Species-Richness Responses to Water-Withdrawal Scenarios and Minimum Flow Levels: Evaluating Presumptive Standards in the Tennessee and Cumberland River Basins


Abstract
1. Introduction
2. Materials and Methods
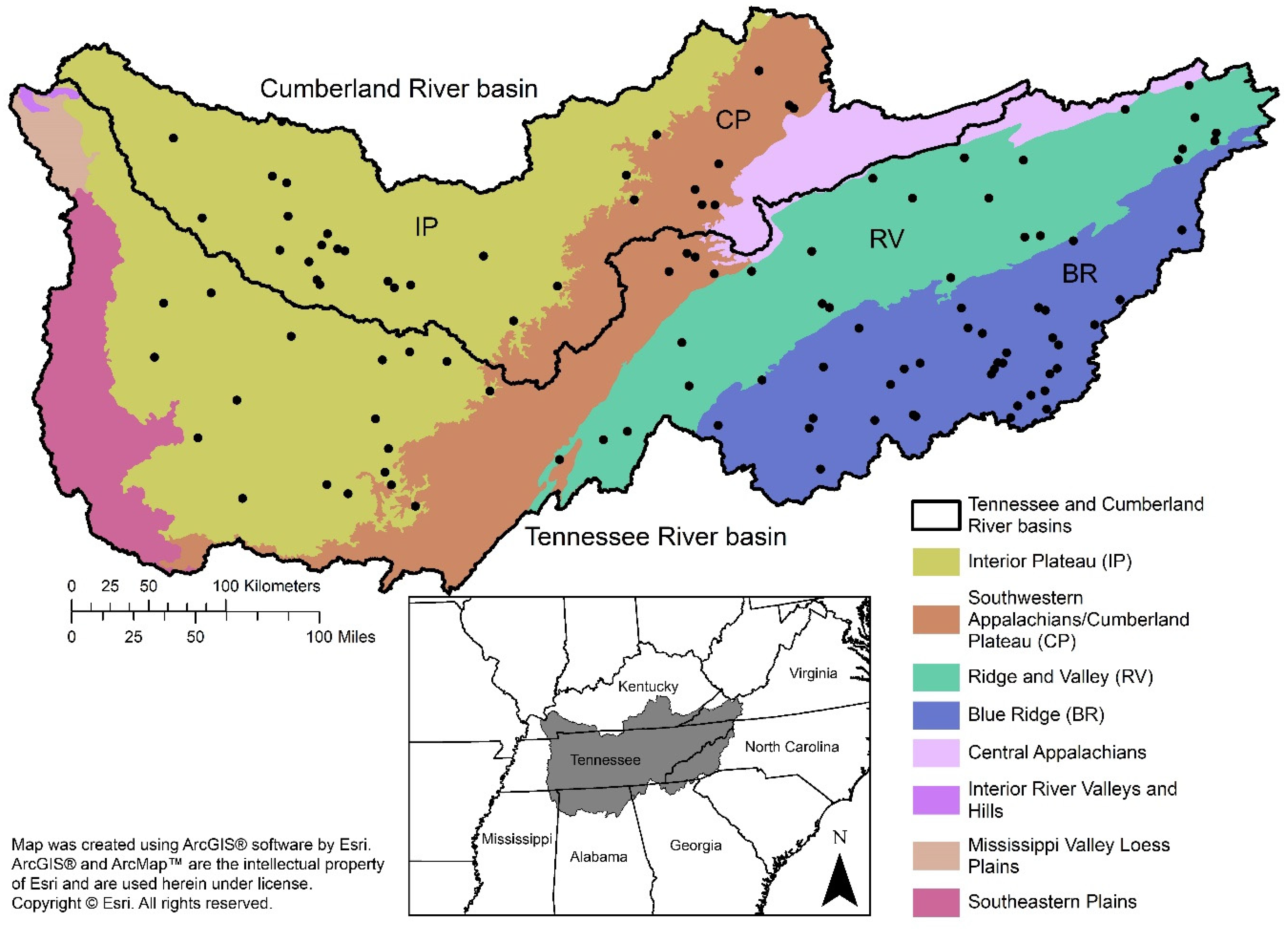
2.1. Study Area
2.2. Site Selection
2.3. Water-Withdrawl Models
2.3.1. Constant-Rate (CR) Withdrawals
2.3.2. Percent-of-Flow (POF) Withdrawals
2.3.3. Minimum Flow Level (MFL) Protection
2.4. Data Processing and Analyses
2.4.1. Calculation of SFCs
2.4.2. SFC Sensitivity
2.4.3. Departure from Reference
2.4.4. Predicted Species Richness Responses to Withdrawal Scenarios
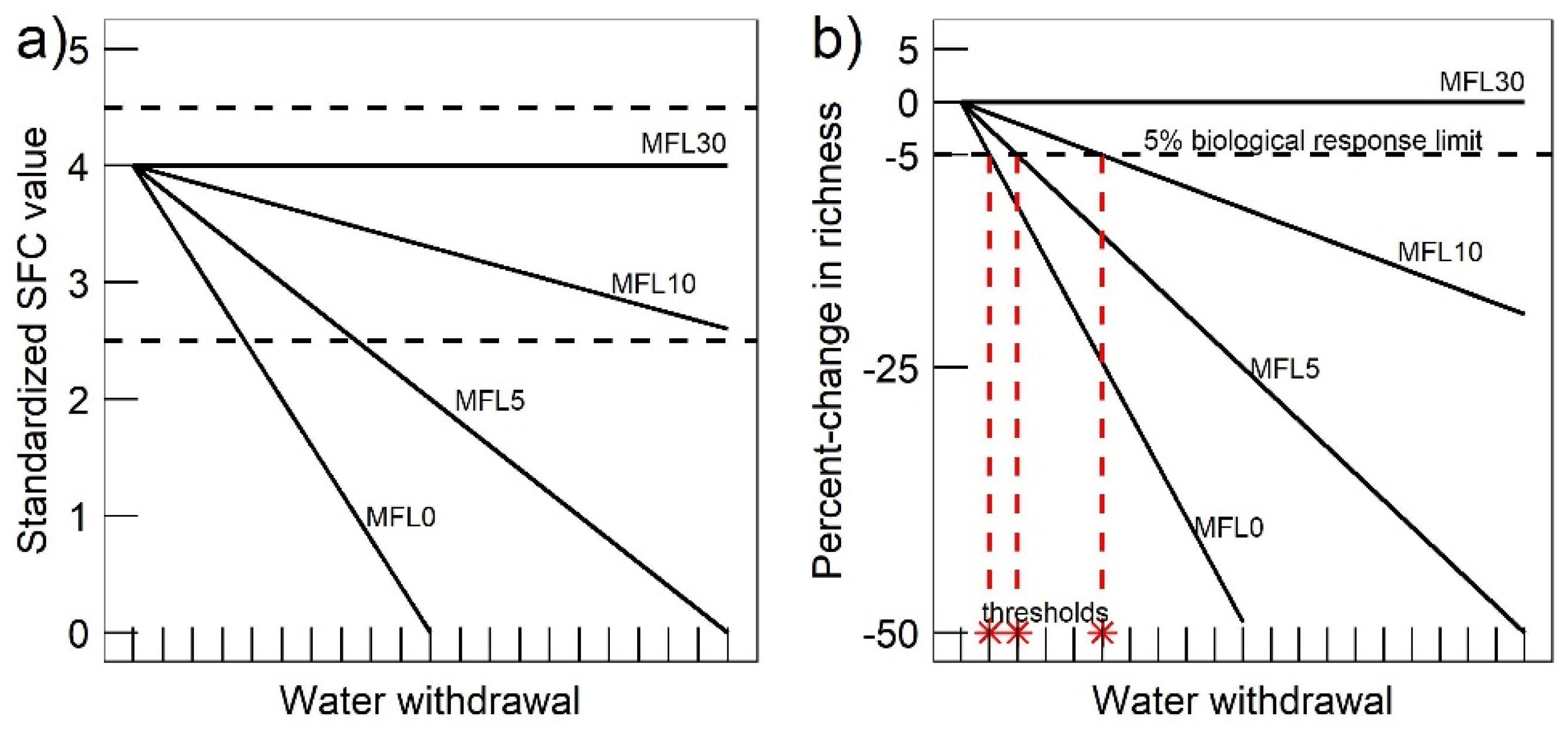
2.4.5. Biological Response Limit and Maximum Withdrawal Thresholds
2.4.6. Model Archive and Bulk Data
3. Results
3.1. Sensitivity of Streamflow Characteristics
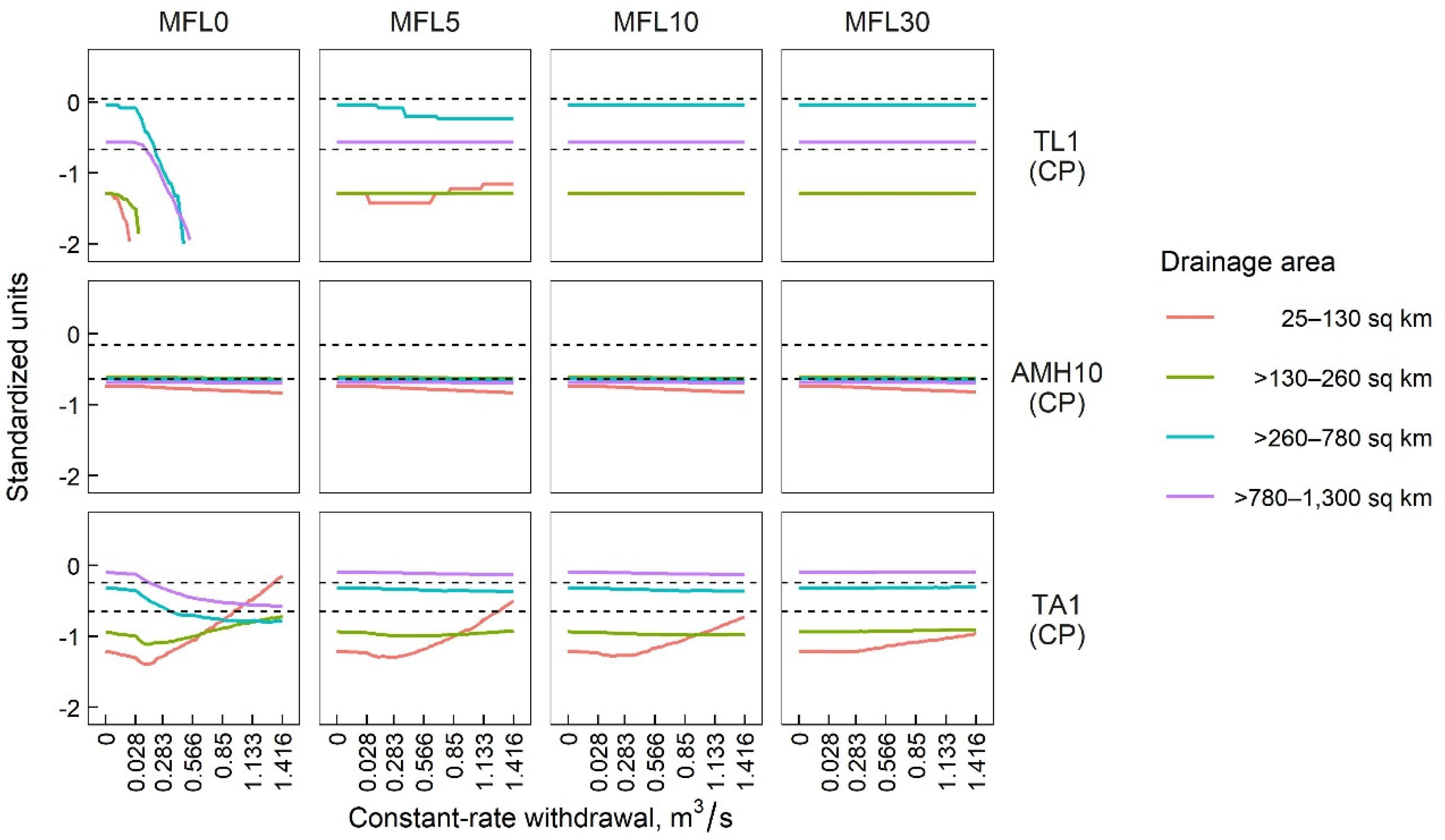
