillard, S.; et al. Specific TP53 mutation pattern in radiation-induced sarcomas. Carcinogenesis 2006, 27, 1266–1272. [Google Scholar] [CrossRef]
- Po, Z.; De-Wen, W.; Guomin, L.; Yabing, G.; Xiaoying, L. Abnormal location of p16 protein and overexpression of p53 protein in human radiation-induced skin cancer. J. Environ. Pathol. Toxicol. Oncol. 1995, 14, 25–28. [Google Scholar]
- Selvanayagam, C.S.; Davis, C.M.; Cornforth, M.N.; Ullrich, R.L. Latent Expression of p53 Mutations and Radiation-induced Mammary Cancer. Cancer Res. 1995, 55, 3310–3317. [Google Scholar] [PubMed]
- Igari, K.; Igari, Y.; Okazaki, R.; Kato, F.; Ootsuyama, A.; Norimura, T. The Delayed Manifestation of T-Cell Receptor (TCR) Variants in X-Irradiated Mice Depends on Trp53 Status. Radiat. Res. 2006, 166, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Mendonca, M.S.; Mayhugh, B.M.; McDowell, B.; Chin-Sinex, H.; Smith, M.L.; Dynlacht, J.R.; Spandau, D.F.; Lewis, D.A. A Radiation-Induced Acute Apoptosis Involving TP53 and BAX Precedes the Delayed Apoptosis and Neoplastic Transformation of CGL1 Human Hybrid Cells. Radiat. Res. 2005, 163, 614–622. [Google Scholar] [CrossRef]
- Hollstein, M.; Bartsch, H.; Wesch, H.; Kure, E.H.; Mustonen, R.; Mühlbauer, K.R.; Spiethoff, A.; Wegener, K.; Wiethege, T.; Müller, K.M. p53 gene mutation analysis in tumors of patients exposed to alpha-particles. Carcinogenesis 1997, 18, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.P.; Kennedy, C.H.; Amstad, P.; Lui, H.; Lechner, J.F.; Harris, C.C. Radon and lung carcinogenesis: Mutability of p53 codons 249 and 250 to 238Pu alpha-particles in human bronchial epithelial cells. Carcinogenesis 1997, 18, 121–125. [Google Scholar] [CrossRef]
- Andersson, M.; Jönsson, M.; Nielsen, L.L.; Vyberg, M.; Visfeldt, J.; Storm, H.H.; Wallin, H. Mutations in the tumor suppressor gene p53 in human liver cancer induced by alpha-particles. Cancer Epidemiol. Biomark. Prev. 1995, 4, 765–770. [Google Scholar]
- Jin, Y.; Burns, F.J.; Garte, S.J.; Hosselet, S. Infrequent alterations of the p53 gene in rat skin cancers induced by ionizing radiation. Carcinogenesis 1996, 17, 873–876. [Google Scholar] [CrossRef]
- Rabes, H.M. Gene rearrangements in radiation-induced thyroid carcinogenesis. Med. Pediatr. Oncol. 2001, 36, 574–582. [Google Scholar] [CrossRef]
- Suchy, B.; Waldmann, V.; Klugbauer, S.; Rabes, H.M. Absence of RAS and p53 mutations in thyroid carcinomas of children after Chernobyl in contrast to adult thyroid tumours. Br. J. Cancer. 1998, 77, 952–955. [Google Scholar] [CrossRef]
- Hillebrandt, S.; Streffer, C.; Demidchik, E.P.; Biko, J.; Reiners, C. Polymorphisms in the p53 gene in thyroid tumours and blood samples of children from areas in Belarus. Mutat. Res. 1997, 381, 201–207. [Google Scholar] [CrossRef]
- Han, Y.; Tan, Y.; Zhao, Y.; Zhang, Y.; He, X.; Yu, L.; Jiang, H.; Lu, H.; Tian, H. TRIM23 overexpression is a poor prognostic factor and contributes to carcinogenesis in colorectal cancer. J. Cell. Mol. Med. 2020, 24, 5491–5500. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Huang, Y.; Wang, Z.; Li, X.; Louie, A.; Wei, G.; Mao, J.H. Temporal mTOR inhibition protects Fbxw7-deficient mice from radiation-induced tumor development. Aging 2013, 5, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.L.; Castle, K.D.; Moding, E.J.; Blum, J.M.; Williams, N.; Luo, L.; Ma, Y.; Borst, L.B.; Kim, Y.; Kirsch, D.G. Acute DNA damage activates the tumour suppressor p53 to promote radiation-induced lymphoma. Nat. Commun. 2015, 6, 8477. [Google Scholar] [CrossRef]
