
Analyses of Morphological Differences between Geographically Distinct Populations of Gymnodiptychus dybowskii

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
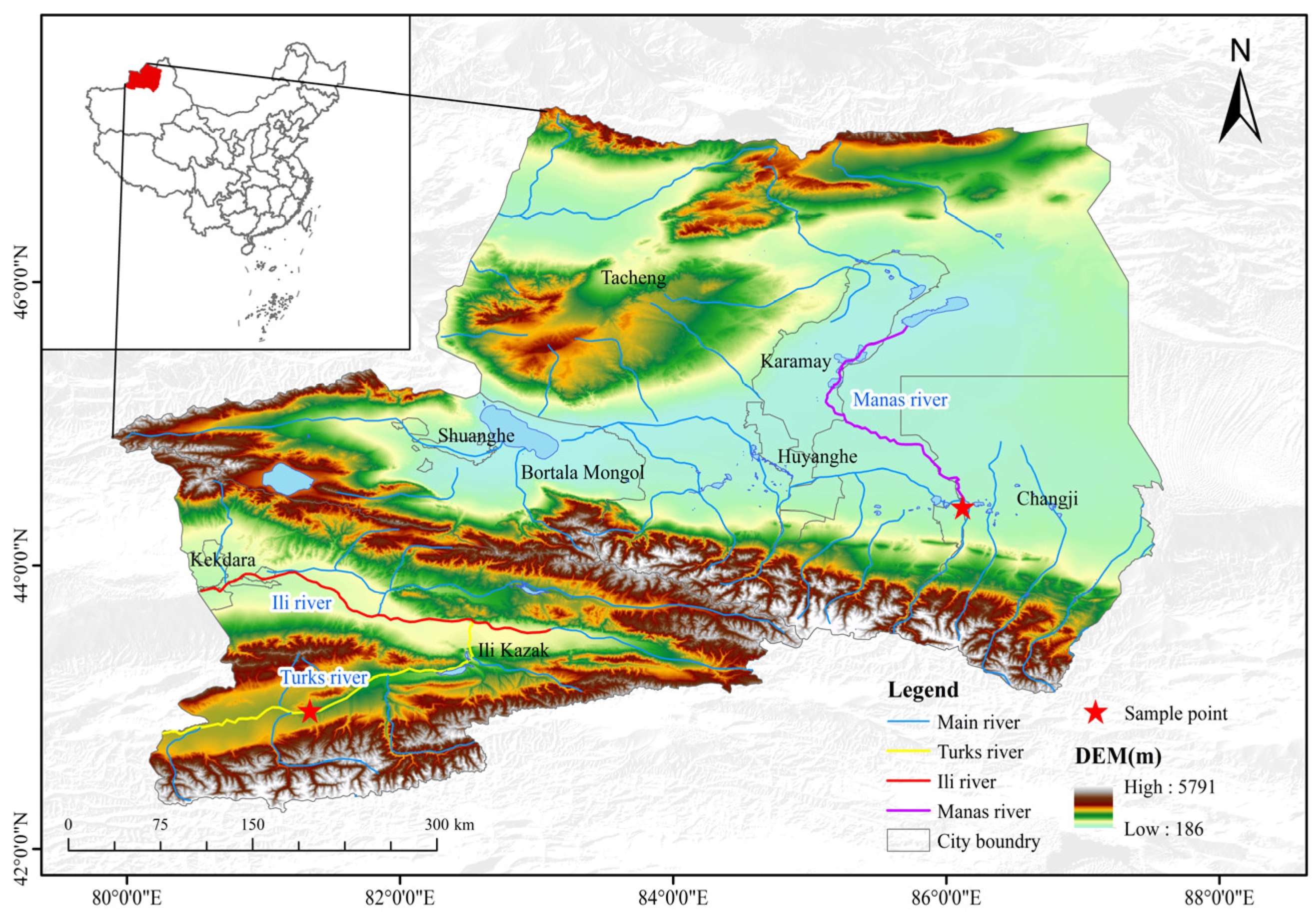
2.1. Materials
2.2. Methodology
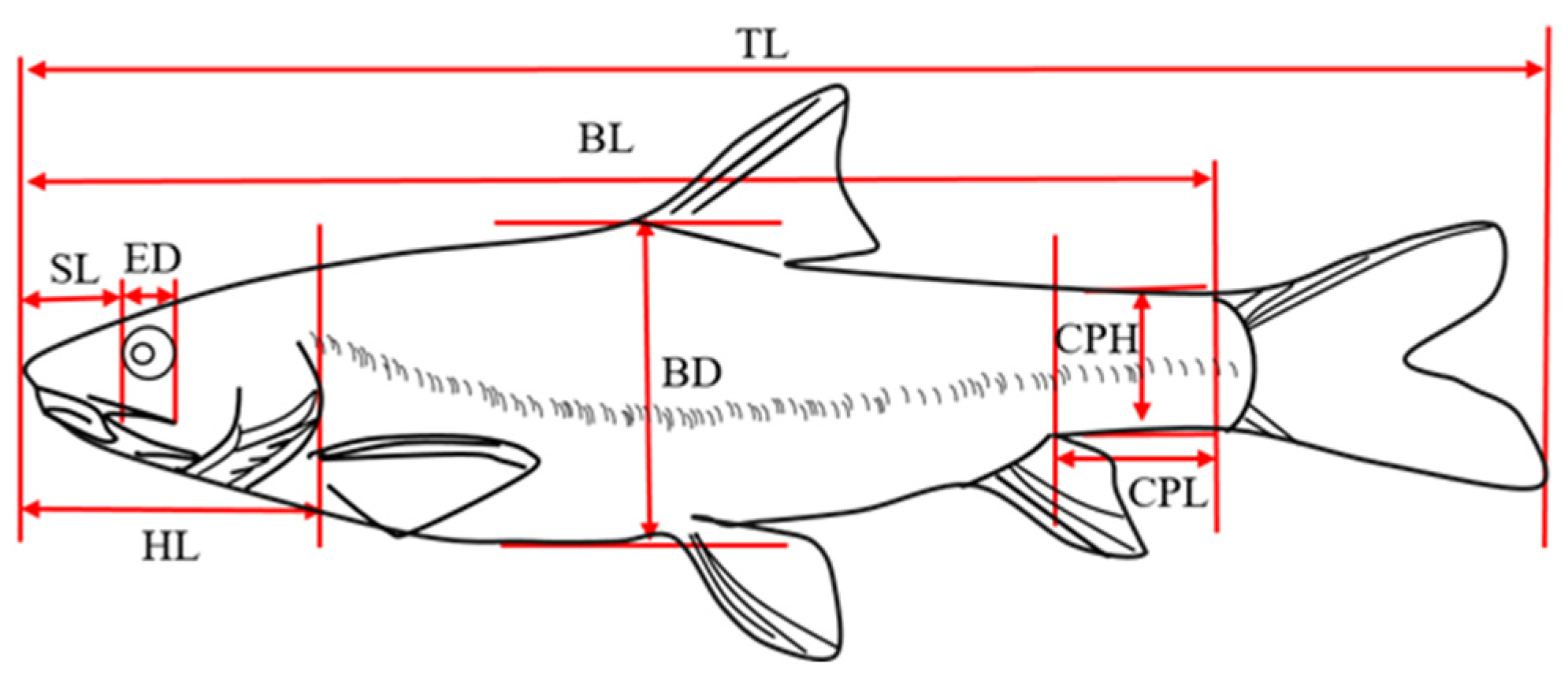
2.2.1. Morphological Indicators
2.2.2. Morphological Analysis
2.3. Data Processing
3. Results
3.1. Characteristics of G. dybowskii
3.2. One-Way ANOVA
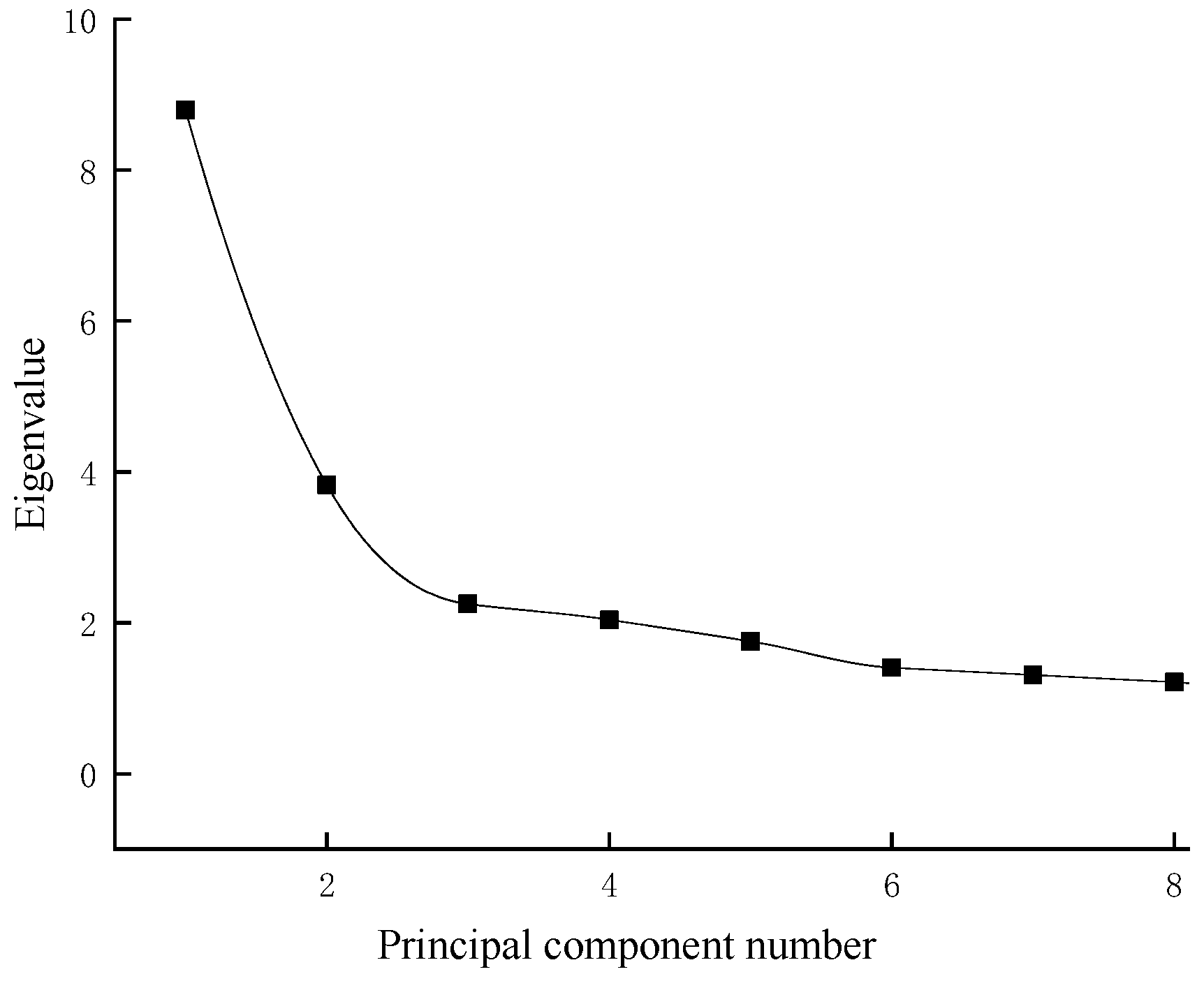
3.3. Principal Component Analysis
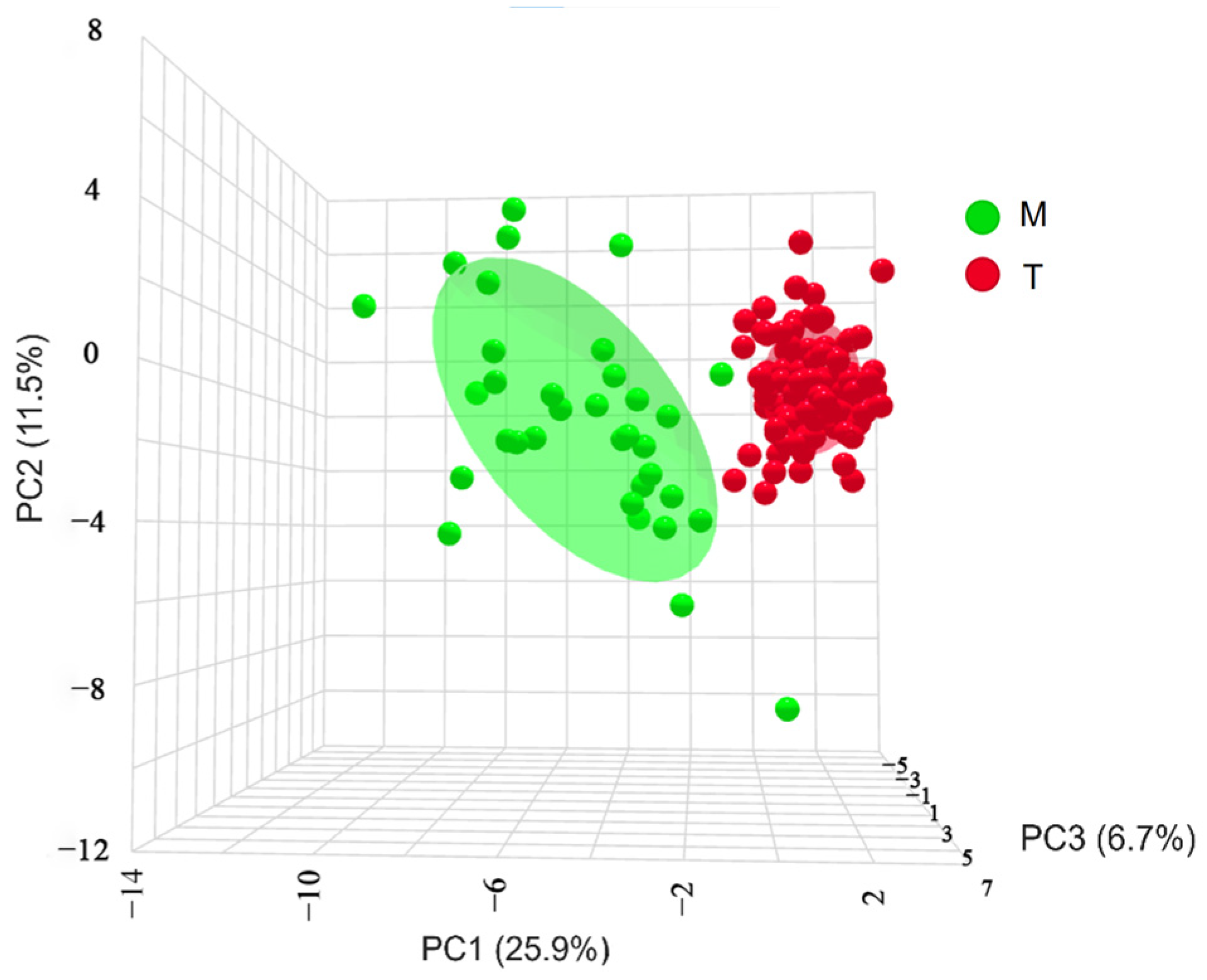
3.4. Orthogonal Partial Least Squares Discriminant Analysis (OPLS-DA)
3.5. Discriminant Analysis
3.6. Analysis of the Coefficient of Difference
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | T | M | p |
|---|---|---|---|
| Mean ± SD | Mean ± SD | ||
| TL/BL | 1.235 ± 0.316 | 1.163 ± 0.049 | 0.46 |
| BD/BL | 0.177 ± 0.012 | 0.180 ± 0.029 | 0.217 |
| CPH/BL | 0.068 ± 0.005 | 0.063 ± 0.009 | 0.128 |
| I—J/BL | 0.284 ± 0.015 | 0.308 ± 0.024 | 0.495 |
| A—J/BL | 0.260 ± 0.013 | 0.237 ± 0.015 | 0.615 |
| AI/BL | 0.537 ± 0.012 | 0.533 ± 0.049 | 0.09 |
| B—J/BL | 0.141 ± 0.008 | 0.140 ± 0.011 | 0.695 |
| B—I/BL | 0.374 ± 0.013 | 0.383 ± 0.053 | 0.091 |
| C—J/BL | 0.244 ± 0.015 | 0.252 ± 0.047 | 0.984 |
| H—G/BL | 0.088 ± 0.009 | 0.130 ± 0.063 | 0.181 |
References
- Guo, Y.; Zhang, R.M.; Cai, L.G. Xinjiang of Fishery; Xinjiang Science and Technology Press: Urumqi, China, 2012; ISBN 9787546615295. [Google Scholar]
- Cao, W.X.; Chen, Y.Y.; Wu, Y.F. Origin and evolution of schizothoracine fishes in relation to the upheaval of the Xizang Plateau. In Tibetan Expedition Team of the Chinese Academy of Science ed. Studies on the Period, Amplitude and Type of the Uplift of the Qinghai-Xizang Plateau; Science Press: Beijing, China, 1981; pp. 118–130. [Google Scholar]
- He, D.K.; Chen, Y.F.; Chen, Y.Y.; Chen, Z.M. Molecular phylogeny of the specialized Schizothoracine fishes (Teleostei: Cyprinidae), with their mplications for the uplift of the Qinghai-Tibetan Plateau. J. Chin. Sci. Bull. 2004, 49, 39–48. [Google Scholar] [CrossRef]