3.1.1. Sensitivity of SFCs to CR Withdrawals
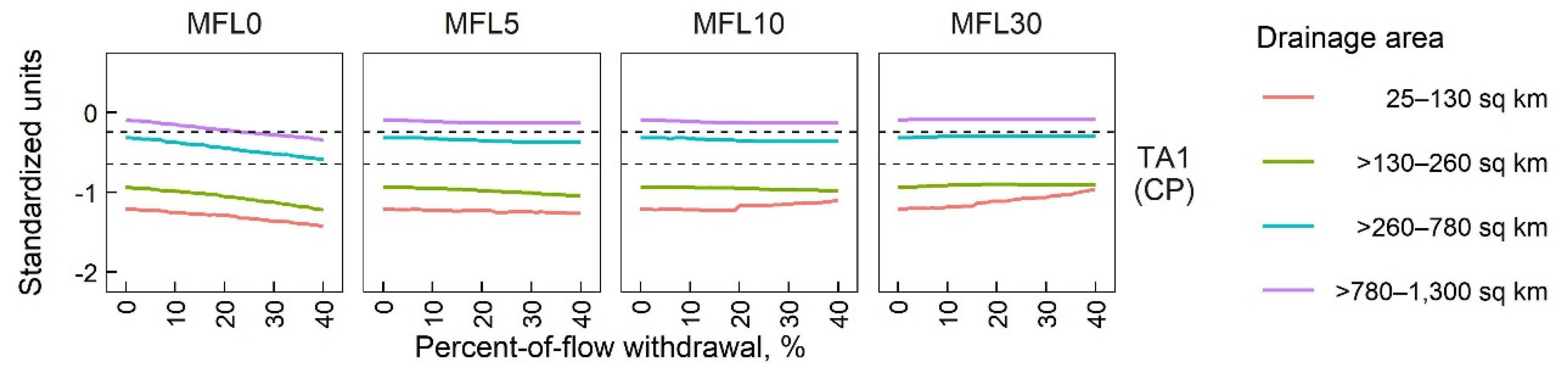
3.1.2. Sensitivity of SFCs to POF Withdrawals
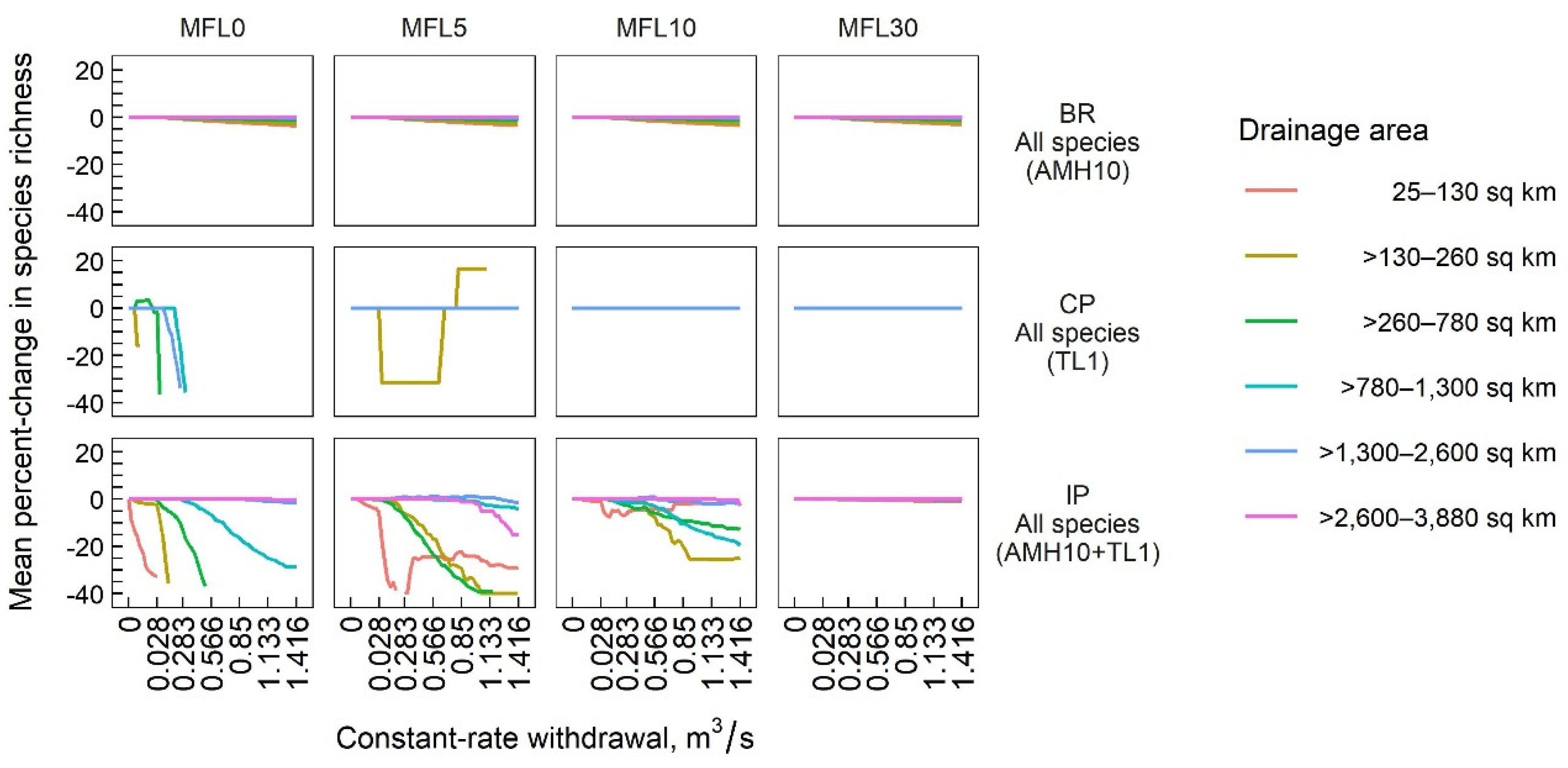
3.2. Species-Richness Responses
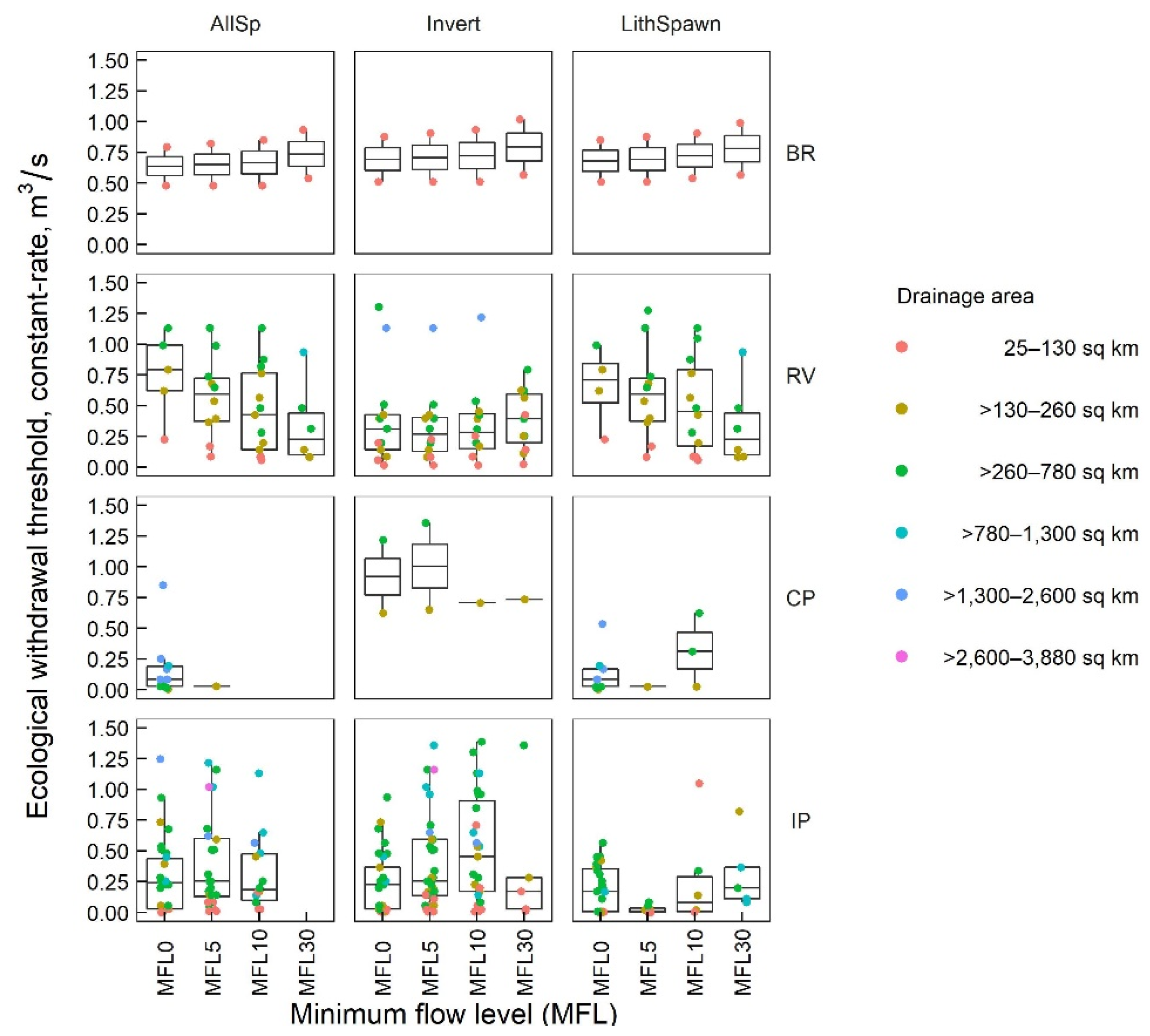
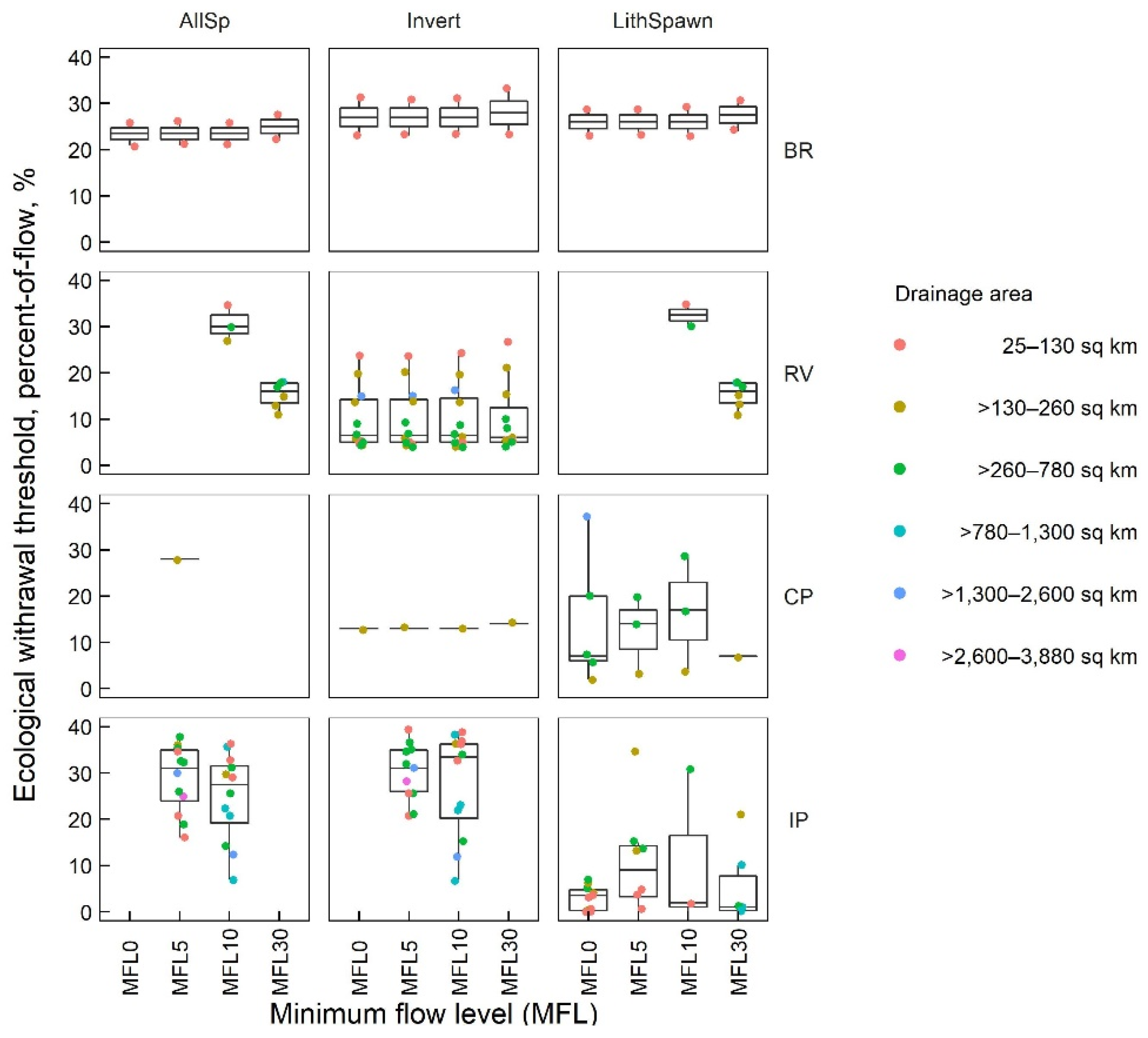
3.3. Ecological Withdrawal Thresholds
3.3.1. CR Withdrawal Scenarios
3.3.2. POF Withdrawal Scenarios
4. Discussion
Considerations and Caveats of Water-Withdrawal Models
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Poff, N.L.; Allan, J.D.; Bain, M.B.; Karr, J.R.; Prestegaard, K.L.; Richter, B.D.; Sparks, R.E.; Stromberg, J.C. The natural flow regime. BioScience 1997, 47, 769–784. [Google Scholar] [CrossRef]
- Bunn, S.E.; Arthington, A.H. Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity. Environ. Manag. 2002, 30, 492–507. [Google Scholar] [CrossRef] [PubMed]