- Moding, E.J.; Min, H.D.; Castle, K.D.; Ali, M.; Woodlief, L.; Williams, N.; Ma, Y.; Kim, Y.; Lee, C.L.; Kirsch, D.G. An extra copy of p53 suppresses development of spontaneous Kras-driven but not radiation-induced cancer. CI Insight 2016, 1, e86698. [Google Scholar] [CrossRef] [PubMed]
- Carr, M.I.; Roderick, J.E.; Gannon, H.S.; Kelliher, M.A.; Jones, S.N. Mdm2 Phosphorylation Regulates Its Stability and Has Contrasting Effects on Oncogene and Radiation-Induced Tumorigenesis. Cell Rep. 2016, 16, 2618–2629. [Google Scholar] [CrossRef]
- Chibaya, L.; Karim, B.; Zhang, H.; Jones, S.N. Mdm2 phosphorylation by Akt regulates the p53 response to oxidative stress to promote cell proliferation and tumorigenesis. Proc. Natl. Acad. Sci. USA 2021, 118, e2003193118. [Google Scholar] [CrossRef]
- Lee, C.L.; Brock, K.D.; Hasapis, S.; Zhang, D.; Sibley, A.B.; Qin, X.; Gresham, J.S.; Caraballo, I.; Luo, L.; Daniel, A.R.; et al. Whole-Exome Sequencing of Radiation-Induced Thymic Lymphoma in Mouse Models Identifies Notch1 Activation as a Driver of p53 Wild-Type Lymphoma. Cancer Res. 2021, 81, 3777–3790. [Google Scholar] [CrossRef]
- McHugh, D.; Gil, J. Senescence and aging: Causes, consequences, and therapeutic avenues. J. Cell Biol. 2018, 217, 65–77. [Google Scholar] [CrossRef]
- Salama, R.; Sadaie, M.; Hoare, M.; Narita, M. Cellular senescence and its effector programs. Genes Dev. 2014, 28, 99–114. [Google Scholar] [CrossRef]
- Kruiswijk, F.; Labuschagne, C.F.; Vousden, K.H. p53 in survival, death and metabolic health: A lifeguard with a licence to kill. Nat. Rev. Mol. Cell. Biol. 2015, 16, 393–405. [Google Scholar] [CrossRef]
- Baker, D.J.; Childs, B.G.; Durik, M.; Wijers, M.E.; Sieben, C.J.; Zhong, J.; Saltness, R.A.; Jeganathan, K.B.; Verzosa, G.C.; Pezeshki, A.; et al. Naturally occurring p16Ink4a-positive cells shorten healthy lifespan. Nature 2016, 530, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Bartoletti-Stella, A.; Mariani, E.; Kurelac, I.; Maresca, A.; Caratozzolo, M.F.; Iommarini, L.; Carelli, V.; Eusebi, L.H.; Guido, A.; Cenacchi, G.; et al. Gamma rays induce a p53-independent mitochondrial biogenesis that is counter-regulated by HIF1α. Cell Death Dis. 2013, 4, e663. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Wang, Y.; Wang, J.; Zhai, J.; He, F.; Zhu, G. Irradiation-induced senescence of bone marrow mesenchymal stem cells aggravates osteogenic differentiation dysfunction via paracrine signaling. Am. J. Physiol. Cell Physiol. 2020, 318, C1005–C1017. [Google Scholar] [CrossRef]
- Suzuki, K.; Mori, I.; Nakayama, Y.; Miyakoda, M.; Kodama, S.; Watanabe, M. Radiation-Induced Senescence-like Growth Arrest Requires TP53 Function but not Telomere Shortening. Radiat. Res. 2001, 155, 248–253. [Google Scholar] [CrossRef]