- Qi, D.; Chao, Y.; Guo, S.; Zhao, L.; Li, T.; Wei, F.; Zhao, X. Convergent, parallel and correlated evolution of trophic morphologies in the subfamily Schizothoracinae from the Qinghai-Tibetan Plateau. PLoS ONE 2012, 7, e34070. [Google Scholar] [CrossRef]
- Guan, L.; Chi, W.; Xiao, W.; Chen, L.; He, S. Analysis of ypoxia-inducible factor alpha polyploidization reveals adaptation to Tibetan plateau in the evolution of Schizothoracine fish. BMC Evol. Biol. 2014, 14, 192. [Google Scholar] [CrossRef]
- Zhu, X.F.; Chen, Y.F. Preliminary study on the age and growth characteristics of Schizothorax macropogon. Chin. J. Zool. 2009, 44, 76–82. [Google Scholar]
- Wang, Z.; Yan, Y.C.; Jiang, P.G.; Yan, A. Effects of different factors on soil moisture in farmland in Manas River Basin. Xinjiang Agric. Sci. 2013, 50, 1879–1886. [Google Scholar]
- Li, Y.Y.; Zhang, H.L.; Zhang, F.H.; Chen, F.; Lai, X.Q. Analysis on the Potential of the Available Agricultur e Water Resour ces in Manas River Valley, Xinjiang. J. Nat. Resour. 2007, 22, 44–50. [Google Scholar] [CrossRef]
- Sui, F.G. Tectonic evolution and its relationship with hydrocarbon accumulation in the northwest margin of Junggar Basin. J. Geol. 2015, 89, 779–793. [Google Scholar] [CrossRef]
- Ren, M.L.; Guo, Y.; Zhang, Q.L.; Zhang, R.M.; Li, H.; A, D.K.; Cai, L.G.; Yong, W.D.; Ren, B.; Gao, H.; et al. Fish Resources and Fishery of Ili River; Heilongjiang Science and Technology Press: Harbin, China, 1998; pp. 13–310. ISBN 7-5388-3369-2. [Google Scholar]
- Chen, L.P. Study on the Ecological Environment of Manas River Basin. Master’s Thesis, Shihezi University, Xinjiang, China, 2020. [Google Scholar] [CrossRef]
- Robinson, W.B.; Parsons, J.K. Changing times, spaces, and faces: Tests and implications of adaptive morphological plasticity in the fishes of northern postglacial lakes. Can. J. Fish. Aquat. Sci. 2002, 59, 1819–1833. [Google Scholar] [CrossRef]
- Langerhans, R.B.; Layman, C.A.; Shokrollahi, A.M.; DeWitt, T. Predator-driven phenotypic diversification in Gambusia affinis. Evolution 2004, 58, 2305–2318. [Google Scholar]
