Prenatal Visual Exposure to a Predator Influences Lateralization in Goldbelly Topminnows




Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Mirror Test
2.2. Rotational Preference in a Familiar Environment
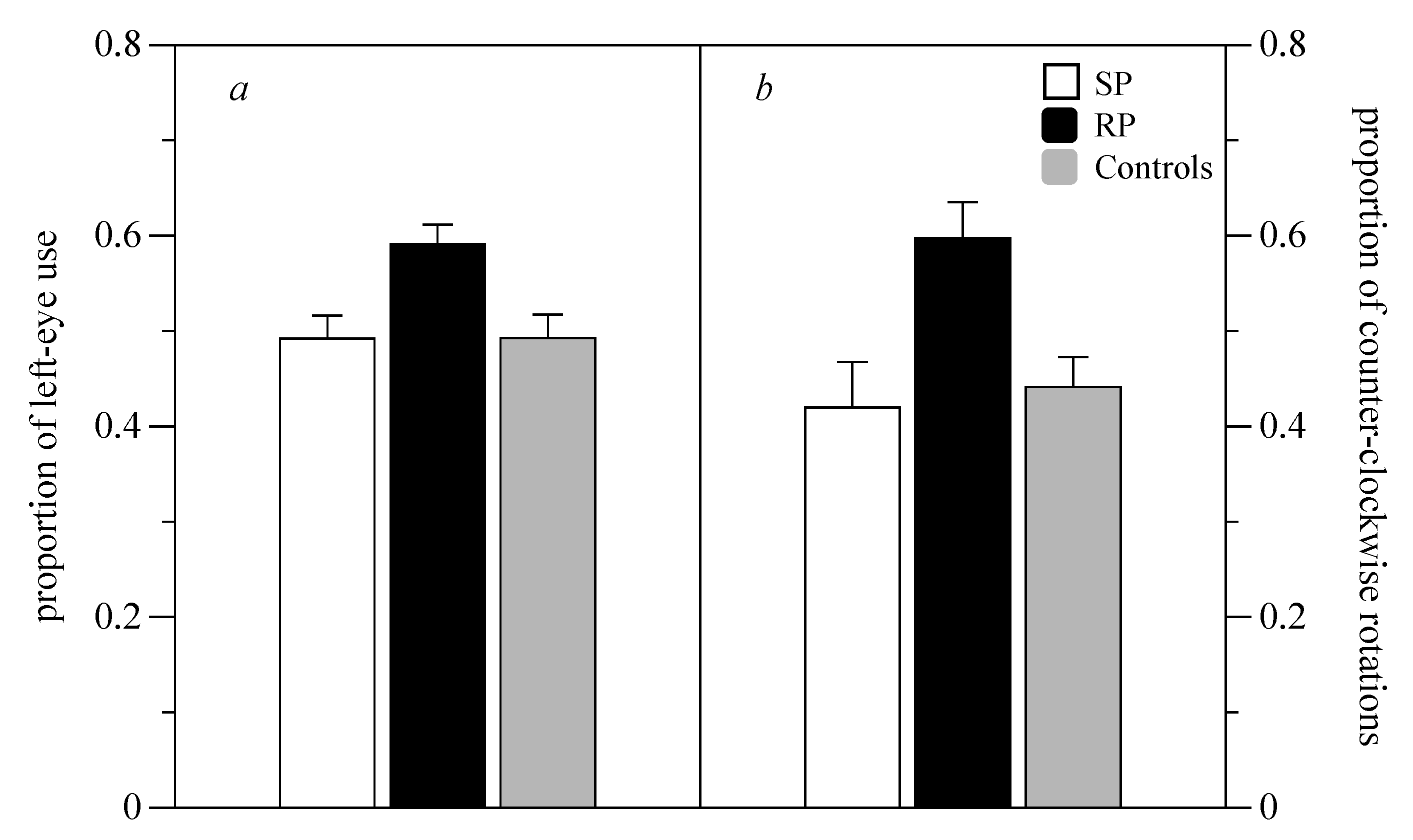
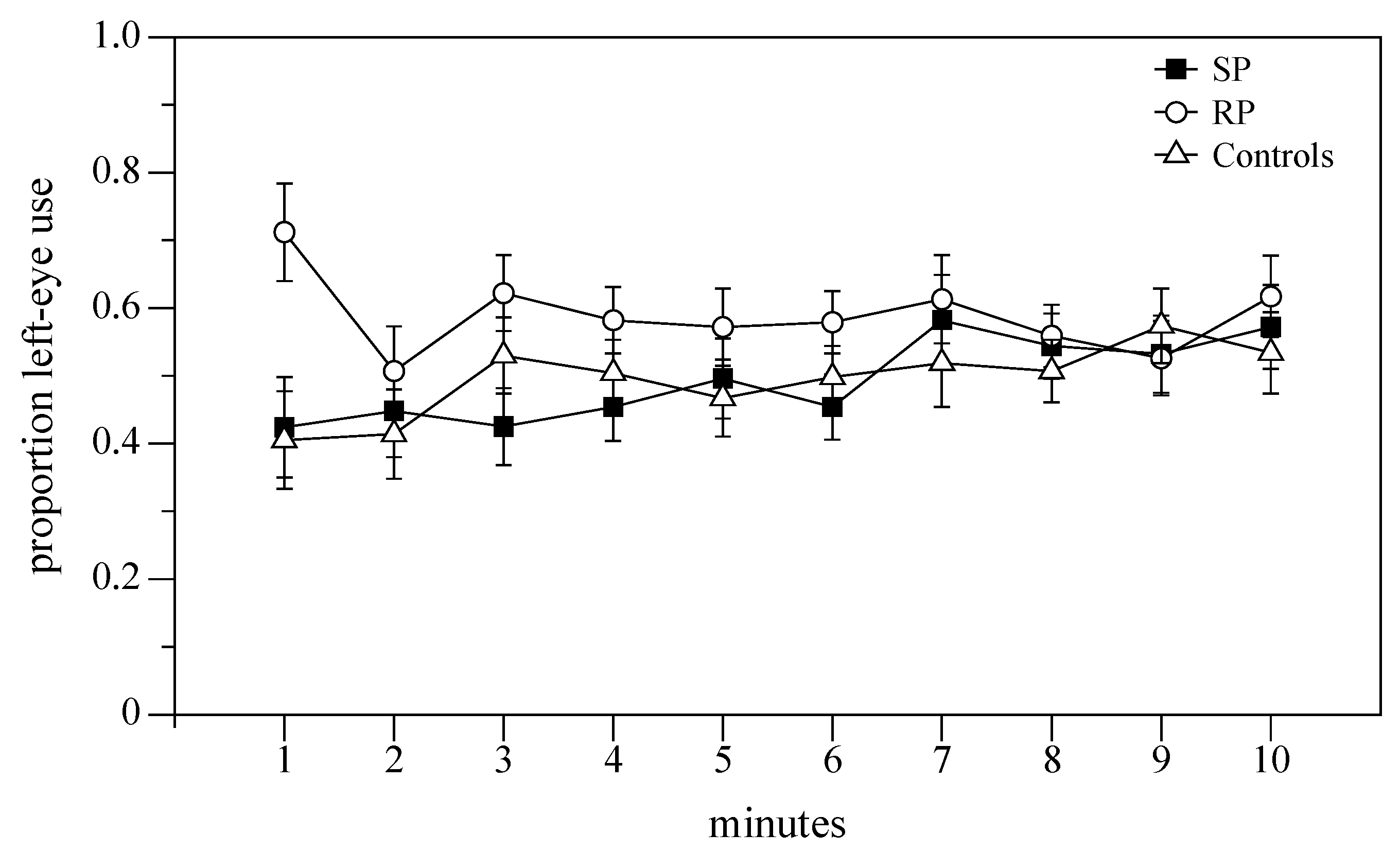
3. Results
3.1. Mirror Test
3.2. Rotational Preference in a Familiar Environment
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Frasnelli, E.; Vallortigara, G.; Rogers, L.J. Left–right asymmetries of behaviour and nervous system in invertebrates. Neurosci. Biobehav. Rev. 2012, 36, 1273–1291. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.J. Lateralization in its many forms, and its evolution and development. Spec. Top. Primatol. 2007, 5, 22–56. [Google Scholar]
- Dadda, M.; Bisazza, A. Does brain asymmetry allow efficient performance of simultaneous tasks? Anim. Behav. 2006, 72, 523–529. [Google Scholar] [CrossRef]
- Rogers, L.J.; Zucca, P.; Vallortigara, G. Advantages of having a lateralized brain. Proc. R. Soc. B Biol. Sci. 2004, 271, S420–S422. [Google Scholar] [CrossRef]
- Dadda, M.; Zandonà, E.; Agrillo, C.; Bisazza, A. The costs of hemispheric specialization in a fish. Proc. R. Soc. B Biol. Sci. 2009, 276, 4399–4407. [Google Scholar] [CrossRef]
- Brown, C.; Gardner, C.; Braithwaite, V.A. Population variation in lateralized eye use in the poeciliid Brachyraphis episcopi. Proc. R. Soc. B Biol. Sci. 2004, 271, S455–S457. [Google Scholar] [CrossRef]
- Ghirlanda, S.; Vallortigara, G. The evolution of brain lateralization: A game-theoretical analysis of population structure. Proc. R. Soc. B Biol. Sci. 2004, 271, 853–857. [Google Scholar] [CrossRef]
- Bisazza, A.; Dadda, M.; Facchin, L.; Vigo, F. Artificial selection on laterality in the teleost fish. Behav. Brain Res. 2007, 178, 29–38. [Google Scholar] [CrossRef]
- Dadda, M.; Domenichini, A.; Piffer, L.; Argenton, F.; Bisazza, A. Early differences in epithalamic left-right asymmetry influence lateralization and personality of adult zebrafish. Behav. Brain Res. 2010, 206, 208–215. [Google Scholar] [CrossRef]
- Hopkins, W.D.; Dahl, J.F.; Pilcher, D. Genetic influence on the expression of hand preferences in Chimpanzees (Pan troglodytes): Evidence in Support of the Right-Shift Theory and Developmental Instability. Psychol. Sci. 2001, 12, 299–303. [Google Scholar] [CrossRef]