- Le, O.N.; Rodier, F.; Fontaine, F.; Coppe, J.P.; Campisi, J.; DeGregori, J.; Laverdiere, C.; Kokta, V.; Haddad, E.; Beausejour, C.M. Ionizing radiation-induced long-term expression of senescence markers in mice is independent of p53 and immune status. Aging Cell 2010, 9, 398–409. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Suzuki, K.; Kodama, S.; Watanabe, M. Interstitial chromatin alteration causes persistent p53 activation involved in the radiation-induced senescence-like growth arrest. Biochem. Biophys. Res. Commun. 2006, 340, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.R.; Elmore, L.W.; Jackson-Cook, C.; Demasters, G.; Povirk, L.F.; Holt, S.E.; Gewirtz, D.A. p53-Dependent accelerated senescence induced by ionizing radiation in breast tumour cells. Int. J. Radiat. Biol. 2005, 81, 445–458. [Google Scholar] [CrossRef]
- Sharma, G.G.; Hall, E.J.; Dhar, S.; Gupta, A.; Rao, P.H.; Pandita, T.K. Telomere stability correlates with longevity of human beings exposed to ionizing radiations. Oncol. Rep. 2003, 10, 1733–1736. [Google Scholar] [CrossRef]
- Meng, A.; Wang, Y.; Van Zant, G.; Zhou, D. Ionizing Radiation and Busulfan Induce Premature Senescence in Murine Bone Marrow Hematopoietic Cells. Cancer Res 2003, 63, 5414–5419. [Google Scholar]
- Yentrapalli, R.; Azimzadeh, O.; Barjaktarovic, Z.; Sarioglu, H.; Wojcik, A.; Harms-Ringdahl, M.; Atkinson, M.J.; Haghdoost, S.; Tapio, S. Quantitative proteomic analysis reveals induction of premature senescence in human umbilical vein endothelial cells exposed to chronic low-dose rate gamma radiation. Proteomics 2013, 13, 1096–1107. [Google Scholar] [CrossRef]
- Dong, X.; Tong, F.; Qian, C.; Zhang, R.; Dong, J.; Wu, G.; Hu, Y. NEMO Modulates Radiation-Induced Endothelial Senescence of Human Umbilical Veins Through NF-κB Signal Pathway. Radiat. Res. 2015, 183, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, E.C.; Dimaio, D. Induced senescence in HeLa cervical carcinoma cells containing elevated telomerase activity and extended telomeres. Cell Growth Differ. 2001, 12, 525–534. [Google Scholar] [PubMed]
- Itjima, M.; Mihara, K.; Kondo, T.; Tsuji, T.; Ishioka, C.; Namba, M. Mutation in p53 and de-regulation of p53-related gene expression in three human cell lines immortalized with 4-nitroquinoline 1-oxide or 60Co gamma rays. Int. J. Cancer 1996, 66, 698–702. [Google Scholar] [CrossRef]
- Zheng, Q.H.; Ma, L.W.; Zhu, W.G.; Zhang, Z.Y.; Tong, T.J. p21Waf1/Cip1 plays a critical role in modulating senescence through changes of DNA methylation. J. Cell. Biochem. 2006, 98, 1230–1248. [Google Scholar] [CrossRef] [PubMed]
- Lim, D.S.; Vogel, H.; Willerford, D.M.; Sands, A.T.; Platt, K.A.; Hasty, P. Analysis of ku80-Mutant Mice and Cells with Deficient Levels of p53. Mol. Cell. Biol. 2000, 20, 3772–3780. [Google Scholar] [CrossRef]
- Naka, K.; Tachibana, A.; Ikeda, K.; Motoyama, N. Stress-induced Premature Senescence in hTERT-expressing Ataxia Telangiectasia Fibroblasts. J. Biol. Chem. 2004, 279, 2030–2037. [Google Scholar] [CrossRef]
- MacLaren, A.; Black, E.J.; Clark, W.; Gillespie, D.A. c-Jun-Deficient Cells Undergo Premature Senescence as a Result of Spontaneous DNA Damage Accumulation. Mol. Cell. Biol. 2004, 24, 9006–9018. [Google Scholar] [CrossRef]
