
Ameliorative Effect of a Neoteric Regimen of Catechin plus Cetirizine on Ovalbumin-Induced Allergic Rhinitis in Rats

 ,
,  ,
,  , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals and Experimental Protocol
2.3. Vascular Permeability Assay
2.4. Determination of Histamine Level and HDC Activity
2.5. Barbiturate-Induced Sleeping Time
2.6. Molecular Docking
2.6.1. Compounds Retrieval and Preparation
2.6.2. HDC Structure Retrieval, Preparation, and Docking
2.6.3. Binding Energy Calculations
2.6.4. Binding-Energy Decomposition
2.7. Statistical Analysis
3. Results
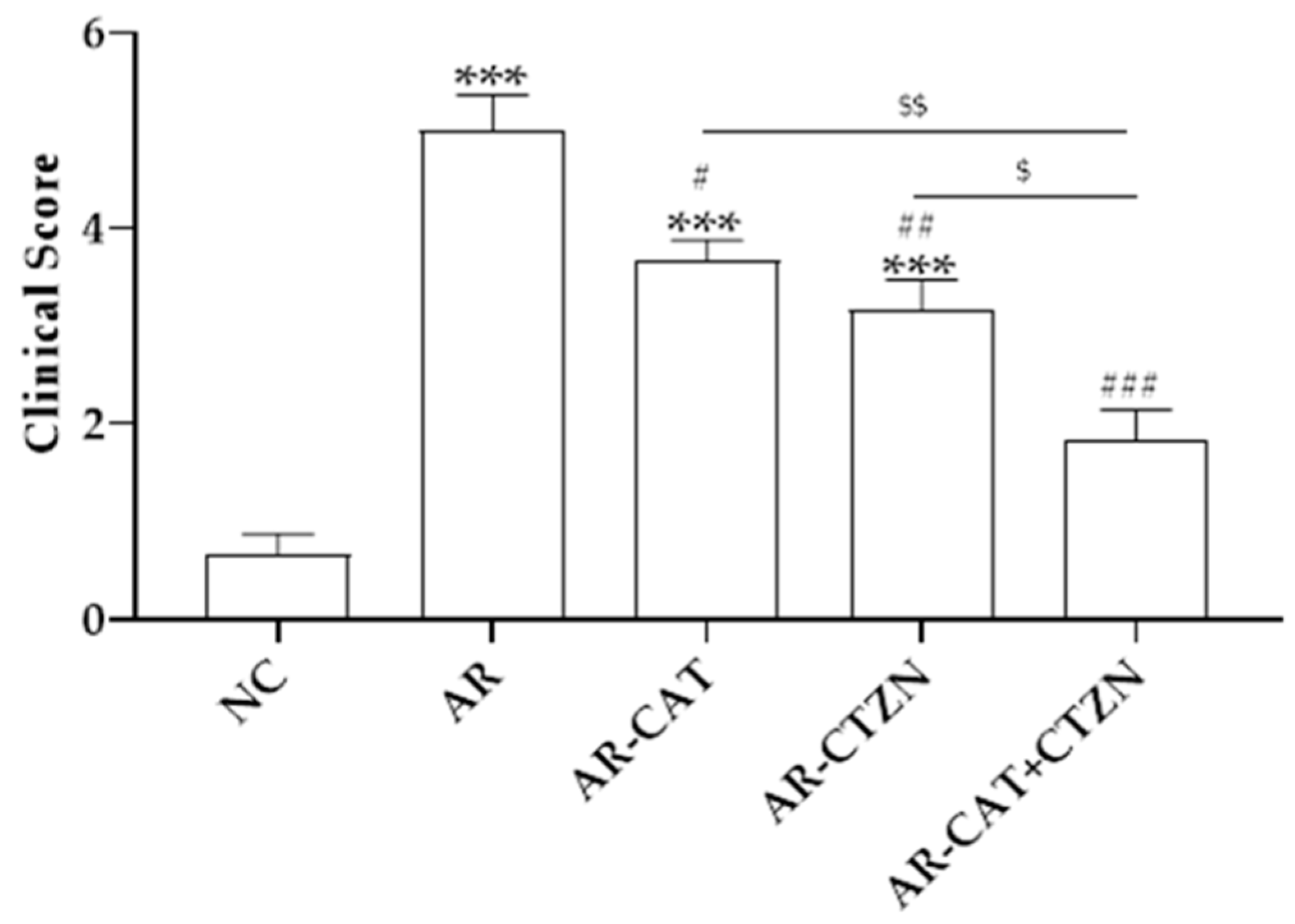
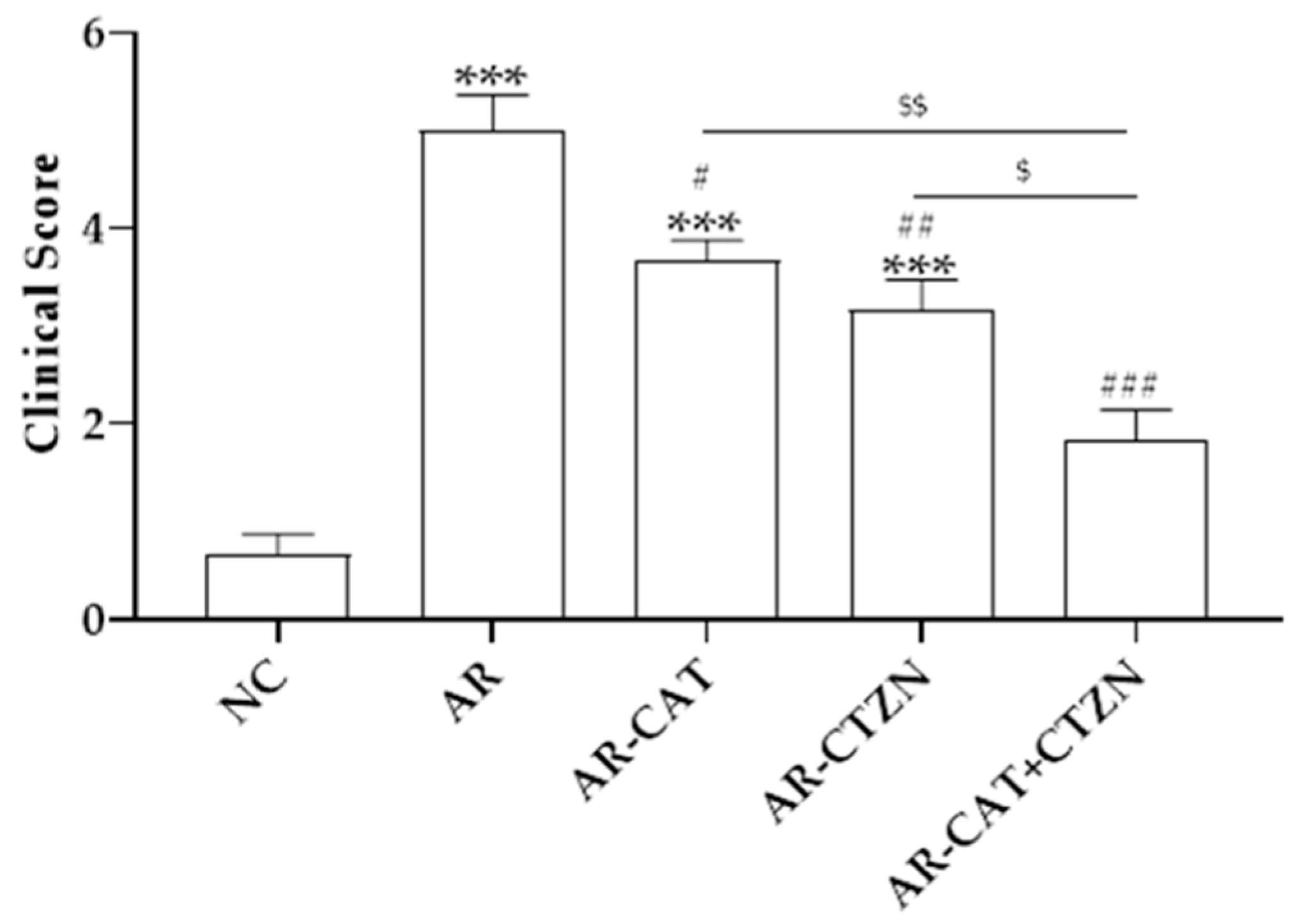
3.1. Effect of Different Treatment Regimens on AR Clinical Scores
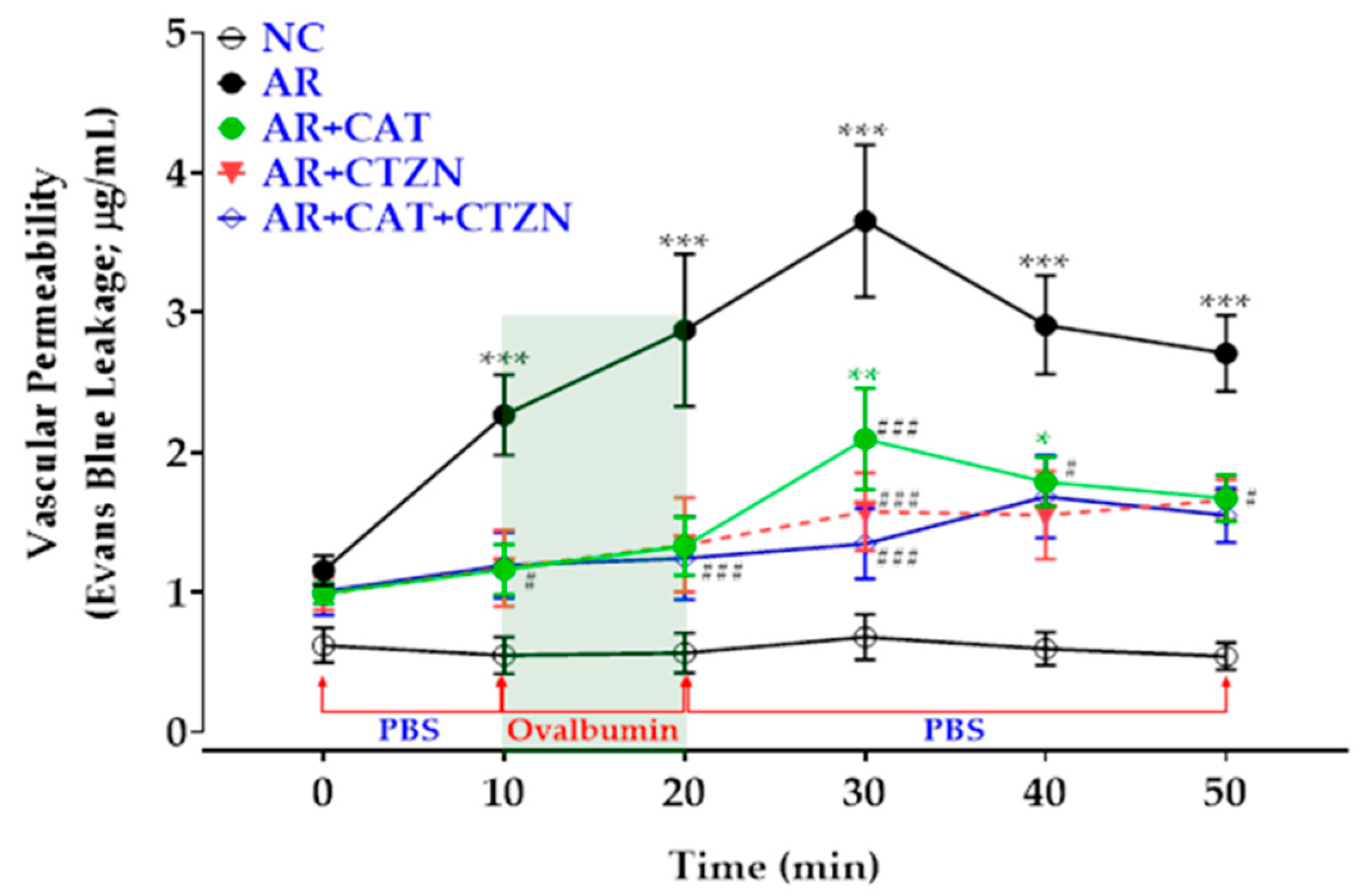
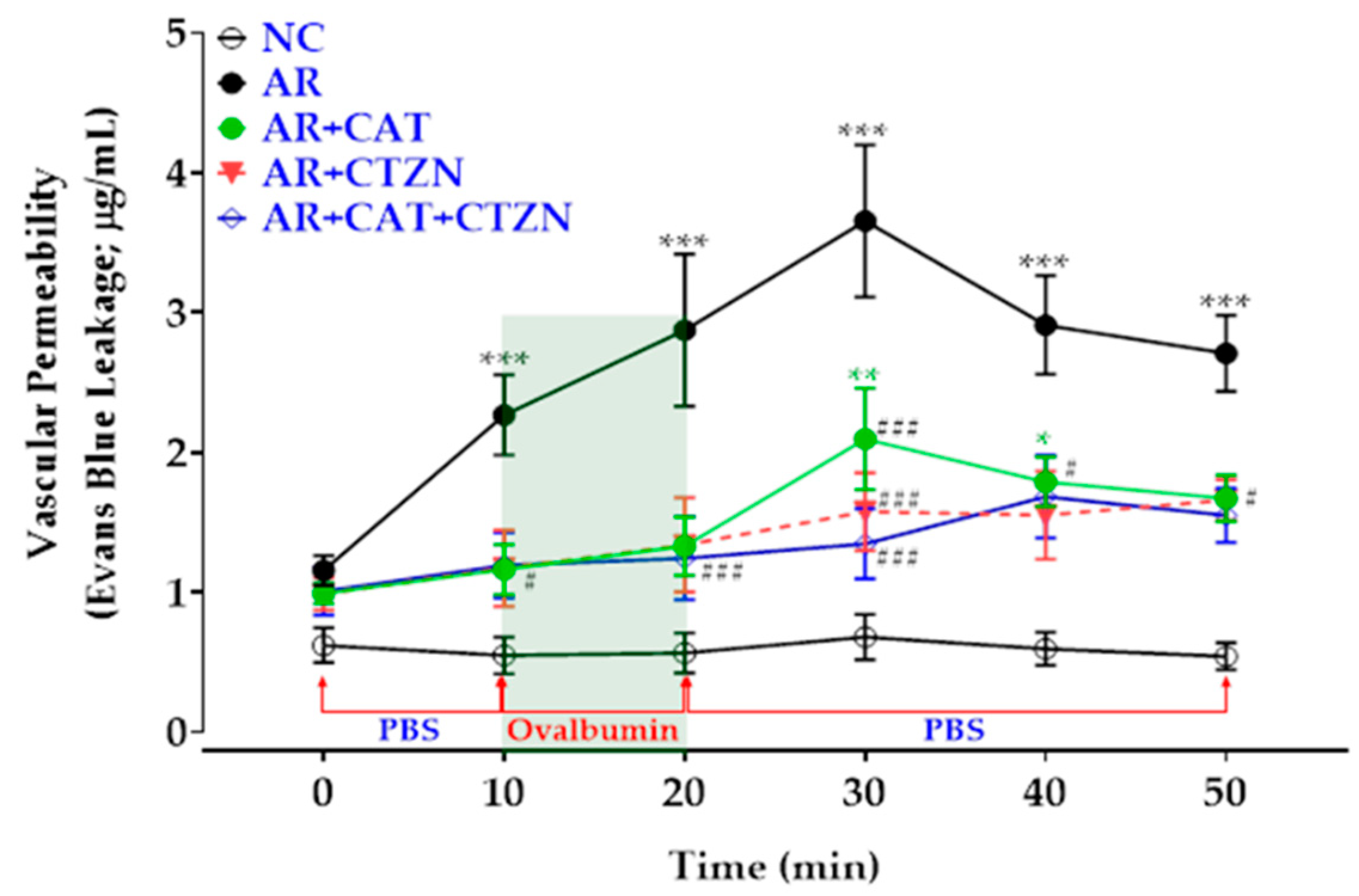
3.2. Effect of Different Treatment Regimens on Vascular Permeability
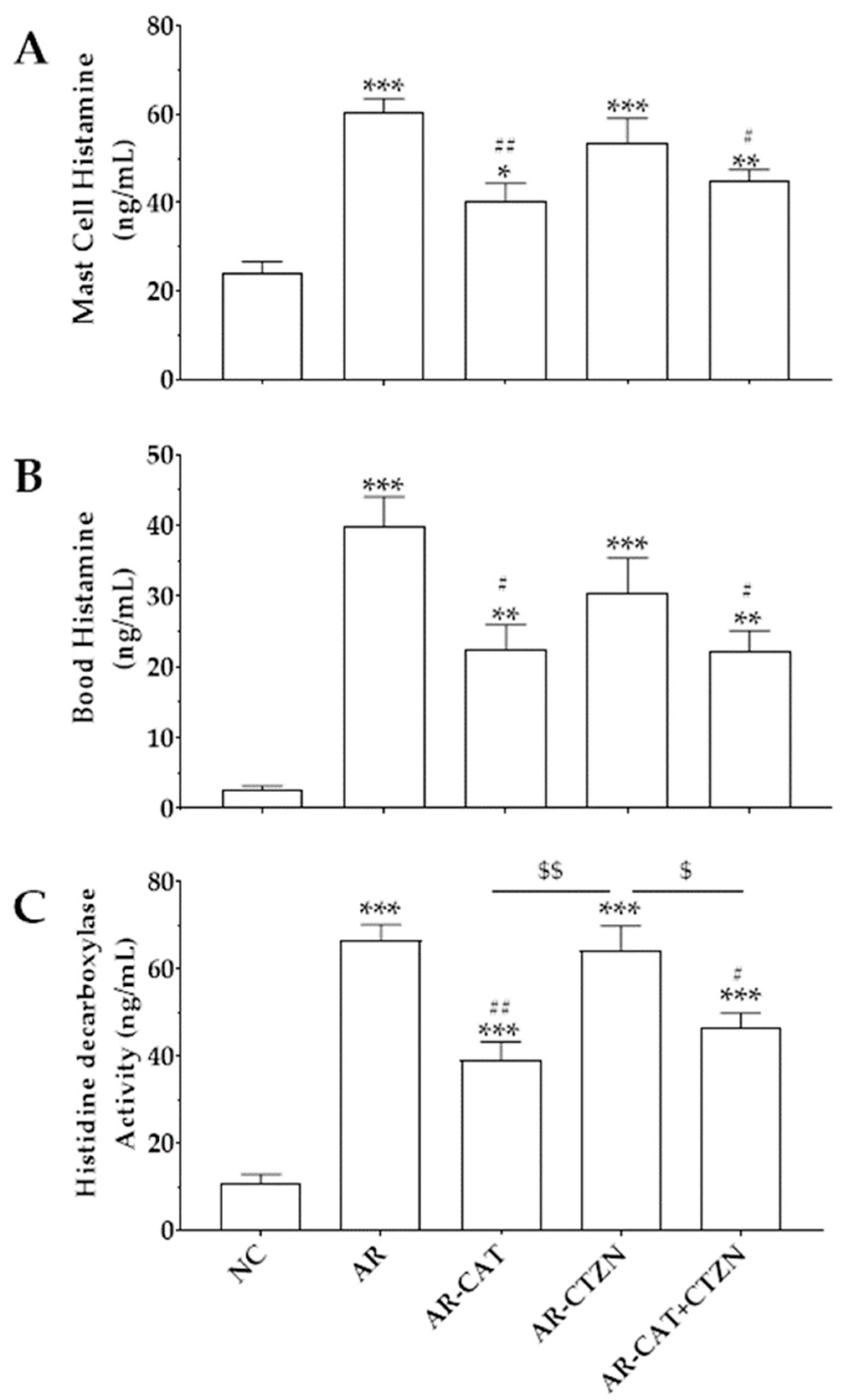
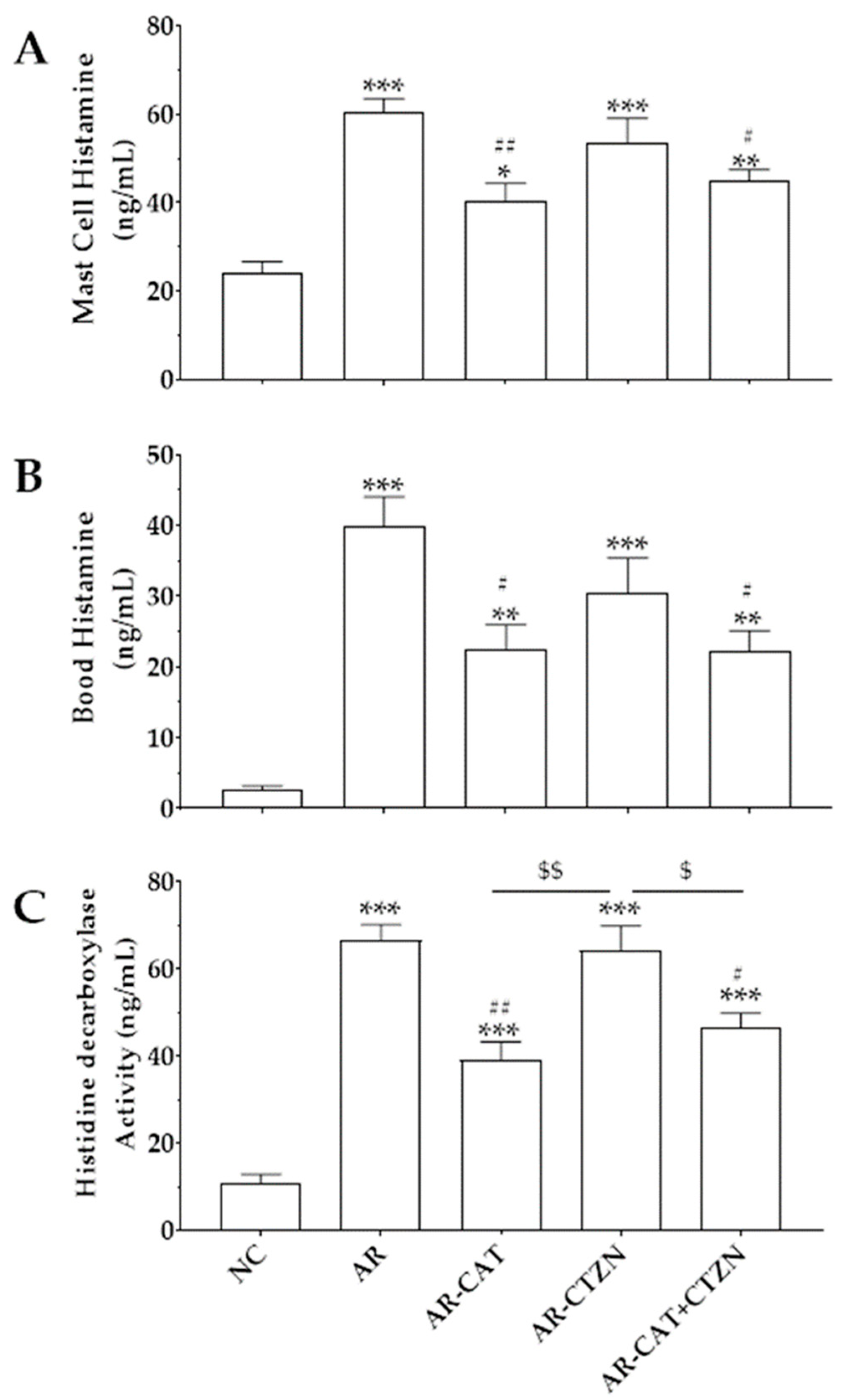
3.3. Effect of Different Treatment Regimens on Mast Cell and Blood Histamine
3.4. Effect of Different Treatment Regimens on HDC Activity
3.5. Effect of Different Treatment Regimens on Barbiturate-Induced Sleeping Time in Balb/c Mice
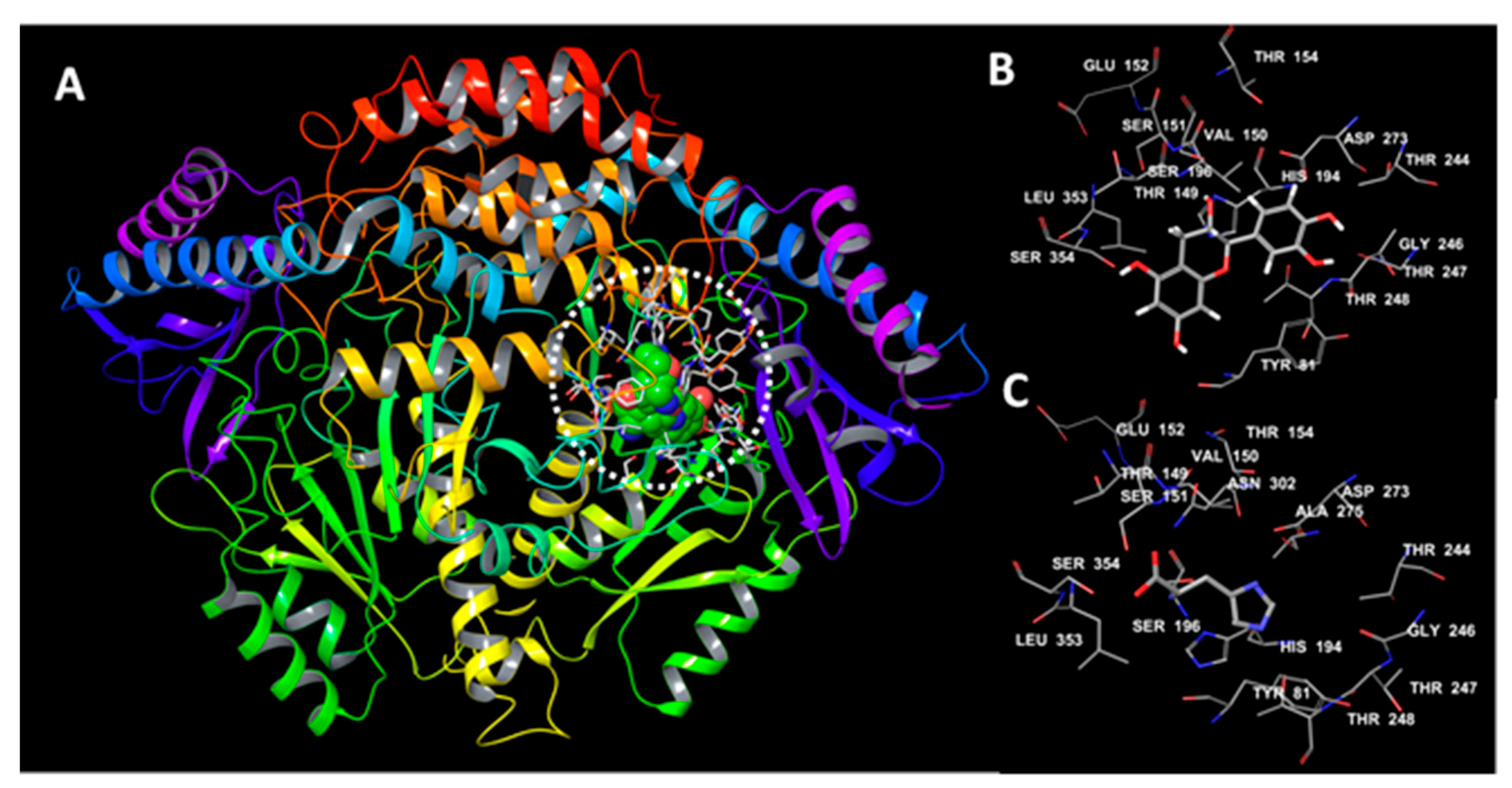
3.6. Molecular Docking Studies
3.7. Binding Energy Decomposition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Terada, T.; Kawata, R. Diagnosis and Treatment of Local Allergic Rhinitis. Pathogens 2022, 11, 80. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, Z.A.; Walker, A.; Pirwani, M.M.; Tahiri, M.; Syed, I. Allergic rhinitis: Diagnosis and management. Br. J. Hosp. Med. 2022, 83, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Seidman, M.D.; Gurgel, R.K.; Lin, S.Y.; Schwartz, S.R.; Baroody, F.M.; Bonner, J.R.; Dawson, D.E.; Dykewicz, M.S.; Hackell, J.M.; Han, J.K.; et al. Clinical practice guideline: Allergic rhinitis executive