- Vallortigara, G.; Versace, E. Laterality at the Neural, Cognitive, and Behavioral Levels. In APA Handbook of Comparative Psychology: Basic Concepts, Methods, Neural Substrate, and Behavior; Call, J., Burghardt, G., Pepperberg, I.M., Snowdon, C.T., Zentall, T., Eds.; American Psychological Association: Washington, DC, USA, 2017; Volume 1, pp. 557–577. [Google Scholar]
- Cowell, P.; Waters, N.; Denenberg, V. The effects of early environment on the development of functional laterality in Morris maze performance. Laterality Asymmetries Body Brain Cogn. 1997, 2, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Gadea, M.; Gomez, C.; González-Bono, E.; Salvador, A.; Espert, R. Salivary testosterone is related to both handedness and degree of linguistic lateralization in normal women. Psychoneuroendocrinology 2003, 28, 274–287. [Google Scholar] [CrossRef]
- Williams, H.; Crane, L.A.; Hale, T.K.; Esposito, M.A.; Nottebohm, F. Right-side dominance for song control in the zebra finch. J. Neurobiol. 1992, 23, 1006–1020. [Google Scholar] [CrossRef]
- Bibost, A.-L.; Kydd, E.; Brown, C. The Effect of Sex and Early Environment on the Lateralization of the Rainbowfish Melanotaenia duboulayi. In Behavioral Lateralization in Vertebrates: Two Sides of the Same Coin; Csermely, D., Regolin, L., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 9–24. [Google Scholar] [CrossRef]
- Pfannkuche, K.A.; Bouma, A.; Groothuis, T.G. Does testosterone affect lateralization of brain and behaviour? A meta-analysis in humans and other animal species. Philos. Trans. R. Soc. B Boil. Sci. 2009, 364, 929–942. [Google Scholar] [CrossRef] [PubMed]
- Schaafsma, S.M.; Groothuis, T.G. Sex-specific effects of postnatal testosterone on lateralization in cichlid fish. Anim. Behav. 2011, 81, 283–288. [Google Scholar] [CrossRef]
- Rogers, L.J. Light input and the reversal of functional lateralization in the chicken brain. Behav. Brain Res. 1990, 38, 211–221. [Google Scholar] [CrossRef]
- Andrew, R.J.; Osorio, D.; Budaev, S. Light during embryonic development modulates patterns of lateralization strongly and similarly in both zebrafish and chick. Philos. Trans. R. Soc. B Boil. Sci. 2009, 364, 983–989. [Google Scholar] [CrossRef]
- Casey, M.B.; Sleigh, M.J. Prenatal visual experience induces postnatal motor laterality in japanese quail chicks (Coturnix coturnix japonica). Dev. Psychobiol. 2013, 56, 489–497. [Google Scholar] [CrossRef]
