
Endocrinopathies and Male Infertility



Abstract
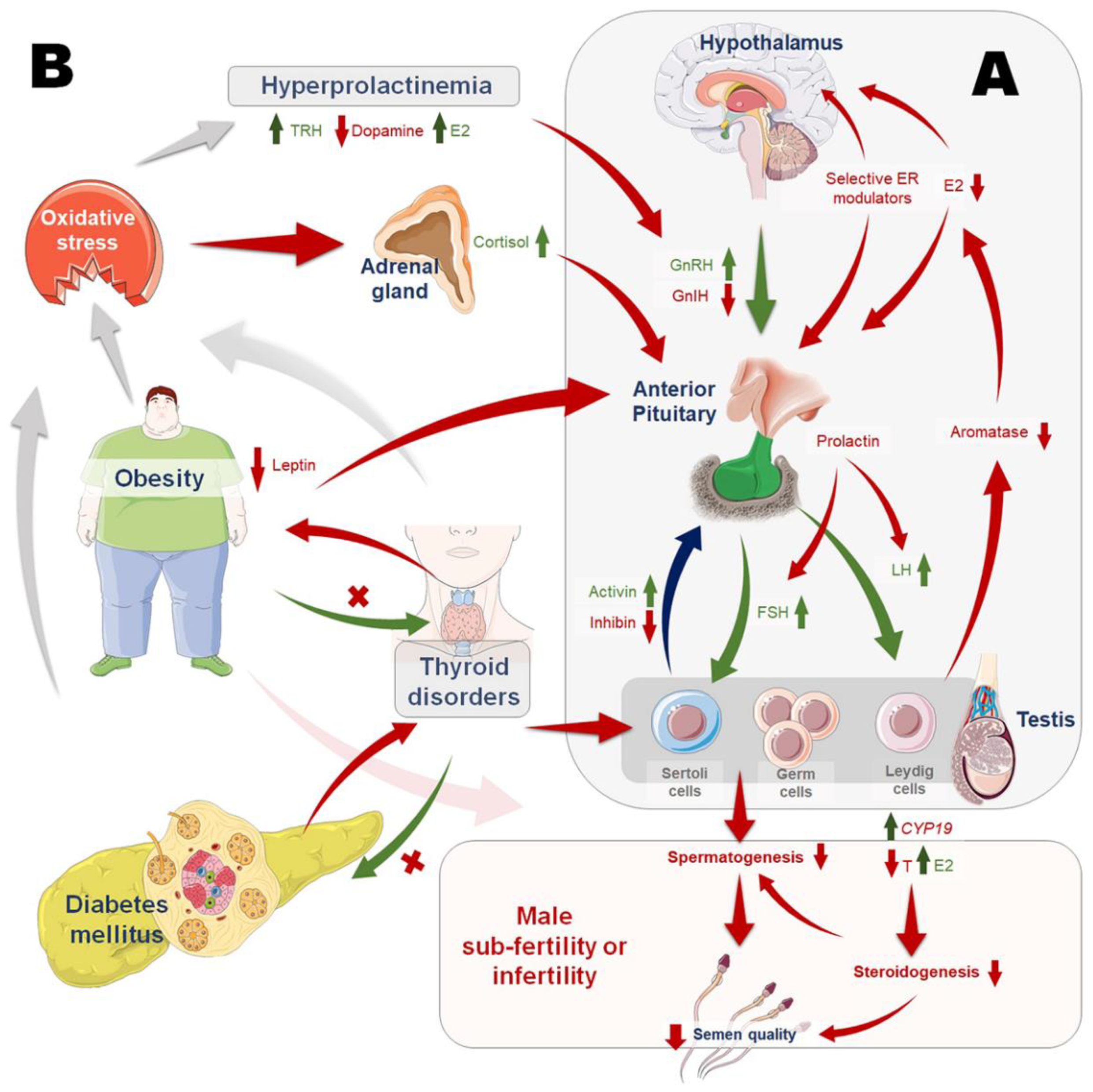
1. Introduction
2. Hypogonadotropic Hypogonadism
3. Hypergonadotropic Hypogonadism
4. Androgen Excess
5. Estrogen Excess
6. Hypothyroidism
7. Hyperthyroidism
8. Hyperprolactinemia
9. Insulin Disorders and Diabetes Mellitus
10. Obesity and Endocrine Disruption
11. Endocrinopathies, Oxidative Status of Male Reproduction
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sengupta, P.; Dutta, S.; Krajewska-Kulak, E. The disappearing sperms: Analysis of reports published between 1980 and 2015. Am. J. Mens Health 2017, 11, 1279–1304. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, P.; Borges, E., Jr.; Dutta, S.; Krajewska-Kulak, E. Decline in sperm count in european men during the past 50 years. Hum. Exp. Toxicol. 2018, 37, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, P.; Nwagha, U.; Dutta, S.; Krajewska-Kulak, E.; Izuka, E. Evidence for decreasing sperm count in african population from 1965 to 2015. Afr. Health Sci. 2017, 17, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, P.; Dutta, S.; Tusimin, M.B.; Irez, T.; Krajewska-Kulak, E. Sperm counts in asian men: Reviewing the trend of past 50 years. Asian Pac. J. Reprod. 2018, 7, 87–92. [Google Scholar] [CrossRef]
- Sengupta, P. Reviewing reports of semen volume and male aging of last 33 years: From 1980 through 2013. Asian Pac. J. Reprod. 2015, 4, 242–246. [Google Scholar] [CrossRef]
- Muhamad, S.; Sengupta, P.; Ramli, R.; Nasir, A. Sociodemographic factors associated with semen quality among malaysian men attending fertility clinic. Andrologia 2019, 51, e13383. [Google Scholar] [CrossRef]
- Barratt, C.L.; Björndahl, L.; De Jonge, C.J.; Lamb, D.J.; Osorio Martini, F.; McLachlan, R.; Oates, R.D.; van der Poel, S.; St John, B.; Sigman, M. The diagnosis of male infertility: An analysis of the evidence to support the development of global who guidance—Challenges and future research opportunities. Hum. Reprod. Update 2017, 23, 660–680. [Google Scholar] [CrossRef]
- Akhigbe, R.; Dutta, S.; Sengupta, P.; Chhikara, B.S. Adropin in immune and energy balance: ‘A molecule of interest’in male reproduction. Chem. Biol. Lett. 2021, 8, 213–223. [Google Scholar]
- Dutta, S.; Sengupta, P.; Muhamad, S. Male reproductive hormones and semen quality. Asian Pac. J. Reprod. 2019, 8, 189–194. [Google Scholar] [CrossRef]
- Sengupta, P.; Arafa, M.; Elbardisi, H. Hormonal regulation of spermatogenesis. In Molecular Signaling in Spermatogenesis and Male Infertility; CRC Press: Boca Raton, FL, USA, 2019; pp. 41–49. [Google Scholar]
- Lipshultz, L.I.; Howards, S.S.; Niederberger, C.S. Infertility in the Male; Cambridge University Press: Cambridge, UK, 2009. [Google Scholar]
- Kim, H.H.; Schlegel, P.N. Endocrine manipulation in male infertility. Urol. Clin. N. Am. 2008, 35, 303–318. [Google Scholar] [CrossRef]
- Haywood, S.; Lam, I.; Laborde, E.L.; Brannigan, R. Endocrinopathies. In Male Infertility; Springer: Berlin/Heidelberg, Germany, 2020; pp. 49–56. [Google Scholar]
- Fraietta, R.; Zylberstejn, D.S.; Esteves, S.C. Hypogonadotropic hypogonadism revisited. Clinics 2013, 68 (Suppl. 1), 81–88. [Google Scholar] [CrossRef]
- Coviello, A.D.; Matsumoto, A.M.; Bremner, W.J.; Herbst, K.L.; Amory, J.K.; Anawalt, B.D.; Sutton, P.R.; Wright, W.W.; Brown, T.R.; Yan, X.; et al. Low-dose human chorionic gonadotropin maintains intratesticular testosterone in normal men with testosterone-induced gonadotropin suppression. J. Clin. Endocrinol. Metab. 2005, 90, 2595–2602. [Google Scholar] [CrossRef]
- Dohle, G.R.; Smit, M.; Weber, R.F. Androgens and male fertility. World J. Urol. 2003, 21, 341–345. [Google Scholar] [CrossRef]
- Menon, D.K. Successful treatment of anabolic steroid-induced azoospermia with human chorionic gonadotropin and human menopausal gonadotropin. Fertil. Steril. 2003, 79 (Suppl. 3), 1659–1661. [Google Scholar] [CrossRef]
- Ioannidou-Kadis, S.; Wright, P.J.; Neely, R.D.; Quinton, R. Complete reversal of adult-onset isolated hypogonadotropic hypogonadism with clomiphene citrate. Fertil. Steril. 2006, 86, e1515–e1519. [Google Scholar]
- Amaral, S.; Moreno, A.J.; Santos, M.S.; Seiça, R.; Ramalho-Santos, J. Effects of hyperglycemia on sperm and testicular cells of goto-kakizaki and streptozotocin-treated rat models for diabetes. Theriogenology 2006, 66, 2056–2067. [Google Scholar] [CrossRef] [PubMed]
- Rama Raju, G.A.; Jaya Prakash, G.; Murali Krishna, K.; Madan, K.; Siva Narayana, T.; Ravi Krishna, C.H. Noninsulin-dependent diabetes mellitus: Effects on sperm morphological and functional characteristics, nuclear DNA integrity and outcome of assisted reproductive technique. Andrologia 2012, 44 (Suppl. 1), 490–498. [Google Scholar] [CrossRef] [PubMed]
- Soudamani, S.; Malini, T.; Balasubramanian, K. Effects of streptozotocin-diabetes and insulin replacement on the epididymis of prepubertal rats: Histological and histomorphometric studies. Endocr. Res. 2005, 31, 81–98. [Google Scholar] [CrossRef] [PubMed]
- Ballester, J.; Muñoz, M.C.; Domínguez, J.; Rigau, T.; Guinovart, J.J.; Rodríguez-Gil, J.E. Insulin-dependent diabetes affects testicular function by fsh- and lh-linked mechanisms. J. Androl. 2004, 25, 706–719. [Google Scholar] [CrossRef]
- Dabaja, A.A.; Schlegel, P.N. Microdissection testicular sperm extraction: An update. Asian J. Androl. 2013, 15, 35–39. [Google Scholar] [CrossRef]
- Pavlovich, C.P.; King, P.; Goldstein, M.; Schlegel, P.N. Evidence of a treatable endocrinopathy in infertile men. J. Urol. 2001, 165, 837–841. [Google Scholar] [CrossRef]
- Dabbous, Z.; Atkin, S.L. Hyperprolactinaemia in male infertility: Clinical case scenarios. Arab J. Urol. 2018, 16, 44–52. [Google Scholar] [CrossRef]
- Singh, P.; Singh, M.; Cugati, G.; Singh, A. Hyperprolactinemia: An often missed cause of male infertility. J. Hum. Reprod. Sci. 2011, 4, 102. [Google Scholar] [CrossRef] [PubMed]