summary. Otolaryngol. Head Neck Surg. 2015, 152, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Bleier, B.; Wei, Y. Definition and characteristics of acute exacerbation in adult patients with chronic rhinosinusitis: A systematic review. J. Otolaryngol. Head Neck Surg. 2020, 49, 62. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, K.; Ogasawara, M. The Role of Histamine in the Pathophysiology of Asthma and the Clinical Efficacy of Antihistamines in Asthma Therapy. Int. J. Mol. Sci. 2019, 20, 1733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liva, G.A.; Karatzanis, A.D.; Prokopakis, E.P. Review of Rhinitis: Classification, Types, Pathophysiology. J. Clin. Med. 2021, 10, 3183. [Google Scholar] [CrossRef]
- Eifan, A.O.; Durham, S.R. Pathogenesis of rhinitis. Clin. Exp. Allergy J. Br. Soc. Allergy Cllin. Immunol. 2016, 46, 1139–1151. [Google Scholar] [CrossRef]
- Van Gerven, L.; Steelant, B.; Hellings, P.W. Nasal hyperreactivity in rhinitis: A diagnostic and therapeutic challenge. Allergy 2018, 73, 1784–1791. [Google Scholar] [CrossRef] [Green Version]
- Brosnan, M.E.; Brosnan, J.T. Histidine Metabolism and Function. J. Nutr. 2020, 150, 2570s–2575s. [Google Scholar] [CrossRef]
- Hirasawa, N. Expression of Histidine Decarboxylase and Its Roles in Inflammation. Int. J. Mol. Sci. 2019, 20, 376. [Google Scholar] [CrossRef] [Green Version]
- Komori, H.; Nitta, Y.; Ueno, H.; Higuchi, Y. Structural study reveals that Ser-354 determines substrate specificity on human histidine decarboxylase. J. Biol. Chem. 2012, 287, 29175–29183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakrania, A.K.; Patel, S.S. Combination treatment for allergic conjunctivitis—Plant derived histidine decarboxylase inhibitor and H1 antihistaminic drug. Exp. Eye Res. 2015, 137, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Hossenbaccus, L.; Linton, S.; Garvey, S.; Ellis, A.K. Towards definitive management of allergic rhinitis: Best use of new and established therapies. Allergy Asthma Clin. Immunol. 2020, 16, 39. [Google Scholar] [CrossRef] [PubMed]