- Manns, M.; Ströckens, F. Functional and structural comparison of visual lateralization in birds–similar but still different. Front. Psychol. 2014, 5, 206. [Google Scholar] [CrossRef]
- Güntürkün, O.; Ocklenburg, S. Ontogenesis of lateralization. Neuron 2017, 94, 249–263. [Google Scholar] [CrossRef]
- Budaev, S.; Andrew, R.J. Patterns of early embryonic light exposure determine behavioural asymmetries in zebrafish: A habenular hypothesis. Behav. Brain Res. 2009, 200, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Dadda, M.; Bisazza, A. Prenatal light exposure affects development of behavioural lateralization in a livebearing fish. Behav. Process. 2012, 91, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Vallortigara, G.; Rogers, L.J. Survival with an asymmetrical brain: Advantages and disadvantages of cerebral lateralization. Behav. Brain Sci. 2005, 28, 575–589. [Google Scholar] [CrossRef] [PubMed]
- Koboroff, A.; Kaplan, G.; Rogers, L.J. Hemispheric specialization in Australian magpies (Gymnorhina tibicen) shown as eye preferences during response to a predator. Brain Res. Bull. 2008, 76, 304–306. [Google Scholar] [CrossRef]
- Martín, J.; López, P.; Bonati, B.; Csermely, D. Lateralization when monitoring predators in the wild: A left eye control in the common wall lizard (Podarcis muralis). Ethology 2010, 116, 1226–1233. [Google Scholar] [CrossRef]
- Lippolis, G.; Westman, W.; McAllan, B.; Rogers, L. Lateralisation of escape responses in the stripe-faced dunnart, Sminthopsis macroura (Dasyuridae: Marsupialia). Laterality 2005, 10, 457–470. [Google Scholar] [CrossRef]
- Siniscalchi, M.; Bertino, D.; Quaranta, A. Laterality and performance of agility-trained dogs. Laterality Asymmetries Body Brain Cogn. 2014, 19, 219–234. [Google Scholar] [CrossRef]
- Shibasaki, M.; Nagumo, S.; Koda, H. Japanese monkeys (Macaca fuscata) spontaneously associate alarm calls with snakes appearing in the left visual field. J. Comp. Psychol. 2014, 128, 332. [Google Scholar] [CrossRef]
- Domenici, P.; Allan, B.; McCormick, M.I.; Munday, P.L. Elevated carbon dioxide affects behavioural lateralization in a coral reef fish. Boil. Lett. 2012, 8, 78–81. [Google Scholar] [CrossRef]
- Brown, C.; Western, J.; Braithwaite, V.A. The influence of early experience on, and inheritance of, cerebral lateralization. Anim. Behav. 2007, 74, 231–238. [Google Scholar] [CrossRef]