- Lostrom, S.; Evans, J.P.; Grierson, P.F.; Collin, S.P.; Davies, P.M.; Kelley, J.L. Linking stream ecology with morphlogical variabilityin a native freshwater fish from semi-arid Australia. Ecol. Evol. 2015, 5, 3272–3287. [Google Scholar] [CrossRef]
- Scharnweber, K. Morphological and trophic divergence of lake and stream minnows (Phoxinus phoxinus). Ecol. Evol. 2020, 10, 8358–8367. [Google Scholar] [CrossRef]
- Franssen, N.R. Anthropogenic habitat alteration induces rapid morphological divergence in a native stream fish. Evol. Appl. 2011, 4, 791–804. [Google Scholar] [CrossRef]
- Lang, J.J.V.; Snyder, R.J.; Clapsadl, M.D.; Michalak, P.; Kang, L.; Pérez-Fuentetaja, A. Morphometric differentiation and gene flow in emerald shiner (Notropis atherinoides) from the lower Great Lakes and the Niagara River. J. Great Lakes Res. 2018, 45, 324–332. [Google Scholar] [CrossRef]
- Ramler, D.; Palandacic, A.; Delmastro, G.B.; Wanzenbock, J.; Ahnelt, H. Morphological divergence of lake and stream Phoxinus of northern Italy and the Denube basin based on geometric morphometric analysis. Ecol. Evol. 2016, 7, 572–584. [Google Scholar] [CrossRef]
- Li, S.F.; Li, C.H.; Li, J.L. Analysis of morphological differences among strains of Nile tilapia. J. Zool. 1998, 44, 75–82. [Google Scholar]
- Yin, M.C. Fish Ecology; China Agricultural Press: Beijing, China, 1995; ISBN 9787109031432. [Google Scholar]
- Wang, W.; Chen, L.Q.; Gu, Z.M.; Peng, S.M.; Li, Y.K. Analysis of Morphological Variations among Seven Populations of Erythroculter ilishaeformis. Freshw. Fish. 2007, 37, 40–44. [Google Scholar]
- Xie, C.X.; Zhang, J.B.; Diao, X.M.; Kong, X.Y.; Li, S.A.; Wu, H.H.; Zhang, J.P.; Wu, Y.F.; Qin, W.; Huang, Q.; et al. Ichthyology; China Agricultural Press: Beijing, China, 2010; ISBN 978-7-109-13579-6. [Google Scholar]
- Wang, J.L.; Chen, Q.H.; Lu, W.Z.; Yuan, S.Q.; Hu, Q.X.; Cen, S.S.; Zhou, C.J.; Meng, X.L.; Nie, G.X.; Gu, Q.H. Morphological differences among five species in rhodeinae in Huaihe River basin in Henan Province. J. Aquat. Biol. 2019, 43, 123–132. [Google Scholar]
- Dray, S.; Dufour, A. The ade4 Package: Implementing the Duality Diagram for Ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef]