- Wang, D.; Jang, D.J. Protein Kinase CK2 Regulates Cytoskeletal Reorganization during Ionizing Radiation-Induced Senescence of Human Mesenchymal Stem Cells. Cancer Res. 2009, 69, 8200–8207. [Google Scholar] [CrossRef]
- Calabrese, V.; Mallette, F.A.; Deschênes-Simard, X.; Ramanathan, S.; Gagnon, J.; Moores, A.; Ilangumaran, S.; Ferbeyre, G. SOCS1 Links Cytokine Signaling to p53 and Senescence. Mol. Cell 2009, 36, 754–767. [Google Scholar] [CrossRef]
- Cho, S.J.; Rossi, A.; Jung, Y.S.; Yan, W.; Liu, G.; Zhang, J.; Zhang, M.; Chen, X. Ninjurin1, a target of p53, regulates p53 expression and p53-dependent cell survival, senescence, and radiation-induced mortality. Proc. Natl. Acad. Sci. USA 2013, 110, 9362–9367. [Google Scholar] [CrossRef]
- Ronald Allan, M.P.; Regina, M.D. Inhibition of IGF-1R prevents ionizing radiation-induced primary endothelial cell senescence. PLoS ONE 2013, 8, e78589. [Google Scholar] [CrossRef]
- Neise, D.; Sohn, D.; Stefanski, A.; Goto, H.; Inagaki, M.; Wesselborg, S.; Budach, W.; Stühler, K.; Jänicke, R.U. The p90 ribosomal S6 kinase (RSK) inhibitor BI-D1870 prevents gamma irradiation-induced apoptosis and mediates senescence via RSK- and p53-independent accumulation of p21WAF1/CIP1. Cell Death Dis. 2013, 4, e859. [Google Scholar] [CrossRef]
- Wang, Y.; Blandino, G.; Givol, D. Induced p21waf expression in H1299 cell line promotes cell senescence and protects against cytotoxic effect of radiation and doxorubicin. Oncogene 1999, 18, 2643–2649. [Google Scholar] [CrossRef]
- Liakou, E.; Mavrogonatou, E.; Pratsinis, H.; Rizou, S.; Evangelou, K.; Panagiotou, P.N.; Karamanos, N.K.; Gorgoulis, V.G.; Kletsas, D. Ionizing radiation-mediated premature senescence and paracrine interactions with cancer cells enhance the expression of syndecan 1 in human breast stromal fibroblasts: The role of TGF-beta. Aging 2016, 8, 1650–1669. [Google Scholar] [CrossRef] [PubMed]
- Byun, H.S.; Cho, E.W.; Kim, J.S.; Moon, M.S.; Yum, J.J.; Kim, K.C.; Kim, I.G. Thioredoxin overexpression in HT-1080 cells induced cellular senescence and sensitization to gamma radiation. FEBS Lett. 2005, 579, 4055–4062. [Google Scholar] [CrossRef][Green Version]
- Quick, Q.A.; Gewirtz, D.A. An accelerated senescence response to radiation in wild-type p53 glioblastoma multiforme cells. J. Neurosurg. 2006, 105, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Toillon, R.A.; Magne, N.; Laios, I.; Castadot, P.; Kinnaert, E.; Van Houtte, P.; Desmedt, C.; Leclercq, G.; Lacroix, M. Estrogens decrease [gamma]-ray-induced senescence and maintain cell cycle progression in breast cancer cells independently of p53. Int. J. Radiat. Oncol. Biol. Phys. 2007, 67, 1187. [Google Scholar] [CrossRef]
- Karimi-Busheri, F.; Rasouli-Nia, A.; Mackey, J.R.; Weinfeld, M. Senescence evasion by MCF-7 human breast tumor-initiating cells. Breast Cancer Res. 2010, 12, R31. [Google Scholar] [CrossRef]
- Shin, J.S.; Sang, H.W.; Jin, D.H.; Park, I.C.; Lee, H.C.; Hong, S.W.; Yoo, D.H.; Hong, S.I.; Lee, W.J.; Lee, M.S.; et al. Low doses of ionizing radiation suppress doxorubicin-induced senescence-like phenotypes by activation of ERK1/2 and suppression of p38 kinase in MCF7 human breast cancer cells. Int. J. Oncol. 2010, 36, 1445–1452. [Google Scholar] [CrossRef]