- Sohrabvand, F.; Jafari, M.; Shariat, M.; Haghollahi, F.; Lotfi, M. Frequency and epidemiologic aspects of male infertility. Acta Med. Iran. 2015, 53, 231–235. [Google Scholar]
- Tsutsumi, R.; Webster, N.J. Gnrh pulsatility, the pituitary response and reproductive dysfunction. Endocr. J. 2009, 56, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Stamou, M.I.; Georgopoulos, N.A. Kallmann syndrome: Phenotype and genotype of hypogonadotropic hypogonadism. Metabolism 2018, 86, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Sussman, E.M.; Chudnovsky, A.; Niederberger, C.S. Hormonal evaluation of the infertile male: Has it evolved? Urol. Clin. N. Am. 2008, 35, 147–155. [Google Scholar] [CrossRef]
- Liu, P.Y.; Baker, H.W.; Jayadev, V.; Zacharin, M.; Conway, A.J.; Handelsman, D.J. Induction of spermatogenesis and fertility during gonadotropin treatment of gonadotropin-deficient infertile men: Predictors of fertility outcome. J. Clin. Endocrinol. Metab. 2009, 94, 801–808. [Google Scholar] [CrossRef]
- Fernandez, C.J.; Chacko, E.C.; Pappachan, J.M. Male obesity-related secondary hypogonadism–pathophysiology, clinical implications and management. Eur. Endocrinol. 2019, 15, 83. [Google Scholar] [CrossRef]
- Santos-Filho, S.D. Menopause practice essentials: A short review. Int. J. Med. Sci. Technol. 2019, 9, 11–24. [Google Scholar]
- Schiff, J.D.; Ramírez, M.L.; Bar-Chama, N. Medical and surgical management male infertility. Endocrinol. Metab. Clin. N. Am. 2007, 36, 313–331. [Google Scholar] [CrossRef]
- Burris, A.S.; Clark, R.V.; Vantman, D.J.; Sherins, R.J. A low sperm concentration does not preclude fertility in men with isolated hypogonadotropic hypogonadism after gonadotropin therapy. Fertil. Steril. 1988, 50, 343–347. [Google Scholar] [CrossRef]
- Miyagawa, Y.; Tsujimura, A.; Matsumiya, K.; Takao, T.; Tohda, A.; Koga, M.; Takeyama, M.; Fujioka, H.; Takada, S.; Koide, T.; et al. Outcome of gonadotropin therapy for male hypogonadotropic hypogonadism at university affiliated male infertility centers: A 30-year retrospective study. J. Urol. 2005, 173, 2072–2075. [Google Scholar] [CrossRef]
- Farhat, R.; Al-zidjali, F.; Alzahrani, A.S. Outcome of gonadotropin therapy for male infertility due to hypogonadotrophic hypogonadism. Pituitary 2010, 13, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Whitten, S.J.; Nangia, A.K.; Kolettis, P.N. Select patients with hypogonadotropic hypogonadism may respond to treatment with clomiphene citrate. Fertil. Steril. 2006, 86, 1664–1668. [Google Scholar] [CrossRef]
- Burge, M.R.; Lanzi, R.A.; Skarda, S.T.; Eaton, R.P. Idiopathic hypogonadotropic hypogonadism in a male runner is reversed by clomiphene citrate. Fertil. Steril. 1997, 67, 783–785. [Google Scholar] [CrossRef]
- Ramasamy, R.; Scovell, J.M.; Kovac, J.R.; Lipshultz, L.I. Testosterone supplementation versus clomiphene citrate for hypogonadism: An age matched comparison of satisfaction and efficacy. J. Urol. 2014, 192, 875–879. [Google Scholar] [CrossRef] [PubMed]
- Layton, J.B.; Li, D.; Meier, C.R.; Sharpless, J.L.; Stürmer, T.; Jick, S.S.; Brookhart, M.A. Testosterone lab testing and initiation in the united kingdom and the united states, 2000 to 2011. J. Clin. Endocrinol. Metab. 2014, 99, 835–842. [Google Scholar] [CrossRef]
- Zirkin, B.R.; Santulli, R.; Awoniyi, C.A.; Ewing, L.L. Maintenance of advanced spermatogenic cells in the adult rat testis: Quantitative relationship to testosterone concentration within the testis. Endocrinology 1989, 124, 3043–3049. [Google Scholar] [CrossRef] [PubMed]
- Kalkanli, A.; Akdere, H.; Cevik, G.; Salabas, E.; Cilesiz, N.C.; Kadioglu, A. Hypergonadotropic hypogonadism: Management of infertility. Curr. Pharm. Design 2021, 27, 2790–2795. [Google Scholar] [CrossRef]
- Raman, J.D.; Schlegel, P.N. Aromatase inhibitors for male infertility. J. Urol. 2002, 167, 624–629. [Google Scholar] [CrossRef]
- Ramasamy, R.; Ricci, J.A.; Palermo, G.D.; Gosden, L.V.; Rosenwaks, Z.; Schlegel, P.N. Successful fertility treatment for klinefelter’s syndrome. J. Urol. 2009, 182, 1108–1113. [Google Scholar] [CrossRef] [PubMed]
- Samango-Sprouse, C.A.; Sadeghin, T.; Mitchell, F.L.; Dixon, T.; Stapleton, E.; Kingery, M.; Gropman, A.L. Positive effects of short course androgen therapy on the neurodevelopmental outcome in boys with 47,xxy syndrome at 36 and 72 months of age. Am. J. Med. Genet. 2013, 161, 501–508. [Google Scholar] [CrossRef]
- Davis, S.M.; Rogol, A.D.; Ross, J.L. Testis development and fertility potential in boys with klinefelter syndrome. Endocrinol. Metab. Clin. N. Am. 2015, 44, 843–865. [Google Scholar] [CrossRef]
- Lo, J.O.; Cori, D.F.; Norton, M.E.; Caughey, A.B. Noninvasive prenatal testing. Obs. Gynecol. Surv. 2014, 69, 89–99. [Google Scholar] [CrossRef]
- Turek, P.J.; Williams, R.H.; Gilbaugh, J.H., 3rd; Lipshultz, L.I. The reversibility of anabolic steroid-induced azoospermia. J. Urol. 1995, 153, 1628–1630. [Google Scholar] [CrossRef]
- Tan, R.S.; Vasudevan, D. Use of clomiphene citrate to reverse premature andropause secondary to steroid abuse. Fertil. Steril. 2003, 79, 203–205. [Google Scholar] [CrossRef]
- Coward, R.M.; Rajanahally, S.; Kovac, J.R.; Smith, R.P.; Pastuszak, A.W.; Lipshultz, L.I. Anabolic steroid induced hypogonadism in young men. J. Urol. 2013, 190, 2200–2205. [Google Scholar] [CrossRef] [PubMed]
- Kanayama, G.; Brower, K.J.; Wood, R.I.; Hudson, J.I.; Pope, H.G., Jr. Anabolic-androgenic steroid dependence: An emerging disorder. Addiction 2009, 104, 1966–1978. [Google Scholar] [CrossRef]
- Falhammar, H.; Nyström, H.F.; Ekström, U.; Granberg, S.; Wedell, A.; Thorén, M. Fertility, sexuality and testicular adrenal rest tumors in adult males with congenital adrenal hyperplasia. Eur. J. Endocrinol. 2012, 166, 441–449. [Google Scholar] [CrossRef]