- Trygg, J.; Wold, S. Orthogonal projections to latent structures (O-PLS). J. Chemom. 2002, 16, 119–128. [Google Scholar] [CrossRef]
- Mayr, E.; Gorton, L.E.; Usinger, R.L. Methods and Principles of Systematic Zoology; Mc-Graw-Hill Book Company: New York, NY, USA, 1953; pp. 125–154. [Google Scholar]
- Zhang, Y.J.; Chen, S.A.; Wang, C.X.; Wang, X.Y.; Li, J.L.; Xie, C.X.; Wei, Q. Morphological differences among different geographic populations of the naked heavy-lipped fishes in Xinjiang. Xinjiang Agric. Sci. 2023, 60, 511–520. [Google Scholar] [CrossRef]
- Pashkov, N.A. Morphological characteristics of the populations of black-striped pipefish Syngnathus abaster Risso, 1827 (Pisces, Actinopterygii, Syngnathidae) in some North Caucasian water bodies. Mar. Biol. J. 2017, 2, 55–69. [Google Scholar]
- Vincent, R.; Marc, P.; Pierre, M. Parallel evolution of morphological traits and body shape in littoral and pelagic brook charr, Salvelinus fontinalis, along a gradient of interspecific competition. Oecologia 2021, 197, 421–436. [Google Scholar]
- Hetzel, C.; Forsythe, P. Phenotypic plasticity of a generalist fish species resident to lotic environments: Insights from the Great Lakes region. Ecol. Evol. 2023, 13, e10715. [Google Scholar] [CrossRef]
- Caiger, P.E.; Croq, C.; Clements, K.D. Environmentally induced morphological variation in the temperate reef fish, Forsterygion lapillum (F. Tripterygiidae). Mar. Biol. 2021, 168, 131. [Google Scholar] [CrossRef]
- Dunn, N.R.; O’Brien, L.K.; Burridge, C.P.; Closs, G.P. Morphological Convergence and Divergence in Galaxias Fishes in Lentic and Lotic Habitats. Diversity 2020, 12, 183. [Google Scholar] [CrossRef]
- Langerhans, R.B.; Layman, C.A.; Langerhans, A.K.; Dewitt, T.J. Habitat-associated morphological divergence in two Neotropical fish species. Biol. J. Linn. Soc. 2003, 80, 689–698. [Google Scholar]
- Ola-Oladimeji, F.A.; Oso, J.A.; Oladimeji, T.E.; Idowu, E.O.; Adeleke, K.; Urihe, F.O. Phenotypic Diversities of Four Populations of Clarias Gariepinus (Siluriformes, Clariidae) Obtained from Ogun and Ondo State Waterbodies in South-Western Nigeria. Vestn. Zool. 2017, 51, 285–294. [Google Scholar]