- Singh Randhawa, A.; Mohd Noor, N.; Md Daud, M.K.; Abdullah, B. Efficacy and Safety of Bilastine in the Treatment of Allergic Rhinitis: A Systematic Review and Meta-analysis. Front. Pharmacol. 2021, 12, 731201. [Google Scholar] [CrossRef]
- Miligkos, M.; Dakoutrou, M.; Statha, E.; Theochari, N.A.; Mavroeidi, I.A.; Pankozidou, Ι.; Papaconstadopoulos, I.; Papadopoulos, N.G. Newer-generation antihistamines and the risk of adverse events in children: A systematic review. Pediatr. Allergy Immunol. 2021, 32, 1533–1558. [Google Scholar] [CrossRef]
- Ozdemir, P.G.; Karadag, A.S.; Selvi, Y.; Boysan, M.; Bilgili, S.G.; Aydin, A.; Onder, S. Assessment of the effects of antihistamine drugs on mood, sleep quality, sleepiness, and dream anxiety. Int. J. Psychiatry Clin. Pract. 2014, 18, 161–168. [Google Scholar] [CrossRef]
- Nitta, Y.; Kikuzaki, H.; Ueno, H. Food components inhibiting recombinant human histidine decarboxylase activity. J. Agric. Food Chem. 2007, 55, 299–304. [Google Scholar] [CrossRef]
- Rodríguez-Caso, C.; Rodríguez-Agudo, D.; Sánchez-Jiménez, F.; Medina, M.A. Green tea epigallocatechin-3-gallate is an inhibitor of mammalian histidine decarboxylase. Cell. Mol. Life Sci. 2003, 60, 1760–1763. [Google Scholar] [CrossRef]
- Ruiz-Pérez, M.V.; Pino-Ángeles, A.; Medina, M.A.; Sánchez-Jiménez, F.; Moya-García, A.A. Structural perspective on the direct inhibition mechanism of EGCG on mammalian histidine decarboxylase and DOPA decarboxylase. J. Chem. Inform. Model. 2012, 52, 113–119. [Google Scholar] [CrossRef]
- Kawai, M.; Hirano, T.; Higa, S.; Arimitsu, J.; Maruta, M.; Kuwahara, Y.; Ohkawara, T.; Hagihara, K.; Yamadori, T.; Shima, Y.; et al. Flavonoids and Related Compounds as Anti-Allergic Substances. Allergol. Int. 2007, 56, 113–123. [Google Scholar] [CrossRef] [Green Version]