- Jääskeläinen, J.; Kiekara, O.; Hippeläinen, M.; Voutilainen, R. Pituitary gonadal axis and child rate in males with classical 21-hydroxylase deficiency. J. Endocrinol. Investig. 2000, 23, 23–27. [Google Scholar] [CrossRef]
- Arlt, W.; Willis, D.S.; Wild, S.H.; Krone, N.; Doherty, E.J.; Hahner, S.; Han, T.S.; Carroll, P.V.; Conway, G.S.; Rees, D.A.; et al. Health status of adults with congenital adrenal hyperplasia: A cohort study of 203 patients. J. Clin. Endocrinol. Metab. 2010, 95, 5110–5121. [Google Scholar] [CrossRef] [PubMed]
- Murphy, H.; George, C.; de Kretser, D.; Judd, S. Successful treatment with icsi of infertility caused by azoospermia associated with adrenal rests in the testes: Case report. Hum. Reprod. 2001, 16, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.M.; Fefferman, R.A.; Shapiro, C.E. Reversible infertility in a man with 21-hydroxylase deficiency congenital adrenal hyperplasia. Fertil. Steril. 2005, 83, 223–225. [Google Scholar] [CrossRef]
- Roth, M.Y.; Amory, J.K.; Page, S.T. Treatment of male infertility secondary to morbid obesity. Nat. Clin. Prac. Endocrinol. Metab. 2008, 4, 415–419. [Google Scholar] [CrossRef]
- Alahmar, A.; Dutta, S.; Sengupta, P. Thyroid hormones in male reproduction and infertility. Asian Pac. J. Reprod. 2019, 8, 203–210. [Google Scholar] [CrossRef]
- Krajewska-Kulak, E.; Sengupta, P. Thyroid function in male infertility. Front. Endocrinol. 2013, 4, 174. [Google Scholar] [CrossRef]
- Sengupta, P.; Dutta, S. Thyroid disorders and semen quality. Biomed. Pharmacol. J. 2018, 11, 1–10. [Google Scholar] [CrossRef]
- Kumar, J.; Khurana, M.; Ammini, A.; Karmarkar, M.; Ahuja, M. Reproductive endocrine functions in men with primary hypothyroidism: Effect of thyroxine replacement. Horm. Res. Paediat. 1990, 34, 215–218. [Google Scholar] [CrossRef]
- Romano, R.M.; Gomes, S.N.; Cardoso, N.C.; Schiessl, L.; Romano, M.A.; Oliveira, C.A. New insights for male infertility revealed by alterations in spermatic function and differential testicular expression of thyroid-related genes. Endocrine 2017, 55, 607–617. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, D.K.; Roy, A.; Bhanja, S.; Chainy, G.B. Hypothyroidism impairs antioxidant defence system and testicular physiology during development and maturation. Gen. Comp. Endocrinol. 2008, 156, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Wortsman, J.; Rosner, W.; Dufau, M.L. Abnormal testicular function in men with primary hypothyroidism. Am. J. Med. 1987, 82, 207–212. [Google Scholar] [CrossRef]
- Carani, C.; Isidori, A.M.; Granata, A.; Carosa, E.; Maggi, M.; Lenzi, A.; Jannini, E.A. Multicenter study on the prevalence of sexual symptoms in male hypo- and hyperthyroid patients. J. Clin. Endocrinol. Metab. 2005, 90, 6472–6479. [Google Scholar] [CrossRef] [PubMed]
- Corrales Hernández, J.J.; Miralles García, J.M.; García Diez, L.C. Primary hypothyroidism and human spermatogenesis. Arch. Androl. 1990, 25, 21–27. [Google Scholar] [CrossRef]
- Donnelly, P.; White, C. Testicular dysfunction in men with primary hypothyroidism; reversal of hypogonadotrophic hypogonadism with replacement thyroxine. Clin. Endocrinol. 2000, 52, 197–201. [Google Scholar] [CrossRef]
- Krassas, G.E.; Perros, P. Thyroid disease and male reproductive function. J. Endocrinol. Investig. 2003, 26, 372–380. [Google Scholar] [CrossRef]
- Krassas, G.E.; Pontikides, N. Male reproductive function in relation with thyroid alterations. Best Pract. Res. Clin. Endocrinol. Metab. 2004, 18, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, S.; Chainy, G.B.; Mishro, M.M. Experimentally induced hypo- and hyper-thyroidism influence on the antioxidant defence system in adult rat testis. Andrologia 2003, 35, 131–140. [Google Scholar] [CrossRef]
- Griboff, S.I. Semen analysis in myxedema. Fertil. Steril. 1962, 13, 436–443. [Google Scholar] [CrossRef]
- Meeker, J.D.; Godfrey-Bailey, L.; Hauser, R. Relationships between serum hormone levels and semen quality among men from an infertility clinic. J. Androl. 2007, 28, 397–406. [Google Scholar] [CrossRef]
- Krassas, G.E.; Papadopoulou, F.; Tziomalos, K.; Zeginiadou, T.; Pontikides, N. Hypothyroidism has an adverse effect on human spermatogenesis: A prospective, controlled study. Thyroid 2008, 18, 1255–1259. [Google Scholar] [CrossRef] [PubMed]
- Jannini, E.A.; Ulisse, S.; D’Armiento, M. Thyroid hormone and male gonadal function. Endocr. Rev. 1995, 16, 443–459. [Google Scholar]
- Mendis-Handagama, S.M.; Siril Ariyaratne, H.B. Leydig cells, thyroid hormones and steroidogenesis. Indian J. Exp. Biol. 2005, 43, 939–962. [Google Scholar]
- Manna, P.R.; Tena-Sempere, M.; Huhtaniemi, I.T. Molecular mechanisms of thyroid hormone-stimulated steroidogenesis in mouse leydig tumor cells. Involvement of the steroidogenic acute regulatory (star) protein. J. Biol. Chem. 1999, 274, 5909–5918. [Google Scholar] [CrossRef]
- Abalovich, M.; Levalle, O.; Hermes, R.; Scaglia, H.; Aranda, C.; Zylbersztein, C.; Oneto, A.; Aquilano, D.; Gutierrez, S. Hypothalamic-pituitary-testicular axis and seminal parameters in hyperthyroid males. Thyroid 1999, 9, 857–863. [Google Scholar] [CrossRef]
- Kocova, M.; Netkov, S.; Sukarova-Angelovska, E. Pituitary pseudotumor with unusual presentation reversed shortly after the introduction of thyroxine replacement therapy. J. Pediat. Endocrinol. Metab. 2001, 14, 1665–1669. [Google Scholar] [CrossRef]
- Wagner, M.S.; Wajner, S.M.; Maia, A.L. Is there a role for thyroid hormone on spermatogenesis? Microsc. Res. Tech. 2009, 72, 796–808. [Google Scholar] [CrossRef] [PubMed]
- Poppe, K.; Glinoer, D.; Tournaye, H.; Maniewski, U.; Haentjens, P.; Velkeniers, B. Is systematic screening for thyroid disorders indicated in subfertile men? Eur. J. Endocrinol. 2006, 154, 363–366. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.S.; Wajner, S.M.; Maia, A.L. The role of thyroid hormone in testicular development and function. J. Endocrinol. 2008, 199, 351–365. [Google Scholar] [CrossRef]