- Lehmann, B.D.; McCubrey, J.A.; Jefferson, H.S.; Paine, M.S.; Chappell, W.H.; Terrian, D.M. A Dominant Role for p53-Dependent Cellular Senescence in Radiosensitization of Human Prostate Cancer Cells. Cell Cycle 2007, 6, 595–605. [Google Scholar] [CrossRef]
- Arya, A.K.; El-Fert, A.; Jones, T.M.; Boyd, M.T.; Devling, T.; Eccles, R.M.; Aslam, M.A.; Rubbi, C.P.; Vlatkovic, N.; Fenwick, J.; et al. Nutlin-3, the small-molecule inhibitor of MDM2, promotes senescence and radiosensitises laryngeal carcinoma cells harbouring wild-type p53. Br. J. Cancer 2010, 103, 186–195. [Google Scholar] [CrossRef]
- Luo, H.; Yount, C.; Lang, H.; Yang, A.; Riemer, E.C.; Lyons, K.; Vanek, K.N.; Silvestri, G.A.; Schulte, B.A.; Wang, G.Y. Activation of p53 with Nutlin-3a radiosensitizes lung cancer cells via enhancing radiation-induced premature senescence. Lung Cancer 2013, 81, 167–173. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Guo, J.; Song, H.; Zhu, H.; Di, X.; Min, H.; Wang, Y.; Chen, G.; Dai, W.; Ma, J.; et al. Nutlin-3, an Antagonist of MDM2, Enhances the Radiosensitivity of Esophageal Squamous Cancer with Wild-Type p53. Pathol. Oncol. Res. 2017, 24, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Efimova, E.V.; Mauceri, H.J.; Kron, S.J.; Weichselbaum, R.R.; Golden, D.W.; Labay, E.; BindokasI, V.P.; Darga, T.E.; Chakraborty, C.; Barreto-Andrade, J.C.; et al. Poly(ADP-ribose) polymerase inhibitor induces accelerated senescence in irradiated breast cancer cells and tumors. Cancer Res. 2010, 70, 6277–6282. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulou, A.; Kletsas, D. Human lung fibroblasts prematurely senescent after exposure to ionizing radiation enhance the growth of malignant lung epithelial cells in vitro and in vivo. Int. J. Oncol. 2011, 39, 989–999. [Google Scholar] [CrossRef] [PubMed]
- Azad, A.; Jackson, S.; Cullinane, C.; Natoli, A.; Neilsen, P.M.; Callen, D.F.; Maira, S.M.; Hackl, W.; McArthur, G.A.; Solomon, B.; et al. Inhibition of DNA-dependent protein kinase induces accelerated senescence in irradiated human cancer cells. Mol. Cancer Res. 2011, 9, 1696–1707. [Google Scholar] [CrossRef]
- Li, X.H.; Ha, C.T.; Fu, D.; Xiao, M. REDD1 protects osteoblast cells from gamma radiation-induced premature senescence. PLoS ONE 2012, 7, e36604. [Google Scholar] [CrossRef]
- Guo, H.; Liu, Z.; Xu, B.; Hu, H.; Wei, Z.; Liu, Q.; Zhang, X.; Ding, X.; Wang, Y.; Zhao, M.; et al. Chemokine receptor CXCR2 is transactivated by p53 and induces p38-mediated cellular senescence in response to DNA damage. Aging Cell 2013, 12, 1110–1121. [Google Scholar] [CrossRef]
- Feys, L.; Descamps, B.; Vanhove, C.; Vral, A.; Veldeman, L.; Vermeulen, S.; De Wagter, C.; Bracke, M.; De Wever, O. Radiation-induced lung damage promotes breast cancer lung-metastasis through CXCR4 signaling. Oncotarget 2015, 6, 26615–26632. [Google Scholar] [CrossRef]
- Skwarska, A.; Ramachandran, S.; Dobrynin, G.; Leszczynska, K.B.; Hammond, E.M. The imidazoacridinone C-1311 induces p53-dependent senescence or p53-independent apoptosis and sensitizes cancer cells to radiation. Oncotarget 2017, 8, 31187–31198. [Google Scholar] [CrossRef]