- Kandeel, M.; Al-Nazawi, M. Virtual screening and repurposing of FDA approved drugs against COVID-19 main protease. Life Sci. 2020, 251, 117627. [Google Scholar] [CrossRef] [PubMed]
- Morsy, M.A.; El-Sheikh, A.A.; Ibrahim, A.R.; Venugopala, K.N.; Kandeel, M. In silico and in vitro identification of secoisolariciresinol as a re-sensitizer of P-glycoprotein-dependent doxorubicin-resistance NCI/ADR-RES cancer cells. Peer J. 2020, 8, e9163. [Google Scholar] [CrossRef] [PubMed]
- Ezz-Eldin, Y.M.; Aboseif, A.A.; Khalaf, M.M. Potential anti-inflammatory and immunomodulatory effects of carvacrol against ovalbumin-induced asthma in rats. Life Sci. 2020, 242, 117222. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, N.; Terawaki, S.; Shimizu, K.; Oikawa, D.; Sakamoto, H.; Sunami, K.; Tokunaga, F. Th2 cell-derived histamine is involved in nasal Th2 infiltration in mice. Inflammat. Res. 2021, 70, 539–541. [Google Scholar] [CrossRef]
- Yu, J.; Tuo, F.; Luo, Y.; Xu, J. Effect of gestational and lactational nonylphenol exposure on airway inflammation in ovalbumin-induced asthmatic rat pups. Chemosphere 2020, 250, 126244. [Google Scholar] [CrossRef]
- Elieh Ali Komi, D.; Wöhrl, S.; Bielory, L. Mast Cell Biology at Molecular Level: A Comprehensive Review. Clin. Rev. Allergy Immunol. 2020, 58, 342–365. [Google Scholar] [CrossRef]
- Shirasaki, H.; Kanaizumi, E.; Seki, N.; Himi, T. Localization and upregulation of the nasal histamine H1 receptor in perennial allergic rhinitis. Mediat. Inflammat. 2012, 2012, 951316. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Cheng, L.; Hong, J. The clinical use of cetirizine in the treatment of allergic rhinitis. Pharmacology 2013, 92, 14–25. [Google Scholar] [CrossRef]