- Tharme, R.E. A global perspective on environmental flow assessment: Emerging trends in the development and application of environmental flow methodologies for rivers. River Res. Appl. 2003, 19, 397–441. [Google Scholar] [CrossRef]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.-I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.-H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef]
- Poff, N.L.; Zimmerman, J.K.H. Ecological responses to altered flow regimes: A literature review to inform the science and management of environmental flows. Freshw. Biol. 2010, 55, 194–205. [Google Scholar] [CrossRef]
- Annear, T.; Chisholm, I.; Beecher, H.; Locke, A.; Aarrestad, P.; Coomer, C.; Estes, C.; Hunt, J.; Jacobson, R.; Jobsis, G.; et al. Instream Flows for Riverine Resource Stewardship, Revised ed.; Instream Flow Council: Cheyenne, WY, USA, 2004. [Google Scholar]
- Rolls, R.J.; Leigh, C.; Sheldon, F. Mechanistic effects of low-flow hydrology on riverine ecosystems: Ecological principles and consequences of alteration. Freshw. Sci. 2012, 31, 1163–1186. [Google Scholar] [CrossRef]
- Walters, A.W. The importance of context dependence for understanding the effects of low-flow events on fish. Freshw. Sci. 2016, 35, 216–228. [Google Scholar] [CrossRef]
- Doll, P.; Fiedler, K.; Zhang, J. Global-scale analysis of river flow alterations due to water withdrawals and reservoirs. Hydrol. Earth Syst. Sci. 2009, 12, 2413–2432. [Google Scholar] [CrossRef]
- Robinson, J. Public-Supply Water Use and Self-Supplied Industrial Water Use in Tennessee, 2010; U.S. Geological Survey Scientific Investigation Report 2018–5009; U.S. Geological Survey: Reston, VA, USA, 2018; p. 30.