- Tesei, A.; Arienti, C.; Bossi, G.; Santi, S.; De Santis, I.; Bevilacqua, A.; Zanoni, M.; Pignatta, S.; Cortesi, M.; Zamagni, A.; et al. TP53 drives abscopal effect by secretion of senescence-associated molecular signals in non-small cell lung cancer. J. Exp. Clin. Cancer Res. 2021, 40, 89. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, G.; Bodycote, J.; Wolff, S. Adaptive response of human lymphocytes to low concentrations of radioactive thymidine. Science 1984, 223, 594–597. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, A.; Ohnishi, K.; Asakawa, I.; Kondo, N.; Nakagawa, H.; Yonezawa, M.; Tachibana, A.; Matsumoto, H.; Ohnishi, T. Radiation response of apoptosis in C57BL/6N mouse spleen after whole-body irradiation. Int. J. Radiat. Biol. 2001, 77, 939–945. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, R.; Ootsuyama, A.; Norimura, T. TP53 and TP53-Related Genes Associated with Protection from Apoptosis in the Radioadaptive Response. Radiat. Res. 2007, 167, 51–57. [Google Scholar] [CrossRef]
- Horie, K.; Kubo, K.; Yonezawa, M. p53 Dependency of Radio-adaptive Responses in Endogenous Spleen Colonies and Peripheral Blood-cell Counts in C57BL Mice. Comp. Study 2002, 43, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Wang, B. Involvement of p53-Dependent Apoptosis in Radiation Teratogenesis and in the Radioadaptive Response in the Late Organogenesis of Mice. J. Radiat. Res. 2001, 42, 1–10. [Google Scholar] [CrossRef]
- Ahmed, K.M.; Nantajit, D.; Fan, M.; Murley, J.S.; Grdina, D.J.; Li, J.J. Coactivation of ATM/ERK/NF-κB in the low-dose radiation-induced radioadaptive response in human skin keratinocytes. Free Radic Biol. Med. 2009, 46, 1543–1550. [Google Scholar] [CrossRef]
- Murley, J.S.; Miller, R.C.; Weichselbaum, R.R.; Grdina, D.J. TP53 Mutational Status and ROS Effect the Expression of the Survivin-Associated Radio-Adaptive Response. Radiat. Res. 2017, 188, 659–670. [Google Scholar] [CrossRef]
- Shadley, J.D. Chromosomal Adaptive Response in Human Lymphocytes. Radiat. Res. 1994, 138, S9–S12. [Google Scholar] [CrossRef]
- Sasaki, M.S. On the Reaction Kinetics of the Radioadaptive Response in Cultured Mouse Cells. Int. J. Radiat. Biol. 1995, 68, 281–291. [Google Scholar] [CrossRef]
- Takahashi, A.; Kondo, N.; Inaba, H.; Uotani, K.; Kiyohara, Y.; Ohnishi, K.; Ohnishi, T. Radiation-induced apoptosis in SCID mice spleen after low dose irradiation. Adv. Space Res. 2003, 31, 1569–1573. [Google Scholar] [CrossRef]
- Wang, B.; Ohyama, H.; Shang, Y.; Tanaka, K.; Aizawa, S.; Yukawa, O.; Hayata, I. Adaptive Response in Embryogenesis: V. Existence of Two Efficient Dose-Rate Ranges for 0.3 Gy of Priming Irradiation to Adapt Mouse Fetuses. Radiat. Res. 2004, 161, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Mitchel, R.E.; Jackson, J.S.; McCann, R.A.; Boreham, D.R. The Adaptive Response Modifies Latency for Radiation-Induced Myeloid Leukemia in CBA/H Mice. Radiat. Res. 1999, 152, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Vares, G.; Wang, B.; Shang, Y.; Ohyama, H.; Tanaka, K.; Nakajima, T.; Nenoi, M.; Hayata, I. Adaptive response in embryogenesis: VI. Comparative microarray analysis of gene expressions in mouse fetuses. Int. J. Radiat. Biol. 2009, 85, 70–86. [Google Scholar] [CrossRef]