- Clyde, H.R.; Walsh, P.C.; English, R.W. Elevated plasma testosterone and gonadotropin levels in infertile males with hyperthyroidism. Fertil. Steril. 1976, 27, 662–666. [Google Scholar] [CrossRef]
- Chopra, I.J.; Tulchinsky, D. Status of estrogen-androgen balance in hyperthyroid men with graves’ disease. J. Clin. Endocrinol. Metab. 1974, 38, 269–277. [Google Scholar] [CrossRef]
- Ridgway, E.C.; Maloof, F.; Longcope, C. Androgen and oestrogen dynamics in hyperthyroidism. J. Endocrinol. 1982, 95, 105–115. [Google Scholar] [CrossRef]
- Zähringer, S.; Tomova, A.; Von Werder, K.; Brabant, G.; Kumanov, P.; Schopohl, J. The influence of hyperthyroidism on the hypothalamic-pituitary-gonadal axis. Exp. Clin. Endocrinol. Diab. 2000, 108, 282–289. [Google Scholar] [CrossRef]
- Krassas, G.E.; Pontikides, N.; Deligianni, V.; Miras, K. A prospective controlled study of the impact of hyperthyroidism on reproductive function in males. J. Clin. Endocrinol. Metab. 2002, 87, 3667–3671. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekhar, Y.; D’occhio, M.; Holland, M.; Setchell, B. Activity of the hypothalamo-pituitary axis and testicular development in prepubertal ram lambs with induced hypothyroidism or hyperthyroidism. Endocrinology 1985, 117, 1645–1651. [Google Scholar] [CrossRef]
- Ruder, H.; Corvol, P.; Mahoudeau, J.; Ross, G.; Lipsett, M. Effects of induced hyperthyroidism on steroid metabolism in man. J. Clin. Endocrinol. Metab. 1971, 33, 382–387. [Google Scholar] [CrossRef]
- Kidd, G.S.; Glass, A.R.; Vigersky, R.A. The hypothalamic-pituitary-testicular axis in thyrotoxicosis. J. Clin. Endocrinol. Metab. 1979, 48, 798–802. [Google Scholar] [CrossRef]
- Röjdmark, S.; Berg, A.; Kallner, G. Hypothalamic-pituitary-testicular axis in patients with hyperthyroidism. Horm. Res. 1988, 29, 185–190. [Google Scholar] [PubMed]
- Liu, X.; Tang, C.; Wen, G.; Zhong, C.; Yang, J.; Zhu, J.; Ma, C. The mechanism and pathways of dopamine and dopamine agonists in prolactinomas. Front. Endocrinol. 2019, 9, 768. [Google Scholar] [CrossRef]
- Thorner, M.O.; McNeilly, A.S.; Hagan, C.; Besser, G.M. Long-term treatment of galactorrhoea and hypogonadism with bromocriptine. Brit. Med. J. 1974, 2, 419–422. [Google Scholar] [CrossRef] [PubMed]
- De Rosa, M.; Colao, A.; Di Sarno, A.; Ferone, D.; Landi, M.L.; Zarrilli, S.; Paesano, L.; Merola, B.; Lombardi, G. Cabergoline treatment rapidly improves gonadal function in hyperprolactinemic males: A comparison with bromocriptine. Eur. J. Endocrinol. 1998, 138, 286–293. [Google Scholar] [CrossRef]
- Gillam, M.P.; Molitch, M.E.; Lombardi, G.; Colao, A. Advances in the treatment of prolactinomas. Endocr. Rev. 2006, 27, 485–534. [Google Scholar] [CrossRef]
- Ribeiro, R.S.; Abucham, J. Recovery of persistent hypogonadism by clomiphene in males with prolactinomas under dopamine agonist treatment. Eur. J. Endocrinol. 2009, 161, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Association, A.D. Diagnosis and classification of diabetes mellitus. Diab. Care 2014, 37 (Suppl. 1), S81–S90. [Google Scholar] [CrossRef] [PubMed]
- Dandona, P.; Dhindsa, S. Update: Hypogonadotropic hypogonadism in type 2 diabetes and obesity. J. Clin. Endocrinol. Metab. 2011, 96, 2643–2651. [Google Scholar] [CrossRef] [PubMed]
- Jelodar, G.; Khaksar, Z.; Pourahmadi, M. Endocrine profile and testicular histomorphometry in adult rat offspring of diabetic mothers. J. Physiol. Sci. 2009, 59, 377–382. [Google Scholar] [CrossRef]
- Bener, A.; Al-Ansari, A.A.; Zirie, M.; Al-Hamaq, A.O. Is male fertility associated with type 2 diabetes mellitus? Int. Urol. Nephrol. 2009, 41, 777–784. [Google Scholar] [CrossRef]
- Fedele, D. Therapy insight: Sexual and bladder dysfunction associated with diabetes mellitus. Nat. Clin. Prac. Urol. 2005, 2, 282–290. [Google Scholar] [CrossRef]
- Corona, G.; Giorda, C.B.; Cucinotta, D.; Guida, P.; Nada, E. Sexual dysfunction at the onset of type 2 diabetes: The interplay of depression, hormonal and cardiovascular factors. J. Sex. Med. 2014, 11, 2065–2073. [Google Scholar] [CrossRef]
- Condorelli, R.A.; La Vignera, S.; Mongioì, L.M.; Alamo, A.; Calogero, A.E. Diabetes mellitus and infertility: Different pathophysiological effects in type 1 and type 2 on sperm function. Front. Endocrinol. 2018, 9, 268. [Google Scholar] [CrossRef]
- Carpino, A.; Rago, V.; Guido, C.; Casaburi, I.; Aquila, S. Insulin and ir-beta in pig spermatozoa: A role of the hormone in the acquisition of fertilizing ability. Int. J. Androl. 2010, 33, 554–562. [Google Scholar] [CrossRef]
- Scarano, W.R.; Messias, A.G.; Oliva, S.U.; Klinefelter, G.R.; Kempinas, W.G. Sexual behaviour, sperm quantity and quality after short-term streptozotocin-induced hyperglycaemia in rats. Int. J. Androl. 2006, 29, 482–488. [Google Scholar] [CrossRef]
- Baccetti, B.; La Marca, A.; Piomboni, P.; Capitani, S.; Bruni, E.; Petraglia, F.; De Leo, V. Insulin-dependent diabetes in men is associated with hypothalamo-pituitary derangement and with impairment in semen quality. Hum. Reprod. 2002, 17, 2673–2677. [Google Scholar] [CrossRef]
- Yu, T.; Robotham, J.L.; Yoon, Y. Increased production of reactive oxygen species in hyperglycemic conditions requires dynamic change of mitochondrial morphology. Proc. Nat. Acad. Sci. USA 2006, 103, 2653–2658. [Google Scholar] [CrossRef] [PubMed]
- Mangoli, E.; Talebi, A.R.; Anvari, M.; Pourentezari, M. Effects of experimentally-induced diabetes on sperm parameters and chromatin quality in mice. Iran. J. Reprod. Med. 2013, 11, 53–60. [Google Scholar]
- Navarro-Casado, L.; Juncos-Tobarra, M.A.; Cháfer-Rudilla, M.; de Onzoño, L.; Blázquez-Cabrera, J.A.; Miralles-García, J.M. Effect of experimental diabetes and stz on male fertility capacity. Study in rats. J. Androl. 2010, 31, 584–592. [Google Scholar] [CrossRef]