- Shen, Y.; Oki, T.; Utsumi, N.; Kanae, S.; Hanasaki, N. Projection of future world water resources under SRES scenarios: Water withdrawal. Hydrolog. Sci. J. 2008, 53, 11–33. [Google Scholar] [CrossRef]
- Meador, M.R.; Carlisle, D.M. Relations between altered streamflow variability and fish assemblages in eastern USA streams. River Res. Appl. 2012, 28, 1359–1368. [Google Scholar] [CrossRef]
- Phelan, J.; Cuffney, T.; Patterson, L.; Eddy, M.; Dykes, R.; Pearsall, S.; Goudreau, C.; Mead, J.; Tarver, F. Fish and invertebrate flow-biology relationships to support the determination of ecological flows for North Carolina. J. Am. Water Resour. As. 2017, 53, 42–55. [Google Scholar] [CrossRef]
- Freeman, M.C.; Marcinek, P.A. Fish assemblage responses to water withdrawals and water supply reservoirs in Piedmont streams. Environ. Manag. 2006, 38, 435–450. [Google Scholar] [CrossRef] [PubMed]
- Kanno, Y.; Vokoun, J.C. Evaluating effects of water withdrawals and impoundments on fish assemblages in southern New England streams, USA. Fish. Manag. Ecol. 2010, 17, 272–283. [Google Scholar] [CrossRef]
- Baron, J.S.; Poff, N.L.; Angermeier, P.L.; Dahm, C.N.; Gleick, P.H.; Hairston, N.G.; Jackson, R.B.; Johnston, C.A.; Richter, B.D.; Steinman, A.D. Meeting ecological and societal needs for freshwater. Ecol. Appl. 2002, 12, 1247–1260. [Google Scholar] [CrossRef]
- Arthington, A.; James, C.; Mackay, S.; Rolls, R.; Sternberg, D.; Barnes, A. Hydro-Ecological Relationships and Thresholds to Inform Environmental Flow Management; Science Report; International WaterCentre: Brisbane, Australia, 2012; p. 224. [Google Scholar]
- Arthington, A.H.; Bunn, S.E.; Poff, N.L.; Naiman, R.J. The challenge of providing environmental flow rules to sustain river ecosystems. Ecol. Appl. 2006, 16, 1311–1318. [Google Scholar] [CrossRef]
- Arthington, A.H.; Brizga, S.; Kennard, M. Comparative Evaluation of Environmental Flow Assessment Techniques: Best Practice Framework; LWRRDC Occasional Paper 25/98; Land and Water Resources Research and Development Corporation: Canberra, Australia, 1998; p. 26.
- Wheeler, K.; Wenger, S.J.; Freeman, M.C. States and rates: Complementary approaches to developing flow-ecology relationships. Freshw. Biol. 2018, 63, 906–916. [Google Scholar] [CrossRef]
- Acreman, M.C.; Overton, I.C.; King, J.; Wood, P.J.; Cowx, I.G.; Dunbar, M.J.; Kendy, E.; Young, W.J. The changing role of ecohydrological science in guiding environmental flows. Hydrol. Sci. J. 2014, 59, 433–450. [Google Scholar] [CrossRef]
- Poff, N.L.; Richter, B.D.; Arthington, A.H.; Bunn, S.E.; Naiman, R.J.; Kendy, E.; Acreman, M.; Apse, C.; Bledsoe, B.P.; Freeman, M.C.; et al. The ecological limits of hydrologic alteration (ELOHA): A new framework for developing regional environmental flow standards: Ecological limits of hydrologic alteration. Freshw. Biol. 2010, 55, 147–170. [Google Scholar] [CrossRef]
- Pastor, A.V.; Ludwig, F.; Biemans, H.; Hoff, H.; Kabat, P. Accounting for environmental flow requirements in global water assessments. Hydrol. Earth Syst. Sci. 2014, 18, 5041–5059. [Google Scholar] [CrossRef]
- Naiman, R.J.; Bunn, S.E.; Nilsson, C.; Petts, G.E.; Pinay, G.; Thompson, L.C. Legitimizing fluvial ecosystems as users of water: An overview. Environ. Manag. 2002, 30, 455–467. [Google Scholar] [CrossRef]
- Poff, N.L.; Allan, J.D.; Palmer, M.A.; Hart, D.D.; Richter, B.D.; Arthington, A.H.; Rogers, K.H.; Meyer, J.L.; Stanford, J.A. River flows and water wars: Emerging science for environmental decision making. Front. Ecol. Environ. 2003, 1, 298–306. [Google Scholar] [CrossRef]
- Davies, P.M.; Naiman, R.J.; Warfe, D.M.; Pettit, N.E.; Arthington, A.H.; Bunn, S.E. Flow–ecology relationships: Closing the loop on effective environmental flows. Mar. Freshw. Res. 2014, 65, 133–141. [Google Scholar] [CrossRef]
- Cartwright, J.; Caldwell, C.; Nebiker, S.; Knight, R. Putting flow–ecology relationships into practice: A decision-support system to assess fish community response to water-management scenarios. Water 2017, 9, 196. [Google Scholar] [CrossRef]
- Tennant, D.L. Instream Flow Regimens for Fish, Wildlife, Recreation and Related Environmental Resources. Fisheries 1976, 1, 6–10. [Google Scholar] [CrossRef]
- Richter, B.; Baumgartner, J.; Wigington, R.; Braun, D. How much water does a river need? Freshw. Biol. 1997, 37, 231–249. [Google Scholar] [CrossRef]
- Richter, B.D.; Davis, M.M.; Apse, C.; Konrad, C. A presumptive standard for environmental flow protection. River Res. Appl. 2012, 28, 1312–1321. [Google Scholar] [CrossRef]
- U.S. Environmental Protection Agency Water: Monitoring and Assessment, Section 5.1 Stream Flow. Available online: https://archive.epa.gov/water/archive/web/html/vms51.html (accessed on 2 January 2020).