- Iqbal, M.Z.; Chamily, F.A.; Rahman, M.M.; Tasnim, R.; Mohiuddin, M.; Sultana, F.; Rahman, S.M.; Ali, M.M. Habitat salinity and source-induced variation in body shape of euryhaline long whiskers catfish (Mystus gulio). Reg. Stud. Mar. Sci. 2024, 69, 103308. [Google Scholar]
- Wang, Y.; Ding, J.; Li, X.; Zhang, J.; Ma, G. Impact of LUCC on ecosystem services values in the Yili River Basin based on an intensity analysis model. J. Ecol. 2022, 42, 3106–3118. [Google Scholar]
- Jiang, L. Analysis on Groundwater Characteristics at Mosuowan Irrigation District of the ManasRiver Basin. Groundwater 2016, 38, 1–3. [Google Scholar]
- Jiang, E.; Yang, Y.; Pan, M.; Tao, Y. Efficacy evaluation and metabolomics analysis of raw and salt-processed Achyranthes bidentata Radix in zebrafish larvae for osteoporosis treatment. J. Pharm. Biomed. Anal. 2024, 237, 115774. [Google Scholar] [CrossRef]
- Mong, Y.X.; Wang, G.H.; Xiong, D.M.; Liu, H.X.; Zhang, J.L.; Wang, J.L.; Wang, L.X.; Liu, X.L. Exploring the validity of subspecies of Qinling Chinook salmon based on morphological differences. J. Aquat. Biol. 2018, 42, 550–560. [Google Scholar]



| Traits | T | M | p |
|---|---|---|---|
| Mean ± SD | Mean ± SD | ||
| BW/BL | 0.139 ± 0.012 | 0.135 ± 0.019 | <0.001 ** |
| HL/BL | 0.253 ± 0.011 | 0.228 ± 0.022 | 0.007 ** |
| SL/BL | 0.075 ± 0.007 | 0.080 ± 0.012 | <0.001 ** |
| ED/BL | 0.055 ± 0.006 | 0.036 ± 0.010 | <0.001 ** |
| EI/BL | 0.074 ± 0.006 | 0.081 ± 0.007 | <0.001 ** |
| CPL/BL | 0.127 ± 0.016 | 0.173 ± 0.046 | <0.001 ** |
| A—B/BL | 0.201 ± 0.010 | 0.187 ± 0.017 | <0.001 ** |
| B—C/BL | 0.267 ± 0.014 | 0.274 ± 0.046 | <0.001 ** |
| C—D/BL | 0.127 ± 0.010 | 0.110 ± 0.016 | <0.001 ** |
| D—E/BL | 0.331 ± 0.029 | 0.403 ± 0.085 | <0.001 ** |
| E—F/BL | 0.071 ± 0.006 | 0.114 ± 0.060 | 0.03 * |
| F—H/BL | 0.205 ± 0.012 | 0.252 ± 0.037 | <0.001 ** |
| H—I/BL | 0.195 ± 0.022 | 0.205 ± 0.029 | <0.001 ** |
| A—C/BL | 0.455 ± 0.025 | 0.457 ± 0.030 | <0.001 ** |
| C—I/BL | 0.180 ± 0.014 | 0.194 ± 0.022 | <0.001 ** |