- Wang, B.; Ohyama, H.; Shang, Y.; Fujita, K.; Tanaka, K.; Nakajima, T.; Aizawa, S.; Yukawa, O.; Hayata, I. Adaptive Response in Embryogenesis: IV. Protective and Detrimental Bystander Effects Induced by X Radiation in Cultured Limb Bud Cells of Fetal Mice. Radiat. Res. 2004, 161, 9–16. [Google Scholar] [CrossRef]
- Wang, B.; Ohyama, H.; Haginoya, K.; Odaka, T.; Itsukaichi, H.; Yukawa, O.; Yamada, T.; Hayata, I. Adaptive Response in Embryogenesis. III. Relationship to Radiation-Induced Apoptosis and Trp53 Gene Status. Radiat. Res. 2000, 154, 277–282. [Google Scholar] [CrossRef]
- Wang, B.; Ohyama, H.; Haginoya, K.; Odaka, T.; Itsukaichi, H.; Nose, M.; Nakajima, T.; Yukawa, O.; Yamada, T.; Hayata, I. Adaptive Response in Embryogenesis: II.Retardation of Postnatal Development of Prenatally Irradiated Mice. Radiat. Res. 1999, 152, 119–123. [Google Scholar] [CrossRef]
- Wang, B.; Ohyama, H.; Nose, T.; Itsukaichi, H.; Nakajima, T.; Yukawa, O.; Odaka, T.; Tanaka, K.; Kojima, E.; Yamada, T.; et al. Adaptive Response in Embryogenesis: I. Dose and Timing of Radiation for Reduction of Prenatal Death and Congenital Malformation during the Late Period of Organogenesis. Radiat. Res. 1998, 150, 120–122. [Google Scholar] [CrossRef]
- Okazaki, R.; Ootsuyama, A.; Norimura, T. Radioadaptive Response for Protection against Radiation-Induced Teratogenesis. Radiat. Res. 2005, 163, 266–270. [Google Scholar] [CrossRef]
- Boreham, D.R.; Dolling, J.A.; Somers, C.; Quinn, J.; Mitchel, R.E. The Adaptive Response and Protection against Heritable Mutations and Fetal Malformation. Dose Response 2006, 4, 317–326. [Google Scholar] [CrossRef]
- Jiang, H.; Li, W.; Li, X.; Cai, L.; Wang, G. Low-dose radiation induces adaptive response in normal cells, but not in tumor cells: In vitro and in vivo studies. J. Radiat. Res 2008, 49, 219–230. [Google Scholar] [CrossRef] [PubMed]
- López-Nieva, P.; Malavé, M.; González-Sánchez, L.; Fernández-Piqueras, J.; Fernández-Navarro, P.; Santos, J. Transcriptomic analysis reveals sex-specific differences in the expression of Dcl1 and Fis1 genes in the radio-adaptive response of thymocytes to TRP53-mediated apoptosis. BMC Genom. 2016, 17, 698. [Google Scholar] [CrossRef] [PubMed]
- Varès, G.; Wang, B.; Tanaka, K.; Kakimoto, A.; Eguchi-Kasai, K.; Nenoi, M. Mutagenic adaptive response to high-LET radiation in human lymphoblastoid cells exposed to low doses of heavy-ion radiation. Mutat. Res. 2011, 712, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Jain, V.; Das, B. Global transcriptome profile reveals abundance of DNA damage response and repair genes in individuals from high level natural radiation areas of Kerala coast. PLoS ONE 2017, 12, e0187274. [Google Scholar] [CrossRef] [PubMed]
- Rajabi, P.M.; Fardid, R.; Zare, T.; Kargar, S.F.; Moslen-Shirazi, M.A.; Behzad, B.A. Assessment of Adaptive Response of Gamma Radiation in the Operating Room Personnel Exposed to Anesthetic Gases by Measuring the Relative Gene Expression Changes Ku80, Ligase1 and P53. J. Biomed. Phys Eng. 2020, 10, 225–234. [Google Scholar] [CrossRef][Green Version]
- Guéguen, Y.; Bontemps, A.; Ebrahimian, T.G. Adaptive responses to low doses of radiation or chemicals: Their cellular and molecular mechanisms. Cell. Mol. Life Sci. 2018, 76, 1255–1273. [Google Scholar] [CrossRef]