- Farmer, W.H.; Over, T.M.; Kiang, J.E. Bias correction of simulated historical daily streamflow at ungauged locations by using independently estimated flow duration curves. Hydrol. Earth Syst. Sci. 2018, 22, 5741–5758. [Google Scholar] [CrossRef]
- Cook, C.N.; Hockings, M.; Carter, R. (Bill) Conservation in the dark? The information used to support management decisions. Front. Ecol. Environ. 2010, 8, 181–186. [Google Scholar] [CrossRef]
- Kendy, E.; Apse, C.; Blann, K. A Practical Guide to Environmental Flows for Policy and Planning, with Nine Case Studies in the United States; The Nature Conservancy: Arlington, VA, USA, 2012. [Google Scholar]
- Davies, S.P.; Jackson, S.K. The biological condition gradient: A descriptive model for interpreting change in aquatic ecosystems. Ecol. Appl. 2006, 16, 1251–1266. [Google Scholar] [CrossRef]
- Hain, E.F.; Kennen, J.G.; Caldwell, P.V.; Nelson, S.A.C.; Sun, G.; McNulty, S.G. Using regional scale flow-ecology modeling to identify catchments where fish assemblages are most vulnerable to changes in water availability. Freshw. Biol. 2018, 63, 928–945. [Google Scholar] [CrossRef]
- Knight, R.R.; Brian Gregory, M.; Wales, A.K. Relating streamflow characteristics to specialized insectivores in the Tennessee River Valley: A regional approach. Ecohydrology 2008, 1, 394–407. [Google Scholar] [CrossRef]
- Knight, R.R.; Gain, W.S.; Wolfe, W.J. Modelling ecological flow regime: An example from the Tennessee and Cumberland River basins. Ecohydrology 2012, 5, 613–627. [Google Scholar] [CrossRef]
- Knight, R.R.; Murphy, J.C.; Wolfe, W.J.; Saylor, C.F.; Wales, A.K. Ecological limit functions relating fish community response to hydrologic departures of the ecological flow regime in the Tennessee River basin, United States. Ecohydrology 2014, 1262–1280. [Google Scholar] [CrossRef]
- Murphy, J.C.; Knight, R.R.; Wolfe, W.J.; S. Gain, W. Predicting ecological flow regime at ungaged sites: A comparison of methods. River Res. Appl. 2012, 29, 660–669. [Google Scholar] [CrossRef]
- Knight, R.R.; Cartwright, J.M.; Ladd, D.E. Streamflow and Fish Community Diversity Data for Use in Developing Ecological Limit Functions for the Cumberland Plateau, Northeastern Middle Tennessee and Southwestern Kentucky, 2016; U.S. Geological Survey data release; U.S. Geological Survey: Reston, VA, USA, 2016. Available online: http://dx.doi.org/10.5066/F7JH3J83 (accessed on 1 September 2018).
- Omernik, J.M. Map Supplement: Ecoregions of the Conterminous United States. Ann. Assoc. Am. Geogr. 1987, 77, 118–125. [Google Scholar] [CrossRef]
- Abell, R.; Olson, D.; Dinerstein, E.; Hurley, P.; Diggs, J.; Eichbaum, W.; Walters, S.; Wettengel, W.; Allnutt, T.; Loucks, C.; et al. Freshwater Ecoregions of North America: A Conservation Assessment; Island Press: Washington, DC, USA, 2000. [Google Scholar]
- Elkins, D.C.; Sweat, S.C.; Hill, K.S.; Kuhajda, B.R.; George, A.L.; Wenger, S.J. The Southeastern Aquatic Biodiversity Conservation Strategy. Final Report; University of Georgia River Basin Center: Athens, Greece, 2016; p. 237. [Google Scholar]
- Fenneman, N.M. Physiographic Divisions of the United States. Ann. Assoc. Am. Geogr. 1928, 18, 261. [Google Scholar] [CrossRef]
- Kennard, M.J.; Mackay, S.J.; Pusey, B.J.; Olden, J.D.; Marsh, N. Quantifying uncertainty in estimation of hydrologic metrics for ecohydrological studies. River Res. Appl. 2010, 137–156. [Google Scholar] [CrossRef]
- U.S. Geological Survey (USGS) USGS Water Data for the Nation: U.S. Geological Survey National Water Information System Database. Available online: http://dx.doi.org/10.5066/F7P55KJN (accessed on 1 September 2018).
- Hirsch, R.; De Cicco, L. User Guide to Exploration and Graphics for RivEr Trends (EGRET) and dataRetrieval: R Packages for Hydrologic Data; U.S. Geological Survey Techniques and Methods book 4, chap. A10; U.S. Geological Survey: Reston, VA, USA, 2015; p. 93.
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019.
- Robinson, J.A. Water Use in Tennessee, 2010; U.S. Geological Survey data release; U.S. Geological Survey: Reston, VA, USA, 2017. Available online: https://doi.org/10.5066/F7V9868K (accessed on 1 September 2018).
- Mills, J.; Blodgett, D. EflowStats: Hydrologic Indicator and Alteration Stats. R Package Version 5.0.0; 2017; Available online: https://github.com/USGS-R/EflowStats (accessed on 1 September 2017).
- Buchanan, B.; McManamay, R.A.; Auerbach, D.; Fuka, D.; Walter, M. Environmental Flow Analysis for the Marcellus Shale Region; Appalachian Landscape Conservation Cooperative: Shepherdstown, WV, USA, 2015; p. 170. [Google Scholar]
- Driver, L.J. Ecological Flow Analyses Results: Streamflow Characteristics, Predicted Fish Responses, and Ecological Withdrawal Thresholds for Select Stream Sites within the Cumberland and Tennessee River Basins; U.S. Geological Survey data release; U.S. Geological Survey: Reston, VA, USA, 2019. Available online: https://doi.org/10.5066/F7Q23Z4B (accessed on 1 September 2019).
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The River Continuum Concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Poff, N.L.; Ward, J.V. Implications of Streamflow Variability and Predictability for Lotic Community Structure: A Regional Analysis of Streamflow Patterns. Can. J. Fish. Aquat. Sci. 1989, 46, 1805–1818. [Google Scholar] [CrossRef]
- Shank, M.K.; Stauffer, J.R. Land use and surface water withdrawal effects on fish and macroinvertebrate assemblages in the Susquehanna River basin, USA. J. Freshw. Ecol. 2015, 30, 229–248. [Google Scholar] [CrossRef]
- Flannery, M.S.; Peebles, E.B.; Montgomery, R.T. A percent-of-flow approach for managing reductions of freshwater inflows from unimpounded rivers to Southwest Florida estuaries. Estuaries 2002, 25, 1318–1332. [Google Scholar] [CrossRef]
- Acreman, M.C.; Ferguson, A.J.D. Environmental flows and the European Water Framework Directive: Environmental flows and WFD. Freshw. Biol. 2010, 55, 32–48. [Google Scholar] [CrossRef]
- Zorn, T.G.; Seelbach, P.W.; Rutherford, E.S.; Wills, T.; Cheng, S.; Wiley, M. A Regional-Scale Habitat Suitability Model to Assess the Effects of Flow Reduction on Fish Assemblages in Michigan Streams; Michigan Department of Natural Resources: Ann Arbor, MI, USA, 2008; pp. 871–895.