| C—H/BL | 0.319 ± 0.026 | 0.319 ± 0.045 | <0.001 ** |
| C—F/BL | 0.482 ± 0.036 | 0.551 ± 0.031 | <0.001 ** |
| D—I/BL | 0.144 ± 0.013 | 0.156 ± 0.026 | <0.001 ** |
| D—H/BL | 0.202 ± 0.013 | 0.225 ± 0.025 | <0.001 ** |
| D—F/BL | 0.358 ± 0.017 | 0.440 ± 0.026 | <0.001 ** |
| E—H/BL | 0.227 ± 0.013 | 0.290 ± 0.016 | <0.001 ** |
| J—K/BL | 0.037 ± 0.005 | 0.093 ± 0.062 | <0.001 ** |
| I—L/BL | 0.043 ± 0.005 | 0.077 ± 0.050 | 0.004 ** |
| Traits | Principal Components | ||
|---|---|---|---|
| PC1 | PC2 | PC3 | |
| TL/BL | 0.0269 | 0.1811 | 0.2311 |
| BD/BL | −0.0677 | 0.3448 | −0.2049 |
| BW/BL | 0.0066 | 0.2441 | −0.3475 a |
| HL/BL | 0.2301 | 0.2124 | 0.0575 |
| SL/BL | −0.0403 | 0.1062 | −0.1167 |
| ED/BL | 0.2400 | 0.1326 | 0.1405 |
| EI/BL | −0.1731 | 0.1897 | −0.0684 |
| CPL/BL | −0.2145 | −0.0778 | 0.1633 |
| CPH/BL | 0.0750 | 0.3008 a | −0.0511 |
| A—B/BL | 0.1543 | 0.2680 a | 0.2055 |
| B—C/BL | −0.0835 | 0.0090 | 0.1412 |
| C—D/BL | 0.1552 | 0.1579 | −0.1804 |
| D—E/BL | −0.1876 | −0.0088 | −0.0190 |
| E—F/BL | −0.1557 | −0.0788 | −0.1160 |
| F—H/BL | −0.2374 | −0.0484 | −0.0071 |
| H—I/BL | −0.0899 | −0.0619 | −0.0867 |
| I—J/BL | −0.1884 | 0.1049 | −0.2560 a |
| A—J/BL | 0.2092 | 0.2177 | 0.2051 |
| A—I/BL | −0.0276 | 0.2177 | 0.0015 |
| B—J/BL | −0.0251 | 0.1918 | 0.1029 |
| B—I/BL | −0.1060 | 0.2287 | 0.0059 |
| A—C/BL | −0.0422 | 0.1484 | 0.0698 |
| C—J/BL | −0.0631 | −0.0088 | −0.0920 |
| C—I/BL | −0.1599 | 0.2499 | −0.0863 |
| C—H/BL | 0.0029 | 0.1119 | −0.1902 |
| C—F/BL | −0.2216 a | −0.0659 | −0.1287 |
| D—I/BL | −0.1699 | 0.3295 a | −0.0890 |
| D—H/BL | −0.2305 | 0.1111 | −0.0782 |
| D—F/BL | −0.2977 | −0.0611 | −0.0914 |
| E—H/BL | −0.2821 a | −0.0496 | −0.0523 |
| J—K/BL | −0.2679 a | 0.0541 | 0.3061 a |
| I—L/BL | −0.2425 | 0.0797 | 0.3426 a |
| H—G/BL | −0.2288 | 0.0614 | 0.3476 a |
| Eigenvalue | 8.80 | 3.92 | 2.27 |
| Percentage of Variance (%) | 25.9 | 11.5 | 6.7 |
| Cumulative (%) | 25.9 | 37.4 | 44.1 |
| Method | Population | Predicted Group Membership | Identification Accuracy/Percent | Discriminant Accuracy/Percent | |
|---|---|---|---|---|---|
| T | M | ||||