- Brown, C.; Braithwaite, V.A. Effects of predation pressure on the cognitive ability of the poeciliid Brachyraphis episcopi. Behav. Ecol. 2004, 16, 482–487. [Google Scholar] [CrossRef]
- Dale Broder, E.; Angeloni, L.M. Predator-induced phenotypic plasticity of laterality. Anim. Behav. 2014, 98, 125–130. [Google Scholar] [CrossRef]
- Jozet-Alves, C.; Hébert, M. Embryonic exposure to predator odour modulates visual lateralization in cuttlefish. Proc. R. Soc. B Biol. Sci. 2013, 280, 1752. [Google Scholar] [CrossRef]
- Ferrari, M.C.; McCormick, M.I.; Meekan, M.G.; Chivers, D.P. Background level of risk and the survival of predator-naive prey: Can neophobia compensate for predator naivety in juvenile coral reef fishes? Proc. R. Soc. B Biol. Sci. 2015, 282, 20142197. [Google Scholar] [CrossRef]
- Lucon-Xiccato, T.; Chivers, D.P.; Mitchell, M.D.; Ferrari, M.C. Prenatal exposure to predation affects predator recognition learning via lateralization plasticity. Behav. Ecol. 2016, 28, arw155. [Google Scholar] [CrossRef]
- Bisazza, A.; Pignatti, R.; Vallortigara, G. Detour tests reveal task- and stimulus-specific behavioural lateralization in mosquitofish (Gambusia holbrooki). Behav. Brain Res. 1997, 89, 237–242. [Google Scholar] [CrossRef]
- Bisazza, A.; Cantalupo, C.; Capocchiano, M.; Vallortigara, G. Population lateralisation and social behaviour: A study with 16 species of fish. Laterality 2000, 5, 269–284. [Google Scholar] [CrossRef]
- Sovrano, V.A.; Bisazza, A.; Vallortigara, G. Lateralization of response to social stimuli in fishes: A comparison between different methods and species. Physiol. Behav. 2001, 74, 237–244. [Google Scholar] [CrossRef]
- Chiandetti, C. Manipulation of strength of cerebral lateralization via embryonic light stimulation in birds. In Lateralized Brain Functions; Springer: Berlin, Germany, 2017; pp. 611–631. [Google Scholar]
- Monteforte, S.; Cattelan, S.; Morosinotto, C.; Pilastro, A.; Grapputo, A. Maternal predator-exposure affects offspring size at birth but not telomere length in a live-bearing fish. Ecol. Evol. 2020, 10, 2030–2039. [Google Scholar] [CrossRef]
- Rogers, L.J.; Deng, C. Corticosterone treatment of the chick embryo affects light-stimulated development of the thalamofugal visual pathway. Behav. Brain Res. 2005, 159, 63–71. [Google Scholar] [CrossRef]
- Rougeot, C.; Krim, A.; Mandiki, S.N.M.; Kestemont, P.; Melard, C. Sex steroid dynamics during embryogenesis and sexual differentiation in Eurasian perch, Perca fluviatilis. Theriogenology 2007, 67, 1046–1052. [Google Scholar] [CrossRef] [PubMed]