- Mora-Esteves, C.; Shin, D. Nutrient supplementation: Improving male fertility fourfold. Sem. Reprod. Med. 2013, 31, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Malini, T.; Rengarajan, S.; Balasubramanian, K. Impact of experimental diabetes and insulin replacement on epididymal secretory products and sperm maturation in albino rats. J. Cell. Biochem. 2009, 108, 1094–1101. [Google Scholar] [CrossRef]
- Steger, R.W.; Rabe, M.B. The effect of diabetes mellitus on endocrine and reproductive function. Proc. Soc. Exp. Biol. Med. 1997, 214, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Urner, F.; Sakkas, D. Glucose participates in sperm-oocyte fusion in the mouse. Biol. Reprod. 1996, 55, 917–922. [Google Scholar] [CrossRef]
- Lampiao, F.; Du Plessis, S.S. Insulin stimulates glut8 expression in human spermatozoa. J. Biosci. Tech. 2010, 1, 90–93. [Google Scholar]
- Scheepers, A.; Joost, H.G.; Schürmann, A. The glucose transporter families sglt and glut: Molecular basis of normal and aberrant function. J. Parent. Ent. Nutr. 2004, 28, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Schürmann, A.; Axer, H.; Scheepers, A.; Doege, H.; Joost, H.G. The glucose transport facilitator glut8 is predominantly associated with the acrosomal region of mature spermatozoa. Cell Tissue Res. 2002, 307, 237–242. [Google Scholar] [CrossRef]
- Gómez, O.; Romero, A.; Terrado, J.; Mesonero, J.E. Differential expression of glucose transporter glut8 during mouse spermatogenesis. Reproduction 2006, 131, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Bucci, D.; Rodriguez-Gil, J.E.; Vallorani, C.; Spinaci, M.; Galeati, G.; Tamanini, C. Gluts and mammalian sperm metabolism. J. Androl. 2011, 32, 348–355. [Google Scholar] [CrossRef]
- Kim, S.T.; Moley, K.H. The expression of glut8, glut9a, and glut9b in the mouse testis and sperm. Reprod. Sci. 2007, 14, 445–455. [Google Scholar]
- Gawlik, V.; Schmidt, S.; Scheepers, A.; Wennemuth, G.; Augustin, R.; Aumüller, G.; Moser, M.; Al-Hasani, H.; Kluge, R.; Joost, H.G.; et al. Targeted disruption of slc2a8 (glut8) reduces motility and mitochondrial potential of spermatozoa. Mol. Memb. Biol. 2008, 25, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, K.; Sengupta, P.; Dutta, S.; Karkada, I.R. Obesity, systemic inflammation and male infertility. Chem. Biol. Lett. 2020, 7, 92–98. [Google Scholar]
- Dutta, S.; Biswas, A.; Sengupta, P. Obesity, endocrine disruption and male infertility. Asian Pac. J. Reprod. 2019, 8, 195–202. [Google Scholar] [CrossRef]
- Jia, Y.F.; Feng, Q.; Ge, Z.Y.; Guo, Y.; Zhou, F.; Zhang, K.S.; Wang, X.W.; Lu, W.H.; Liang, X.W.; Gu, Y.Q. Obesity impairs male fertility through long-term effects on spermatogenesis. BMC Urol. 2018, 18, 42. [Google Scholar] [CrossRef]
- Bessesen, D.; Hill, J.; Wyatt, H. Hormones and obesity. J. Clin. Endocrinol. Metab. 2004, 89. [Google Scholar] [CrossRef]
- de Oliveira Leal, V.; Mafra, D. Adipokines in obesity. Clin. Chim. Acta 2013, 419, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Tsai, E.C.; Matsumoto, A.M.; Fujimoto, W.Y.; Boyko, E.J. Association of bioavailable, free, and total testosterone with insulin resistance: Influence of sex hormone-binding globulin and body fat. Diab. Care 2004, 27, 861–868. [Google Scholar] [CrossRef]
- Davidson, L.M.; Millar, K.; Jones, C.; Fatum, M.; Coward, K. Deleterious effects of obesity upon the hormonal and molecular mechanisms controlling spermatogenesis and male fertility. Hum. Fertil. 2015, 18, 184–193. [Google Scholar] [CrossRef]
- Katib, A. Mechanisms linking obesity to male infertility. Centr. Eur. J. Urol. 2015, 68, 79. [Google Scholar] [CrossRef]
- Hammoud, A.O.; Gibson, M.; Peterson, C.M.; Hamilton, B.D.; Carrell, D.T. Obesity and male reproductive potential. J. Androl. 2006, 27, 619–626. [Google Scholar] [CrossRef]
- Schulster, M.; Bernie, A.M.; Ramasamy, R. The role of estradiol in male reproductive function. Asian J. Androl. 2016, 18, 435. [Google Scholar] [PubMed]
- Chimento, A.; Sirianni, R.; Casaburi, I.; Pezzi, V. Role of estrogen receptors and g protein-coupled estrogen receptor in regulation of hypothalamus–pituitary–testis axis and spermatogenesis. Front. Endocrinol. 2014, 5, 1. [Google Scholar] [CrossRef]
- Salazar, M.; Sánchez, J.; Álvarez, P.; Frusch, J.; Mejía, R. The impact of obesity on fertility. J. Reprod. Med. Gynecol. Obstet. 2018, 3, 9. [Google Scholar]
- Kim, J.Y.; Van De Wall, E.; Laplante, M.; Azzara, A.; Trujillo, M.E.; Hofmann, S.M.; Schraw, T.; Durand, J.L.; Li, H.; Li, G. Obesity-associated improvements in metabolic profile through expansion of adipose tissue. J. Clin. Investig. 2007, 117, 2621–2637. [Google Scholar] [CrossRef] [PubMed]
- Boden, G. Obesity and free fatty acids. Endocrinol. Metab. Clin. N. Am. 2008, 37, 635–646. [Google Scholar] [CrossRef]
- Sengupta, P.; Bhattacharya, K.; Dutta, S. Leptin and male reproduction. Asian Pac. J. Reprod. 2019, 8, 220–226. [Google Scholar] [CrossRef]
- Dutta, S.; Biswas, A.; Sengupta, P.; Nwagha, U. Ghrelin and male reproduction. Asian Pac. J.Reprod. 2019, 8, 227–232. [Google Scholar] [CrossRef]
- Sengupta, P.; Dutta, S.; Tusimin, M.; Karkada, I.R. Orexins and male reproduction. Asian Pac. J. Reprod. 2019, 8, 233. [Google Scholar] [CrossRef]
- İrez, T.; Karkada, I.R.; Dutta, S.; Sengupta, P. Obestatin in male reproduction and infertility. Asian Pac. J. Reprod. 2019, 8, 239–243. [Google Scholar]
- Dutta, S.; Sengupta, P.; Chakravarthi, S.; Irez, T.; Baktir, G. Adiponectin: ‘A metabolic ballcock’modulating immune responses and male reproduction. Chem. Biol. Lett. 2021, 8, 171–182. [Google Scholar]
- Morton, G.; Cummings, D.; Baskin, D.; Barsh, G.; Schwartz, M. Central nervous system control of food intake and body weight. Nature 2006, 443, 289. [Google Scholar] [CrossRef] [PubMed]
- Mounzih, K.; Lu, R.; Chehab, F.F. Leptin treatment rescues the sterility of genetically obese ob/ob males. Endocrinology 1997, 138, 1190–1193. [Google Scholar] [CrossRef] [PubMed]