| Stepwise discrimination | T | 123 | 0 | 100 | 100 |
| M | 0 | 35 | 100 | ||
| Traits | T | M | CD | ||
|---|---|---|---|---|---|
| Min~Max | Mean ± SD | Min~Max | Mean ± SD | ||
| TL/BL | 0.1106–2.2928 | 1.2354 ± 0.3158 | 0.9567–1.2606 | 1.1633 ± 0.0494 | 0.1490 |
| BD/BL | 0.1600–0.2300 | 0.1772 ± 0.0124 | 0.1200–0.2500 | 0.1797 ± 0.0292 | −0.4488 |
| BW/BL | 0.1100–0.1700 | 0.1386 ± 0.0116 | 0.0900–0.1700 | 0.1354 ± 0.0187 | −2.2707 |
| HL/BL | 0.2100–0.2700 | 0.2530 ± 0.0109 | 0.1400–0.2600 | 0.2280 ± 0.0219 | 0.8481 |
| SL/BL | 0.0600–0.1000 | 0.0754 ± 0.0069 | 0.0500–0.1000 | 0.0797 ± 0.0120 | −4.7870 |
| ED/BL | 0.0400–0.0700 | 0.0554 ± 0.0060 | 0.0200–0.0600 | 0.0363 ± 0.0100 | 12.6230 |
| EI/BL | 0.0600–0.0900 | 0.0737 ± 0.0063 | 0.0700–0.0900 | 0.0814 ± 0.0069 | 1.5063 * |
| CPL/BL | 0.1100–0.1900 | 0.1269 ± 0.0160 | 0.0700–0.3700 | 0.1726 ± 0.0463 | −1.3499 |
| CPH/BL | 0.0600–0.0800 | 0.0675 ± 0.0049 | 0.0400–0.0800 | 0.0626 ± 0.0085 | −1.8644 |
| A—B/BL | 0.1800–0.2200 | 0.2006 ± 0.0098 | 0.1500–0.2300 | 0.1874 ± 0.0169 | 0.2266 |
| B—C/BL | 0.2200–0.3000 | 0.2670 ± 0.0140 | 0.1800–0.4900 | 0.2743 ± 0.0462 | −2.5602 |
| C—D/BL | 0.1000–0.1600 | 0.1267 ± 0.0097 | 0.0500–0.1400 | 0.1097 ± 0.0164 | 1.3048 * |
| D—E/BL | 0.1000–0.3700 | 0.3305 ± 0.0289 | 0.0500–0.4600 | 0.4034 ± 0.0848 | 0.7900 |
| E—F/BL | 0.0600–0.0900 | 0.0707 ± 0.0055 | 0.0600–0.4500 | 0.1140 ± 0.0604 | 1.9013 * |
| F—H/BL | 0.1800–0.2400 | 0.2051 ± 0.0123 | 0.0900–0.3000 | 0.2517 ± 0.0368 | 1.4903 * |
| H—I/BL | 0.0200–0.2300 | 0.1949 ± 0.0222 | 0.0900–0.2600 | 0.2049 ± 0.0289 | 2.5655 * |
| I—J/BL | 0.2500–0.3200 | 0.2842 ± 0.0151 | 0.2400–0.3600 | 0.3077 ± 0.0243 | −8.3643 |
| A—J/BL | 0.2200–0.2900 | 0.2596 ± 0.0126 | 0.2000–0.2600 | 0.2371 ± 0.0153 | −0.0874 |
| A—I/BL | 0.5000–0.5700 | 0.5367 ± 0.0117 | 0.3000–0.6000 | 0.5334 ± 0.0495 | −0.3953 |
| B—J/BL | 0.1200–0.1600 | 0.1410 ± 0.0080 | 0.1200–0.1700 | 0.1400 ± 0.0106 | 0.2191 |
| B—I/BL | 0.3500–0.4100 | 0.3742 ± 0.0126 | 0.1200–0.4500 | 0.3831 ± 0.0532 | 0.4773 |
| A—C/BL | 0.2300–0.5000 | 0.4550 ± 0.0255 | 0.3800–0.5300 | 0.4571 ± 0.0299 | 0.2541 |