- Driver, L.J.; Hoeinghaus, D.J. Spatiotemporal dynamics of intermittent stream fish metacommunities in response to prolonged drought and reconnectivity. Mar. Freshw. Res. 2016, 67, 1667–1679. [Google Scholar] [CrossRef]
- Loftin, K.A.; Clark, J.M.; Journey, C.A.; Kolpin, D.W.; Van Metre, P.C.; Carlisle, D.; Bradley, P.M. Spatial and temporal variation in microcystin occurrence in wadeable streams in the southeastern United States: Microcystins in southeastern US streams. Environ. Toxicol. Chem. 2016, 35, 2281–2287. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.L.; Dubrovsky, N.M.; Foster, G.M.; King, L.R.; Loftin, K.A.; Rosen, B.H.; Stelzer, E.A. Cyanotoxin occurrence in large rivers of the United States. Inland Waters 2020, 10, 109–117. [Google Scholar] [CrossRef]
- North Carolina Ecological Flows Science Advisory Board. Recommendations for Estimating Flows to Maintain Ecological Integrity in Streams and Rivers in North Carolina; North Carolina Department of Environmental Quality: Raleigh, NC, USA, 2013. Available online: https://files.nc.gov/ncdeq/Water%20Resources/files/eflows/sab/EFSAB_Final_Report_to_NCDENR.pdf (accessed on 4 January 2019).
- Rosenfeld, J.S. Developing flow-ecology relationships: Implications of nonlinear biological responses for water management. Freshw. Biol. 2017, 62, 1305–1324. [Google Scholar] [CrossRef]
- Olden, J.D.; Poff, N.L. Redundancy and the choice of hydrologic indices for characterizing streamflow regimes. River Res. Appl. 2003, 19, 101–121. [Google Scholar] [CrossRef]
- Jackson, D.A.; Peres-Neto, P.R.; Olden, J.D. What controls who is where in freshwater fish communities–the roles of biotic, abiotic, and spatial factors. Can. J. Fish. Aquat. Sci. 2001, 58, 157–170. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flow Category | Streamflow Characteristic (SFC) | Definition (Units) | Eco-Region |
|---|---|---|---|
| Magnitude | MA41: mean annual runoff | Annual mean streamflow divided by the drainage area (ft3s−1mi−2) | RV, CP |
| AMH10: maximum October streamflow | Maximum October flow across period of record divided by watershed area (ft3s−1mi−2) | BR, CP, IP | |
| LRA7: rate of streamflow recession | Log of the median change in log of flow for days that the change is negative across the entire flow record (flow units per day) | IP | |
| Ratio | LDH13: average 30-day maximum | Log of the average over period of record of annual maximum 30-day moving average flows divided by median for entire record | CP |
| ML20: base flow | Divide daily flow record into 5-day blocks. Assign minimum (min.) flow for the block as a base flow if 90% of that min. flow is less than the min. flows for blocks on either side; otherwise, set to zero. Fill in zero values using linear interpolation. Compute total flow and total base flow for entire record. ML20 is total flow: total base flow (ratio) | CP | |
| TA1: constancy | Measure the stability of flow regimes by dividing daily flows into predetermined flow classes | RV, CP, IP | |
| Frequency | FH6: frequency of moderate flooding | Average number of high-flow events per year ≥ 3 times the median annual flow for the period of record (number per year) | IP |
| Variability | LDL6: variability of annual minimum daily average streamflow | Log of the standard deviation for the annual minimum daily average streamflow. Multiply by 100 and divide by the mean streamflow for the period (%) | CP |
| LDH16: variability in high-pulse duration | Log of the standard deviation for the yearly average high-flow pulse durations (daily flow > 75th percentile) (%) | RV | |
| FL2: variability in low-pulse count | Coefficient of variation for the number of annual occurrences of daily flows < 25th percentile | RV | |
| Timing | TL1: annual min. flow | Date of annual mininum (min.) flow occurrence (Julian day) | CP, IP |
| Constant-Rate Withdrawal | Percent-of-Flow Withdrawal | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Eco-Region | Flow Category | SFC | MFL0 | MFL5 | MFL10 | MFL30 | MFL0 | MFL5 | MFL10 | MFL30 |
| BR | Magnitude | AMH10 | 0.04 | 0.04 | 0.04 | 0.04 | 0.06 | 0.06 | 0.06 | 0.06 |
| RV | Magnitude | MA41 | 0.32 | 0.30 | 0.28 | 0.21 | 0.28 | 0.28 | 0.28 | 0.24 |
| Ratio | TA1 | 0.60 | 0.44 | 0.30 | 0.16 | 0.16 | 0.15 | 0.13 | 0.09 | |
| Variability | LDH16 | 0.01 | 0.01 | 0.01 | 0.02 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Variability | FL2 | 0.02 | 3.17 | 3.09 | 0.24 | 0.00 | 0.00 | 0.03 | 0.23 | |
| CP | Magnitude | MA41 | 0.16 | 0.14 | 0.13 | 0.10 | 0.20 | 0.19 | 0.19 | 0.16 |
| Magnitude | AMH10 | 0.01 | 0.01 | 0.01 | 0.01 | 0.02 | 0.02 | 0.02 | 0.02 | |
| Ratio | LDH13 | 0.29 | 0.34 | 0.30 | 0.14 | 0.23 | 0.23 | 0.23 | 0.17 | |
| Ratio | ML20 | 0.08 | 0.18 | 0.15 | 0.07 | 0.16 | 0.15 | 0.14 | 0.09 | |