- Bisazza, A.; Brown, C. Lateralization of Cognitive Functions in Fish. In Fish Cognition and Behavior; Brown, C., Laland, K., Krause, J., Eds.; Wiley-Blackwell, Wiley: Chichester, UK; Ames, IA, USA, 2011; pp. 298–324. [Google Scholar] [CrossRef]
- McCormick, M.I. Behaviorally induced maternal stress in a fish influences progeny quality by a hormonal mechanism. Ecology 1998, 79, 1873–1883. [Google Scholar] [CrossRef]
- Schaafsma, S.M.; Groothuis, T.G. Sex-specific effects of maternal testosterone on lateralization in a cichlid fish. Anim. Behav. 2012, 83, 437–443. [Google Scholar] [CrossRef]
- Barcellos, L.J.G.; Volpato, G.L.; Barreto, R.E.; Coldebella, I.; Ferreira, D. Chemical communication of handling stress in fish. Physiol. Behav. 2011, 103, 372–375. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Masuda, R. Net-chasing training improves the behavioral characteristics of hatchery-reared red sea bream (Pagrus major) juveniles. Can. J. Fish. Aquat. Sci. 2018, 75, 861–867. [Google Scholar] [CrossRef]
- Smith, B.R.; Blumstein, D.T. Structural consistency of behavioural syndromes: Does predator training lead to multi-contextual behavioural change? Behaviour 2012, 149, 187–213. [Google Scholar] [CrossRef]
- Moscicki, M.K.; Hurd, P.L. Sex, boldness and stress experience affect convict cichlid, Amatitlania nigrofasciata, open field behaviour. Anim. Behav. 2015, 107, 105–114. [Google Scholar] [CrossRef]
- Brown, C.; Jones, F.; Braithwaite, V. In situ examination of boldness–shyness traits in the tropical poeciliid, Brachyraphis episcopi. Anim. Behav. 2005, 70, 1003–1009. [Google Scholar] [CrossRef]
- Brown, C.; Gardner, C.; Braithwaite, V.A. Differential stress responses in fish from areas of high-and low-predation pressure. J. Comp. Physiol. B 2005, 175, 305–312. [Google Scholar] [CrossRef]
- Peñalva-Arana, D.C.; Lynch, M.; Robertson, H.M. The chemoreceptor genes of the waterflea Daphnia pulex: Many Grs but no Ors. BMC Evol. Boil. 2009, 9, 79. [Google Scholar] [CrossRef]
- Chivers, D.P.; Zhao, X.; Brown, G.E.; Marchant, T.A.; Ferrari, M.C. Predator-induced changes in morphology of a prey fish: The effects of food level and temporal frequency of predation risk. Evol. Ecol. 2008, 22, 561–574. [Google Scholar] [CrossRef]