- Komi, D.E.A.; Mortaz, E.; Amani, S.; Tiotiu, A.; Folkerts, G.; Adcock, I.M. The Role of Mast Cells in IgE-Independent Lung Diseases. Clin. Rev. Allergy Immunol. 2020, 58, 377–387. [Google Scholar] [CrossRef] [Green Version]
- Franova, S.; Joskova, M.; Sadlonova, V.; Pavelcikova, D.; Mesarosova, L.; Novakova, E.; Sutovska, M. Experimental model of allergic asthma. Adv. Exp. Med. Biol. 2013, 756, 49–55. [Google Scholar] [CrossRef]
- Spilsbury, A.; Vauzour, D.; Spencer, J.P.E.; Rattray, M. Regulation of NF-κB activity in astrocytes: Effects of flavonoids at dietary-relevant concentrations. Biochem. Biophysical. Res. Commun. 2012, 418, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Dalal, P.J.; Muller, W.A.; Sullivan, D.P. Endothelial Cell Calcium Signaling during Barrier Function and Inflammation. Am. J. Pathol. 2020, 190, 535–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muntean, I.A.; Bocsan, I.C.; Miron, N.; Buzoianu, A.D.; Deleanu, D. How Could We Influence Systemic Inflammation in Allergic Rhinitis? The Role of H1 Antihistamines. Oxid. Med. Cell. Longev. 2018, 2018, 3718437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaguzel, D.; Sarac, B.E.; Akel Bilgic, H.; Summak, G.Y.; Unal, M.A.; Kalayci, O.; Karaaslan, C. House dust mite-derived allergens effect on matrix metalloproteases in airway epithelial cells. Exp. Lung Res. 2021, 47, 436–450. [Google Scholar] [CrossRef]
- Narita, S.-I.; Goldblum, R.M.; Watson, C.S.; Brooks, E.G.; Estes, D.M.; Curran, E.M.; Terumi, M.-H. Environmental Estrogens Induce Mast Cell Degranulation and Enhance IgE-Mediated Release of Allergic Mediators. Environ. Health Persp. 2007, 115, 48–52. [Google Scholar] [CrossRef] [Green Version]
- Neumann, J.; Grobe, J.M.; Weisgut, J.; Schwelberger, H.G.; Fogel, W.A.; Marušáková, M.; Wache, H.; Bähre, H.; Buchwalow, I.B.; Dhein, S.; et al. Histamine can be Formed and Degraded in the Human and Mouse Heart. Front. Pharmacol. 2021, 12, 582916. [Google Scholar] [CrossRef] [PubMed]