| Ratio | TA1 | 0.27 | 0.05 | 0.03 | 0.02 | 0.12 | 0.03 | 0.03 | 0.02 | |
| Variability | LDL6 | 1.16 | 0.05 | 0.00 | 0.00 | 0.00 | 0.05 | 0.00 | 0.00 | |
| Timing | TL1 | 3.80 | 0.02 | 0.00 | 0.00 | 0.00 | 0.01 | 0.00 | 0.00 | |
| IP | Magnitude | AMH10 | 0.03 | 0.03 | 0.03 | 0.02 | 0.03 | 0.03 | 0.03 | 0.03 |
| Magnitude | LRA7 | 1.33 | 0.95 | 0.68 | 0.19 | 0.11 | 0.13 | 0.14 | 0.10 | |
| Ratio | TA1 | 0.63 | 0.35 | 0.23 | 0.11 | 0.15 | 0.11 | 0.08 | 0.06 | |
| Frequency | FH6 | 0.17 | 0.15 | 0.14 | 0.13 | 0.05 | 0.05 | 0.05 | 0.07 | |
| Timing | TL1 | 4.59 | 1.34 | 0.46 | 0.00 | 0.00 | 0.10 | 0.12 | 0.01 | |
| Constant-Rate Withdrawal Mean Change in Richness | Percent-of-Flow Withdrawal Mean Change in Richness | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Eco-Region | Fish Group 1,2,3 | Streamflow Characteristic (s) | MFL0 | MFL5 | MFL10 | MFL30 | MFL0 | MFL5 | MFL10 | MFL30 |
| BR | All species | AMH10 | −0.28 | −0.28 | −0.28 | −0.26 | −0.40 | −0.40 | −0.40 | −0.39 |
| RV | All species | FL2 | −0.10 | −4.89 | −5.57 | −1.01 | 0.00 | 0.00 | −0.19 | −0.85 |
| Specialized insectivores | TA1 + LDH16 | −0.71 | 0.94 | 1.11 | 0.15 | 0.51 | 0.67 | 0.81 | −0.29 | |
| CP | All species | TL1 | −12.69 | −0.02 | 0.00 | 0.00 | 0.00 | −0.09 | 0.00 | 0.00 |
| Specialized insectivores | AMH10 + TA1 + TL1 | −5.54 | 0.16 | 0.01 | 0.04 | −0.25 | −0.16 | −0.07 | 0.03 | |
| IP | All species | AMH10 + TL1 | −21.50 | −8.13 | −2.97 | −0.05 | −0.09 | −0.52 | −0.91 | −0.08 |
| Specialized insectivores | AMH10 + TA1 | −0.12 | 0.53 | 0.59 | 0.31 | −0.02 | 0.11 | 0.21 | −0.01 | |
| Mean Loss in Richness (Number of Species) | Mean Percent Loss in Richness (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 10% Flow Withdrawal | 20% Flow Withdrawal | 10% Flow Withdrawal | 20% Flow Withdrawal | ||||||
| Eco-Region | Number of Fish Groups1 | Range | Mean | Range | Mean | Range | Mean | Range | Mean |
| BR | 3 | 0.11–0.32 | 0.19 | 0.16–0.43 | 0.27 | 0.85–0.99 | 0.93 | 1.20–1.34 | 1.29 |
| RV | 2 | 0.34–0.43 | 0.39 | 0.61–0.81 | 0.71 | 3.86–10.59 | 7.23 | 7.31–13.19 | 10.25 |
| CR | 9 | 0.02–0.31 | 0.13 | 0.05–0.61 | 0.30 | 0.75–12.95 | 3.45 | 1.28–18.89 | 7.67 |
| IP | 10 | 0.01–0.66 | 0.15 | 0.02–1.34 | 0.29 | 0.15–7.12 | 1.34 | 0.22–13.00 | 2.57 |
| Overall Mean | 0.17 | 0.33 | 2.57 | 4.96 | |||||
| Constant-Rate (CR) Ecological Withdrawal Thresholds (m3/s) 1 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MFL0 | MFL5 | MFL10 | MFL30 | |||||||||
| Eco-Region | n 2 | Range | Mean | n | Range | Mean | n | Range | Mean | n | Range | Mean |
| BR | 6 | 0.64−0.68 | 0.67 | 6 | 0.64−0.68 | 0.68 | 6 | 0.67−0.72 | 0.70 | 6 | 0.74−0.79 | 0.77 |
| RV | 47 | 0.20−0.81 | 0.62 | 73 | 0.24−0.68 | 0.51 | 80 | 0.06−0.64 | 0.39 | 52 | 0.23−0.57 | 0.34 |
| CP | 98 | 0.01−0.92 | 0.16 | 25 | 0.03−1.01 | 0.23 | 23 | 0.01−0.92 | 0.35 | 11 | 0.03−0.85 | 0.40 |
| IP | 267 | 0.17−0.37 | 0.23 | 187 | 0.10−0.37 | 0.20 | 118 | 0.17−0.37 | 0.26 | 33 | 0.17−1.33 | 0.67 |
| Percent-of-flow (POF) ecological withdrawal thresholds (%) 1 | ||||||||||||
| MFL0 | MFL5 | MFL10 | MFL30 | |||||||||
| Eco-region | n 2 | Range | Mean | n | Range | Mean | n | Range | Mean | n | Range | Mean |
| BR | 6 | 23.5−27.0 | 25.5 | 6 | 23.5−27.0 | 25.5 | 6 | 23.5−27.0 | 25.5 | 6 | 25.0−28.0 | 26.8 |
| RV | 17 | 6.0−6.5 | 6.3 | 16 | 6.5−14.5 | 10.5 | 25 | 1.0−32.5 | 22.9 | 49 | 6.0−16.0 | 13.0 |
| CP | 33 | 4.0−16.0 | 9.4 | 29 | 4.0−28.0 | 15.7 | 22 | 4.0−29.0 | 13.7 | 14 | 2.0−17.0 | 7.6 |
| IP | 63 | 3.5−31.0 | 18.8 | 102 | 9.0−31.0 | 22.6 | 76 | 2.0−33.5 | 18.6 | 23 | 1.0−37.0 | 14.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Driver, L.J.; Cartwright, J.M.; Knight, R.R.; Wolfe, W.J. Species-Richness Responses to Water-Withdrawal Scenarios and Minimum Flow Levels: Evaluating Presumptive Standards in the Tennessee and Cumberland River Basins. Water 2020, 12, 1334. https://doi.org/10.3390/w12051334
Driver LJ, Cartwright JM, Knight RR, Wolfe WJ. Species-Richness Responses to Water-Withdrawal Scenarios and Minimum Flow Levels: Evaluating Presumptive Standards in the Tennessee and Cumberland River Basins. Water. 2020; 12(5):1334. https://doi.org/10.3390/w12051334
Chicago/Turabian StyleDriver, Lucas J., Jennifer M. Cartwright, Rodney R. Knight, and William J. Wolfe. 2020. "Species-Richness Responses to Water-Withdrawal Scenarios and Minimum Flow Levels: Evaluating Presumptive Standards in the Tennessee and Cumberland River Basins" Water 12, no. 5: 1334. https://doi.org/10.3390/w12051334
APA StyleDriver, L. J., Cartwright, J. M., Knight, R. R., & Wolfe, W. J. (2020). Species-Richness Responses to Water-Withdrawal Scenarios and Minimum Flow Levels: Evaluating Presumptive Standards in the Tennessee and Cumberland River Basins. Water, 12(5), 1334. https://doi.org/10.3390/w12051334