- Bhat, G.K.; Sea, T.L.; Olatinwo, M.O.; Simorangkir, D.; Ford, G.D.; Ford, B.D.; Mann, D.R. Influence of a leptin deficiency on testicular morphology, germ cell apoptosis, and expression levels of apoptosis-related genes in the mouse. J. Androl. 2006, 27, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Ramos, C.F.; Zamoner, A. Thyroid hormone and leptin in the testis. Front. Endocrinol. 2014, 5, 198. [Google Scholar] [CrossRef]
- Bouloumie, A.; Marumo, T.; Lafontan, M.; Busse, R. Leptin induces oxidative stress in human endothelial cells. FASEB J. 1999, 13, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, S.I.; Edelstein, D.; Du, X.L.; Kaneda, Y.; Guzman, M.; Brownlee, M. Leptin induces mitochondrial superoxide production and monocyte chemoattractant protein-1 expression in aortic endothelial cells by increasing fatty acid oxidation via protein kinase a. J. Biol. Chem. 2001, 276, 25096–25100. [Google Scholar] [CrossRef] [PubMed]
- Wauters, M.; Considine, R.V.; Van Gaal, L.F. Human leptin: From an adipocyte hormone to an endocrine mediator. Eur. J. Endocrinol. 2000, 143, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Page, S.T.; Herbst, K.L.; Amory, J.K.; Coviello, A.D.; Anawalt, B.D.; Matsumoto, A.M.; Bremner, W.J. Testosterone administration suppresses adiponectin levels in men. J. Androl. 2005, 26, 85–92. [Google Scholar] [PubMed]
- Yuan, F.; Li, Y.N.; Liu, Y.H.; Yi, B.; Tian, J.W.; Liu, F.Y. Adiponectin inhibits the generation of reactive oxygen species induced by high glucose and promotes endothelial no synthase formation in human mesangial cells. Mol. Med. Rep. 2012, 6, 449–453. [Google Scholar] [CrossRef]
- Wolfe, A.; Hussain, M.A. The emerging role(s) for kisspeptin in metabolism in mammals. Front. Endocrinol. 2018, 9, 184. [Google Scholar] [CrossRef]
- Clarke, H.; Dhillo, W.S.; Jayasena, C.N. Comprehensive review on kisspeptin and its role in reproductive disorders. Endocrinol. Metab. 2015, 30, 124–141. [Google Scholar] [CrossRef]
- Zheng, D.; Zhao, Y.; Shen, Y.; Chang, X.; Ju, S.; Guo, L. Orexin a-mediated stimulation of 3β-hsd expression and testosterone production through mapk signaling pathways in primary rat leydig cells. J. Endocrinol. Investig. 2014, 37, 285–292. [Google Scholar] [CrossRef]
- Duffy, C.M.; Nixon, J.P.; Butterick, T.A. Orexin a attenuates palmitic acid-induced hypothalamic cell death. Mol. Cell. Neurosci. 2016, 75, 93–100. [Google Scholar] [CrossRef]
- Sengupta, P.; Hassan, M.F.; Dutta, S.; Jegasothy, R.; Chhikara, B.S. Orexins: The ‘multitasking’neuropeptides in the energy metabolism and immune regulation of male reproduction. Chem. Biol. Lett. 2021, 8, 202–212. [Google Scholar]
- Trayhurn, P.; Beattie, J.H. Physiological role of adipose tissue: White adipose tissue as an endocrine and secretory organ. Proc. Nutr. Soc. 2001, 60, 329–339. [Google Scholar] [CrossRef]
- Flehmig, G.; Scholz, M.; Klöting, N.; Fasshauer, M.; Tönjes, A.; Stumvoll, M.; Youn, B.-S.; Blüher, M. Identification of adipokine clusters related to parameters of fat mass, insulin sensitivity and inflammation. PLoS ONE 2014, 9, e99785. [Google Scholar]
- Dutta, S.; Sengupta, P.; Jegasothy, R.; Akhigbe, R. Resistin and visfatin: ‘Connecting threads’ of immunity, energy modulations and male reproduction. Chem. Biol. Lett. 2021, 8, 192–201. [Google Scholar]
- Irez, T.; Bicer, S.; Sahin, E.; Dutta, S.; Sengupta, P. Cytokines and adipokines in the regulation of spermatogenesis and semen quality. Chem. Biol. Lett. 2020, 7, 131–139. [Google Scholar]
- Du Plessis, S.S.; Cabler, S.; McAlister, D.A.; Sabanegh, E.; Agarwal, A. The effect of obesity on sperm disorders and male infertility. Nat. Rev. Urol. 2010, 7, 153. [Google Scholar] [CrossRef]
- Ishikawa, T.; Fujioka, H.; Ishimura, T.; Takenaka, A.; Fujisawa, M. Ghrelin expression in human testis and serum testosterone level. J. Androl. 2007, 28, 320–324. [Google Scholar] [CrossRef]
- Wang, L.; Fang, F.; Li, Y.; Zhang, Y.; Pu, Y.; Zhang, X. Role of ghrelin on testosterone secretion and the mrna expression of androgen receptors in adult rat testis. Syst. Biol. Reprod. Med. 2011, 57, 119–123. [Google Scholar] [CrossRef]
- Greenman, Y.; Rouach, V.; Limor, R.; Gilad, S.; Stern, N. Testosterone is a strong correlate of ghrelin levels in men and postmenopausal women. Neuroendocrinology 2009, 89, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Matsuzaki, J.; Hibi, T. Ghrelin and oxidative stress in gastrointestinal tract. J. Clin. Biochem. Nutr. 2010, 48, 122–125. [Google Scholar] [CrossRef]
- Ahima, R.S. Metabolic actions of adipocyte hormones: Focus on adiponectin. Obesity 2006, 14, 9S–15S. [Google Scholar] [CrossRef] [PubMed]
- Polyzos, S.A.; Kountouras, J.; Zavos, C.; Tsiaousi, E. The role of adiponectin in the pathogenesis and treatment of non-alcoholic fatty liver disease. Diabetes Obes. Metab. 2010, 12, 365–383. [Google Scholar] [CrossRef] [PubMed]
- Seeger, J.; Ziegelmeier, M.; Bachmann, A.; Lossner, U.; Kratzsch, J.; Bluher, M.; Stumvoll, M.; Fasshauer, M. Serum levels of the adipokine vaspin in relation to metabolic and renal parameters. J. Clin. Endocrinol. Metab. 2008, 93, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Kurowska, P.; Barbe, A.; Różycka, M.; Chmielińska, J.; Dupont, J.; Rak, A. Apelin in reproductive physiology and pathology of different species: A critical review. Int. J. Endocrinol. 2018, 2018, 9170480. [Google Scholar] [CrossRef]
- Ismail, S.A.; Mahran, A.M.; Mosaad, E.; ElSayed, S.K. Omentin-1 in serum and seminal plasma correlate with semen quality. Hum. Androl. 2017, 7, 120–126. [Google Scholar] [CrossRef][Green Version]
- Liew, F.F.; Dutta, S.; Sengupta, P.; Chhikara, B.S. Chemerin and male reproduction: ‘A tangled rope’connecting metabolism and inflammation. Chem. Biol. Lett. 2021, 8, 224–237. [Google Scholar]