- Alcalay, Y.; Puzhevsky, D.; Tsurim, I.; Scharf, I.; Ovadia, O. Interactive and sex-specific life-history responses of Culex pipiens mosquito larvae to multiple environmental factors. J. Zool. 2018, 306, 268–278. [Google Scholar] [CrossRef]
- Ferrari, M.C.; McCormick, M.I.; Mitchell, M.D.; Allan, B.J.; Gonçalves, E.J.; Chivers, D.P. Daily variation in behavioural lateralization is linked to predation stress in a coral reef fish. Anim. Behav. 2017, 133, 189–193. [Google Scholar] [CrossRef]
- Siniscalchi, M.; Bertino, D.; d’Ingeo, S.; Quaranta, A. Relationship between Motor Laterality and Aggressive Behavior in Sheepdogs. Symmetry 2019, 11, 233. [Google Scholar] [CrossRef]
- Siniscalchi, M.; d’Ingeo, S.; Quaranta, A. Lateralized functions in the dog brain. Symmetry 2017, 9, 71. [Google Scholar] [CrossRef]
- Bell, A.M.; Sih, A. Exposure to predation generates personality in threespined sticklebacks (Gasterosteus aculeatus). Ecol. Lett. 2007, 10, 828–834. [Google Scholar] [CrossRef]
- Dingemanse, N.J.; Wright, J.; Kazem, A.J.; Thomas, D.K.; Hickling, R.; Dawnay, N. Behavioural syndromes differ predictably between 12 populations of three-spined stickleback. J. Anim. Ecol. 2007, 76, 1128–1138. [Google Scholar] [CrossRef]
- Deng, C.; Rogers, L.J. Factors affecting the development of lateralization in chicks. In Comparative Vertebrate Lateralization; Cambridge University Press: Cambridge, UK, 2002; pp. 206–246. [Google Scholar]
- Halpern, M.E.; Güntürkün, O.; Hopkins, W.D.; Rogers, L.J. Lateralization of the vertebrate brain: Taking the side of model systems. J. Neurosci. 2005, 25, 10351–10357. [Google Scholar] [CrossRef]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dadda, M.; Vendramin, V.; Agrillo, C. Prenatal Visual Exposure to a Predator Influences Lateralization in Goldbelly Topminnows. Symmetry 2020, 12, 1257. https://doi.org/10.3390/sym12081257
Dadda M, Vendramin V, Agrillo C. Prenatal Visual Exposure to a Predator Influences Lateralization in Goldbelly Topminnows. Symmetry. 2020; 12(8):1257. https://doi.org/10.3390/sym12081257
Chicago/Turabian StyleDadda, Marco, Veronica Vendramin, and Christian Agrillo. 2020. "Prenatal Visual Exposure to a Predator Influences Lateralization in Goldbelly Topminnows" Symmetry 12, no. 8: 1257. https://doi.org/10.3390/sym12081257
APA StyleDadda, M., Vendramin, V., & Agrillo, C. (2020). Prenatal Visual Exposure to a Predator Influences Lateralization in Goldbelly Topminnows. Symmetry, 12(8), 1257. https://doi.org/10.3390/sym12081257