- Oda, Y.; Nakata, K.; Miyano, H.; Mizukoshi, T.; Yamaguchi, H.; Kashiwagi, T. Structural insights into the enhanced thermostability of cysteine substitution mutants of L-histidine decarboxylase from Photobacterium phosphoreum. J. Biochem. 2022, 171, 31–40. [Google Scholar] [CrossRef]
- Olmo, M.T.; Sánchez-Jiménez, F.; Medina, M.A.; Hayashi, H. Spectroscopic Analysis of Recombinant Rat Histidine Decarboxylase. J. Biochem. 2002, 132, 433–439. [Google Scholar] [CrossRef] [Green Version]
- Grevers, G.; Kastenbauer, E. Functional Morphology of Nasal Blood Vessels in Humans. Acta Oto-Laryngol. 1996, 116, 312–315. [Google Scholar] [CrossRef]
- Korosec, P.; Turner, P.J.; Silar, M.; Kopac, P.; Kosnik, M.; Gibbs, B.F.; Shamji, M.H.; Custovic, A.; Rijavec, M. Basophils, high-affinity IgE receptors, and CCL2 in human anaphylaxis. J. Allergy Clin. Immunol. 2017, 140, 750–758.e715. [Google Scholar] [CrossRef] [Green Version]
- Mizuguchi, H.; Das, A.K.; Maeyama, K.; Dev, S.; Shahriar, M.; Kitamura, Y.; Takeda, N.; Fukui, H. Antihistamines suppress upregulation of histidine decarboxylase gene expression with potencies different from their binding affinities for histamine H1 receptor in toluene 2,4-diisocyanate-sensitized rats. J. Pharmacol. Sci. 2016, 130, 212–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nitta, Y.; Kikuzaki, H.; Azuma, T.; Ye, Y.; Sakaue, M.; Higuchi, Y.; Komori, H.; Ueno, H. Inhibitory activity of Filipendula ulmaria constituents on recombinant human histidine decarboxylase. Food Chem. 2013, 138, 1551–1556. [Google Scholar] [CrossRef] [PubMed]
- Ain, Q.; Khan, H. Pharmacological basis for sedative and hypnotic like effects of Pyrus pashia using in vivo experimental models. Int. J. Geriatr. Psychiatry 2019, 34, 1345–1350. [Google Scholar] [CrossRef]