- Schlichtholz, B.; Trédaniel, J.; Lubin, R.; Zalcman, G.; Hirsch, A.; Soussi, T. Analyses of p53 antibodies in sera of patients with lung carcinoma define immunodominant regions in the p53 protein. Br. J. Cancer 1994, 69, 809–816. [Google Scholar] [CrossRef][Green Version]
- Luo, J.C.; Zehab, R.; Anttila, S.; Ridanpaa, M.; Husgafvel-Pursiainen, K.; Vainio, H.; Carney, W.; De Vivo, I.; Milling, C.; Brandt-Rauf, P.W. Detection of Serum p53 Protein in Lung Cancer Patients. J. Occup. Med. 1994, 36, 155–160. [Google Scholar] [CrossRef]
- Lubin, R.; Zalcman, G.; Bouchet, L.; Tredanel, J.; Legros, Y.; Cazals, D.; Hirsch, A.; Soussi, T. Serum p53 antibodies as early markers of lung cancer. Nat. Med. 1995, 1, 701–702. [Google Scholar] [CrossRef]
- Brandt-Rauf, P.W.; Smith, S.; Hemminki, K.; Koskinen, H.; Vainio, H.; Niman, H.; Ford, J. Serum oncoproteins and growth factors in asbestosis and silicosis patients. Int. J. Cancer 1992, 50, 881–885. [Google Scholar] [CrossRef]
- Huang, R.; Liu, X.; Li, H.; Zhou, Y.; Zhou, P.K. Integrated analysis of transcriptomic and metabolomic profiling reveal the p53 associated pathways underlying the response to ionizing radiation in HBE cells. Cell. Biosci. 2020, 10, 56. [Google Scholar] [CrossRef] [PubMed]
- Popp, W.; Plappert, U.; Müller, W.; Rehn, B.; Schneider, J.; Braun, A.; Bauer, P.C.; Vahrenholz, C.; Presek, P.; Brauksiepe, A.; et al. Biomarkers of genetic damage and inflammation in blood and bronchoalveolar lavage fluid among former German uranium miners: A pilot study. Radiat. Environ. Biophys. 2000, 39, 275–282. [Google Scholar] [CrossRef] [PubMed]
- El-Saghire, H.; Thierens, H.; Monsieurs, P.; Michaux, A.; Vandevoorde, C.; Baatout, S. Gene set enrichment analysis highlights different gene expression profiles in whole blood samples X-irradiated with low and high doses. Int. J. Radiat. Biol. 2013, 89, 628–638. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhou, S.; Jia, M.; Li, X.; Li, L.; Wang, Q.; Qi, Z.; Zhou, P.; Li, Y.; Wang, Z. Early Biomarkers Associated with P53 Signaling for Acute Radiation Injury. Life 2022, 12, 99. [Google Scholar] [CrossRef]
- Hayashi, T.; Morishita, Y.; Khattree, R.; Misumi, M.; Sasaki, K.; Hayashi, I.; Yoshida, K.; Kajimura, J.; Kyoizumi, S.; Imai, K.; et al. Evaluation of systemic markers of inflammation in atomic-bomb survivors with special reference to radiation and age effects. FASEB J. 2012, 26, 4765–4773. [Google Scholar] [CrossRef]
- Hayashi, T.; Kusunoki, Y.; Hakoda, M.; Morishita, Y.; Kubo, Y.; Maki, M.; Kasagi, F.; Kodama, K.; Macphee, D.G.; Kyoizumi, S. Radiation dose-dependent increases in inflammatory response markers in A-bomb survivors. Int. J. Radiat. Biol. 2003, 79, 129–136. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okazaki, R. Role of p53 in Regulating Radiation Responses. Life 2022, 12, 1099. https://doi.org/10.3390/life12071099
Okazaki R. Role of p53 in Regulating Radiation Responses. Life. 2022; 12(7):1099. https://doi.org/10.3390/life12071099
Chicago/Turabian StyleOkazaki, Ryuji. 2022. "Role of p53 in Regulating Radiation Responses" Life 12, no. 7: 1099. https://doi.org/10.3390/life12071099
APA StyleOkazaki, R. (2022). Role of p53 in Regulating Radiation Responses. Life, 12(7), 1099. https://doi.org/10.3390/life12071099