- Sengupta, P.; Dutta, S.; Karkada, I.R.; Akhigbe, R.E.; Chinni, S.V. Irisin, energy homeostasis and male reproduction. Front. Physiol. 2021, 12, 746049. [Google Scholar] [CrossRef]
- Low, S.; Chin, M.C.; Deurenberg-Yap, M. Review on epidemic of obesity. Ann. Acad. Med. Singap. 2009, 38, 57. [Google Scholar]
- De Rooij, D.; Van Alphen, M.; Van de Kant, H. Duration of the cycle of the seminiferous epithelium and its stages in the rhesus monkey (macaca mulatta). Biol. Reprod. 1986, 35, 587–591. [Google Scholar] [CrossRef]
- Hikim, A.S.; Swerdloff, R.S. Hormonal and genetic control of germ cell apoptosis in the testis. Rev. Reprod. 1999, 4, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, I.; Ody, C.; Araki, K.; Garcia, I.; Vassalli, P. An early and massive wave of germinal cell apoptosis is required for the development of functional spermatogenesis. EMBO J. 1997, 16, 2262–2270. [Google Scholar] [CrossRef]
- Garolla, A.; Torino, M.; Sartini, B.; Cosci, I.; Patassini, C.; Carraro, U.; Foresta, C. Seminal and molecular evidence that sauna exposure affects human spermatogenesis. Hum. Reprod. 2013, 28, 877–885. [Google Scholar] [CrossRef]
- Xin, W.; Li, X.; Lu, X.; Niu, K.; Cai, J. Involvement of endoplasmic reticulum stress-associated apoptosis in a heart failure model induced by chronic myocardial ischemia. Int. J. Mol. Med. 2011, 27, 503–509. [Google Scholar]
- Li, C.; Dong, Z.; Lan, X.; Zhang, X.; Li, S. Endoplasmic reticulum stress promotes the apoptosis of testicular germ cells in hyperlipidemic rats. Nat. J. Androl. 2015, 21, 402–407. [Google Scholar]
- Sengupta, P. Current trends of male reproductive health disorders and the changing semen quality. Int. J. Prev. Med. 2014, 5, 1–5. [Google Scholar] [PubMed]
- Sengupta, P. Recent trends in male reproductive health problems. Asian J. Pharm. Clin. Res. 2014, 7, 1–5. [Google Scholar]
- Sermondade, N.; Dupont, C.; Faure, C.; Boubaya, M.; Cédrin-Durnerin, I.; Chavatte-Palmer, P.; Sifer, C.; Lévy, R. Body mass index is not associated with sperm–zona pellucida binding ability in subfertile males. Asian J. Androl. 2013, 15, 626. [Google Scholar] [CrossRef] [PubMed]
- Håkonsen, L.B.; Thulstrup, A.M.; Aggerholm, A.S.; Olsen, J.; Bonde, J.P.; Andersen, C.Y.; Bungum, M.; Ernst, E.H.; Hansen, M.L.; Ernst, E.H. Does weight loss improve semen quality and reproductive hormones? Results from a cohort of severely obese men. Reprod. Health 2011, 8, 24. [Google Scholar] [CrossRef]
- Bieniek, J.M.; Kashanian, J.A.; Deibert, C.M.; Grober, E.D.; Lo, K.C.; Brannigan, R.E.; Sandlow, J.I.; Jarvi, K.A. Influence of increasing body mass index on semen and reproductive hormonal parameters in a multi-institutional cohort of subfertile men. Fertil. Steril. 2016, 106, 1070–1075. [Google Scholar] [CrossRef]
- Chavarro, J.E.; Toth, T.L.; Wright, D.L.; Meeker, J.D.; Hauser, R. Body mass index in relation to semen quality, sperm DNA integrity, and serum reproductive hormone levels among men attending an infertility clinic. Fertil. Steril. 2010, 93, 2222–2231. [Google Scholar] [CrossRef]
- Jensen, T.K.; Andersson, A.M.; Jørgensen, N.; Andersen, A.G.; Carlsen, E.; Skakkebæk, N.E. Body mass index in relation to semen quality and reproductive hormonesamong 1,558 danish men. Fertil. Steril. 2004, 82, 863–870. [Google Scholar] [CrossRef]
- Kahn, B.E.; Brannigan, R.E. Obesity and male infertility. Curr. Opin. Urol. 2017, 27, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, A.; Stewart, A.; Farquhar, C. Body mass index in relation to semen quality and reproductive hormones in new zealand men: A cross-sectional study in fertility clinics. Hum. Reprod. 2013, 28, 3178–3187. [Google Scholar] [CrossRef]
- Darbandi, S.; Darbandi, M.; Mokarram, P.; Sadeghi, M.R.; Owji, A.A.; Khorram Khorshid, H.R.; Zhao, B.; Heidari, M. Cme article: The acupuncture-affected gene expressions and epigenetic modifications in oxidative stress–associated diseases. Med. Acup. 2016, 28, 16–27. [Google Scholar] [CrossRef]
- Agarwal, A.; Mulgund, A.; Alshahrani, S.; Assidi, M.; Abuzenadah, A.M.; Sharma, R.; Sabanegh, E. Reactive oxygen species and sperm DNA damage in infertile men presenting with low level leukocytospermia. Reprod. Biol. Endocrinol. 2014, 12, 126. [Google Scholar] [CrossRef] [PubMed]
- Zirkin, B.R.; Chen, H. Regulation of leydig cell steroidogenic function during aging. Biol. Reprod. 2000, 63, 977–981. [Google Scholar] [CrossRef]
- Turner, T.T.; Bang, H.J.; Lysiak, J.J. Experimental testicular torsion: Reperfusion blood flow and subsequent testicular venous plasma testosterone concentrations. Urology 2005, 65, 390–394. [Google Scholar] [CrossRef]
- Hanukoglu, I. Antioxidant protective mechanisms against reactive oxygen species (ros) generated by mitochondrial p450 systems in steroidogenic cells. Drug Metab. Rev. 2006, 38, 171–196. [Google Scholar] [CrossRef]
- Luo, L.; Chen, H.; Trush, M.A.; Show, M.D.; Anway, M.D.; Zirkin, B.R. Aging and the brown norway rat leydig cell antioxidant defense system. J. Androl. 2006, 27, 240–247. [Google Scholar] [CrossRef]
- Agarwal, A.; Said, T.M. Role of sperm chromatin abnormalities and DNA damage in male infertility. Hum. Reprod. Update 2003, 9, 331–345. [Google Scholar] [CrossRef]
- Aitken, R.J.; Baker, M.A.; Sawyer, D. Oxidative stress in the male germ line and its role in the aetiology of male infertility and genetic disease. Reprod. Biomed. Online 2003, 7, 65–70. [Google Scholar] [CrossRef]
- Said, T.M.; Paasch, U.; Glander, H.J.; Agarwal, A. Role of caspases in male infertility. Hum. Reprod. Update 2004, 10, 39–51. [Google Scholar] [CrossRef]
- Gil-Guzman, E.; Ollero, M.; Lopez, M.; Sharma, R.; Alvarez, J.; Thomas, A.; Agarwal, A. Differential production of reactive oxygen species by subsets of human spermatozoa at different stages of maturation. Hum. Reprod. 2001, 16, 1922–1930. [Google Scholar] [CrossRef] [PubMed]