- Ishola, I.O.; Katola, F.O.; Adeyemi, O.O. Involvement of GABAergic and nitrergic systems in the anxiolytic and hypnotic effects of Curcuma longa: Its interaction with anxiolytic-hypnotics. Drug Metabol. Pers. Ther. 2020. [Google Scholar] [CrossRef] [PubMed]
- Polidori, M. Antioxidant micronutrients in the prevention of age-related diseases. J. Postgrad. Med. 2003, 49, 229. [Google Scholar] [PubMed]
- Gehanno, P.; Bremard-Oury, C.; Zeisser, P. Comparison of ebastine to cetirizine in seasonal allergic rhinitis in adults. Ann. Allergy Asthma Immunol. 1996, 76, 507–512. [Google Scholar] [CrossRef]
- Stromsnes, K.; Mas-Bargues, C.; Gambini, J.; Gimeno-Mallench, L. Protective Effects of Polyphenols Present in Mediterranean Diet on Endothelial Dysfunction. Oxid. Med. Cell. Longev. 2020, 2020, 2097096. [Google Scholar] [CrossRef]
- Heenan, K.A.; Carrillo, A.E.; Fulton, J.L.; Ryan, E.J.; Edsall, J.R.; Rigopoulos, D.; Markofski, M.M.; Flouris, A.D.; Dinas, P.C. Effects of Nutrition/Diet on Brown Adipose Tissue in Humans: A Systematic Review and Meta-Analysis. Nutrients 2020, 12, 2752. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feature | Characteristic | Clinical Scoring |
|---|---|---|
| Nasal itch | None | 0 |
| Scratching nose lightly one to two times | 1 | |
| Scratching the nose and face constantly | 2 | |
| Sneeze | None | 0 |
| One to three times | 1 | |
| Four to ten times | 2 | |
| Eleven or more times | 3 | |
| Nasal discharge | None | 0 |
| Secretions flow to anterior nostril | 1 | |
| Secretions surpass anterior nostril | 2 | |
| Secretions cover the face | 3 |
| Parameter | Vehicle | CAT | CTZN | CAT+CTZN |
|---|---|---|---|---|
| Barbiturate-induced sleeping time (min) | 31.33 ± 1.66 | 29.66 ± 0.76 | 34.66 ± 1.02 * | 30.66 ± 0.4 |
| Compound | XP Docking Score | H-Bond | Elec | S354 H-Bond | S354 Distance |
|---|---|---|---|---|---|
| Catechin | −8.49 | −3.25 | −39.09 | −0.47 | 2.9 |