| C—J/BL | 0.1700–0.2800 | 0.2437 ± 0.0146 | 0.0100–0.3100 | 0.2520 ± 0.0473 | 1.8431 * |
| C—I/BL | 0.1500–0.2800 | 0.1799 ± 0.0143 | 0.1500–0.2500 | 0.1940 ± 0.0220 | 0.0105 |
| C—H/BL | 0.2600–0.5100 | 0.3187 ± 0.0265 | 0.0800–0.3600 | 0.3189 ± 0.0454 | −15.8851 |
| C—F/BL | 0.1400–0.5500 | 0.4815 ± 0.0356 | 0.4400–0.6000 | 0.5506 ± 0.0312 | 0.9605 |
| D—I/BL | 0.1100–0.1800 | 0.1439 ± 0.0132 | 0.1200–0.6000 | 0.1563 ± 0.0261 | 1.8586 * |
| D—H/BL | 0.1700–0.2400 | 0.2020 ± 0.0129 | 0.1600–0.2700 | 0.2246 ± 0.0250 | 8.7849 * |
| D—F/BL | 0.3100–0.4100 | 0.3580 ± 0.0165 | 0.3600–0.5100 | 0.4397 ± 0.0258 | 22.7899 * |
| E—H/BL | 0.2000–0.2700 | 0.2274 ± 0.0132 | 0.2400–0.3200 | 0.2903 ± 0.0160 | 0.9998 |
| J—K/BL | 0.0300–0.0500 | 0.0370 ± 0.0054 | 0.0300–0.1700 | 0.0934 ± 0.0618 | 0.7654 |
| I—L/BL | 0.0300–0.0500 | 0.0427 ± 0.0051 | 0.0200–0.1400 | 0.0769 ± 0.0498 | 0.7736 |
| H—G/BL | 0.0500–0.1100 | 0.0880 ± 0.0088 | 0.0600–0.2900 | 0.1300 ± 0.0631 | 0.2706 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, L.; Yao, N.; Wang, C.; Yang, L.; Serekbol, G.; Huo, B.; Qiu, X.; Zi, F.; Song, Y.; Chen, S. Analyses of Morphological Differences between Geographically Distinct Populations of Gymnodiptychus dybowskii. Water 2024, 16, 755. https://doi.org/10.3390/w16050755
Hu L, Yao N, Wang C, Yang L, Serekbol G, Huo B, Qiu X, Zi F, Song Y, Chen S. Analyses of Morphological Differences between Geographically Distinct Populations of Gymnodiptychus dybowskii. Water. 2024; 16(5):755. https://doi.org/10.3390/w16050755
Chicago/Turabian StyleHu, Linghui, Na Yao, Chengxin Wang, Liting Yang, Gulden Serekbol, Bin Huo, Xuelian Qiu, Fangze Zi, Yong Song, and Shengao Chen. 2024. "Analyses of Morphological Differences between Geographically Distinct Populations of Gymnodiptychus dybowskii" Water 16, no. 5: 755. https://doi.org/10.3390/w16050755
APA StyleHu, L., Yao, N., Wang, C., Yang, L., Serekbol, G., Huo, B., Qiu, X., Zi, F., Song, Y., & Chen, S. (2024). Analyses of Morphological Differences between Geographically Distinct Populations of Gymnodiptychus dybowskii. Water, 16(5), 755. https://doi.org/10.3390/w16050755