- Mancini, A.; Festa, R.; Silvestrini, A.; Nicolotti, N.; Di Donna, V.; La Torre, G.; Pontecorvi, A.; Meucci, E. Hormonal regulation of total antioxidant capacity in seminal plasma. J. Androl. 2009, 30, 534–540. [Google Scholar] [CrossRef]
- Chainy, G.; Samantaray, S.; Samanta, L. Testosterone-induced changes in testicular antioxidant system. Andrologia 1997, 29, 343–349. [Google Scholar] [CrossRef]
- Shang, X.; Huang, Y.; Ye, Z.; Yu, X.; Gu, W. Protection of melatonin against damage of sperm mitochondrial function induced by reactive oxygen species. Nat. J. Androl. 2004, 10, 604–607. [Google Scholar]
- Lakpour, N.; Mahfouz, R.Z.; Akhondi, M.M.; Agarwal, A.; Kharrazi, H.; Zeraati, H.; Amirjannati, N.; Sadeghi, M.R. Relationship of seminal plasma antioxidants and serum male hormones with sperm chromatin status in male factor infertility. Syst. Biol. Reprod. Med. 2012, 58, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Oluboyo, A.; Adijeh, R.; Onyenekwe, C.; Oluboyo, B.; Mbaeri, T.; Odiegwu, C.; Chukwuma, G.; Onwuasoanya, U. Relationship between serum levels of testosterone, zinc and selenium in infertile males attending fertility clinic in nnewi, south east nigeria. Afr. J. Med. Med. Sci. 2012, 41, 51–54. [Google Scholar] [PubMed]
- Mancini, A.; Leone, E.; Festa, R.; Grande, G.; Silvestrini, A.; Marinis, L.; Pontecorvi, A.; Maira, G.; Littarru, G.P.; Meucci, E. Effects of testosterone on antioxidant systems in male secondary hypogonadism. J. Androl. 2008, 29, 622–629. [Google Scholar] [CrossRef]
- Safarinejad, M.R. Efficacy of coenzyme q10 on semen parameters, sperm function and reproductive hormones in infertile men. J. Urol. 2009, 182, 237–248. [Google Scholar] [CrossRef]
- Richthoff, J.; Spano, M.; Giwercman, Y.; Frohm, B.; Jepson, K.; Malm, J.; Elzanaty, S.; Stridsberg, M.; Giwercman, A. The impact of testicular and accessory sex gland function on sperm chromatin integrity as assessed by the sperm chromatin structure assay (scsa). Hum. Reprod. 2002, 17, 3162–3169. [Google Scholar] [CrossRef]
- Meeker, J.D.; Singh, N.P.; Hauser, R. Serum concentrations of estradiol and free t4 are inversely correlated with sperm DNA damage in men from an infertility clinic. J. Androl. 2008, 29, 379–388. [Google Scholar] [CrossRef]
- Ahmadi, S.; Bashiri, R.; Ghadiri-Anari, A.; Nadjarzadeh, A. Antioxidant supplements and semen parameters: An evidence based review. Int. J. Reprod. BioMed. 2016, 14, 729. [Google Scholar] [CrossRef]
- Darbandi, M.; Darbandi, S.; Agarwal, A.; Sengupta, P.; Durairajanayagam, D.; Henkel, R.; Sadeghi, M.R. Reactive oxygen species and male reproductive hormones. Reprod. Biol. Endocrinol. 2018, 16, 87. [Google Scholar] [CrossRef] [PubMed]
- Palomba, S.; Falbo, A.; Espinola, S.; Rocca, M.; Capasso, S.; Cappiello, F.; Zullo, F. Effects of highly purified follicle-stimulating hormone on sperm DNA damage in men with male idiopathic subfertility: A pilot study. J. Endocrinol. Investig. 2011, 34, 747–752. [Google Scholar]
- Colacurci, N.; Monti, M.G.; Fornaro, F.; Izzo, G.; Izzo, P.; Trotta, C.; Mele, D.; Franciscis, P. Recombinant human fsh reduces sperm DNA fragmentation in men with idiopathic oligoasthenoteratozoospermia. J. Androl. 2012, 33, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Tesarik, J.; Martinez, F.; Rienzi, L.; Iacobelli, M.; Ubaldi, F.; Mendoza, C.; Greco, E. In-vitro effects of fsh and testosterone withdrawal on caspase activation and DNA fragmentation in different cell types of human seminiferous epithelium. Hum. Reprod. 2002, 17, 1811–1819. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nematollahi-Mahani, S.N.; Azizollahi, G.H.; Baneshi, M.R.; Safari, Z.; Azizollahi, S. Effect of folic acid and zinc sulphate on endocrine parameters and seminal antioxidant level after varicocelectomy. Andrologia 2014, 46, 240–245. [Google Scholar] [CrossRef]


{kind=link}
| Endocrinopathy | Changes in Male Reproduction | Study |
|---|---|---|
| Hypogonadotropic hypogonadism (Genetic: Kallman syndrome) | Delayed puberty and infertility caused by a malfunction of GnRH-secreting neurons to migrate; cessation of gonadotropin secretion | [12,14] |
| Hypergonadotropic hypogonadism | Increased FSH/LH, normal or ↓testis volume, decreased pubic hair and penis size, infertility | [15] |
| Androgen excess | Inhibition to GnRH secretion, normal or ↓FSH, ↓LH, | [16,17] |
| Estrogen excess | ↓T:E2, ↓semen parameters | [18,19] |
| Hyperprolactinemia | Normal or ↓FSH/LH, ↓testosterone | [20,21,22,23] |
| Insulin disorders | ↓spermatogenesis, ↓reduced vacuolization in the Sertoli cells, ↓fertility, ↓semen parameters, ↓Leydig cells count, ↓testosterone | [24,25,26,27,28] |
| Hypothyroidism | Hyperthyroidism | References | |
|---|---|---|---|
| Prepubertal testicular volume and function | ↑ Early onset of spermatogenesis | ↓ | [75,76,77] |
| Sperm count | Normal or ↓ | ↓ | [78,79] |
| Testicular germ cell count | ↓ | [63,64,71] | |
| Sperm motility | ↓ | ↓ | [80,81] |
| Sexual function | Impaired | Impaired; precocious ejaculation | [73,79,81,82] |
| Erectile function | ↓ | ↓ | [73,82,83] |
| Free testosterone level | ↓ | ↓ | [68] |
| LH and FSH levels | ↓ | ↑ and SHBG | [69] |
| E2 | ↓ | ↑ | [84,85,86] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sengupta, P.; Dutta, S.; Karkada, I.R.; Chinni, S.V. Endocrinopathies and Male Infertility. Life 2022, 12, 10. https://doi.org/10.3390/life12010010
Sengupta P, Dutta S, Karkada IR, Chinni SV. Endocrinopathies and Male Infertility. Life. 2022; 12(1):10. https://doi.org/10.3390/life12010010
Chicago/Turabian StyleSengupta, Pallav, Sulagna Dutta, Ivan Rolland Karkada, and Suresh V. Chinni. 2022. "Endocrinopathies and Male Infertility" Life 12, no. 1: 10. https://doi.org/10.3390/life12010010
APA StyleSengupta, P., Dutta, S., Karkada, I. R., & Chinni, S. V. (2022). Endocrinopathies and Male Infertility. Life, 12(1), 10. https://doi.org/10.3390/life12010010