| L-histidine | −6.33 | −3.6 | −25.57 | −1 | 2.7 |
| Histamine | −5.46 | −1.44 | −20.88 | −0.1 | 2.2 |
| Cetirizine | −9.4 | 0 | −8.9 | 0 | 2.7 |
| Histidine methyl ester | −5.29 | −1 | −16.01 | −1 | 1.7 |
| Compound | Binding Energy | ∆Eelec | ∆Evdw | ∆EH | ∆ELipo | ∆Esolv |
|---|---|---|---|---|---|---|
| Catechin | −50.86 | −36.83 | −28.22 | −1.60 | −28.10 | 39.09 |
| L-histidine | −37.64 | −35.87 | −18.28 | −2.55 | −10.19 | 25.57 |
| Histamine | −32.27 | −24.75 | −16.00 | −0.81 | −11.74 | 20.88 |
| Cetirizine | −49.36 | −13.19 | −15.25 | −1 | −59.5 | 25.53 |
| Histidine methyl ester | −51.1 | −46.13 | −23.67 | −2.56 | −12.82 | 31.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morsy, M.A.; Patel, S.S.; Bakrania, A.; Kandeel, M.; Nair, A.B.; Shah, J.N.; Akrawi, S.H.; El-Daly, M. Ameliorative Effect of a Neoteric Regimen of Catechin plus Cetirizine on Ovalbumin-Induced Allergic Rhinitis in Rats. Life 2022, 12, 820. https://doi.org/10.3390/life12060820
Morsy MA, Patel SS, Bakrania A, Kandeel M, Nair AB, Shah JN, Akrawi SH, El-Daly M. Ameliorative Effect of a Neoteric Regimen of Catechin plus Cetirizine on Ovalbumin-Induced Allergic Rhinitis in Rats. Life. 2022; 12(6):820. https://doi.org/10.3390/life12060820
Chicago/Turabian StyleMorsy, Mohamed A., Snehal S. Patel, Anita Bakrania, Mahmoud Kandeel, Anroop B. Nair, Jigar N. Shah, Sabah H. Akrawi, and Mahmoud El-Daly. 2022. "Ameliorative Effect of a Neoteric Regimen of Catechin plus Cetirizine on Ovalbumin-Induced Allergic Rhinitis in Rats" Life 12, no. 6: 820. https://doi.org/10.3390/life12060820
APA StyleMorsy, M. A., Patel, S. S., Bakrania, A., Kandeel, M., Nair, A. B., Shah, J. N., Akrawi, S. H., & El-Daly, M. (2022). Ameliorative Effect of a Neoteric Regimen of Catechin plus Cetirizine on Ovalbumin-Induced Allergic Rhinitis in Rats. Life, 12(6), 820. https://doi.org/10.3390/life12060820