Attenuation of Tides and Surges by Mangroves: Contrasting Case Studies from New Zealand



Abstract
1. Introduction
2. Materials and Methods
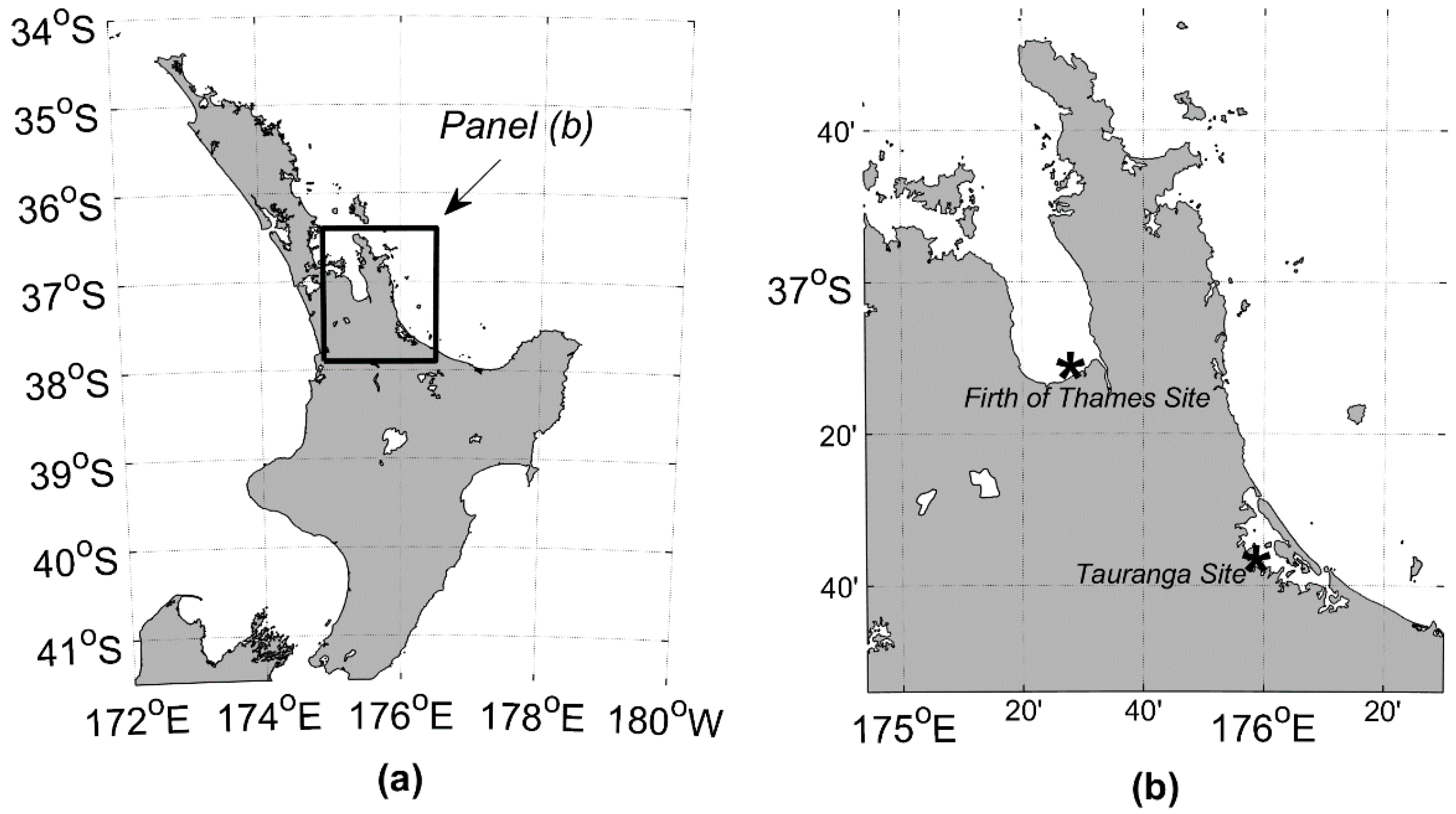
2.1. Study Sites
2.1.1. Firth of Thames
2.1.2. Tauranga Site
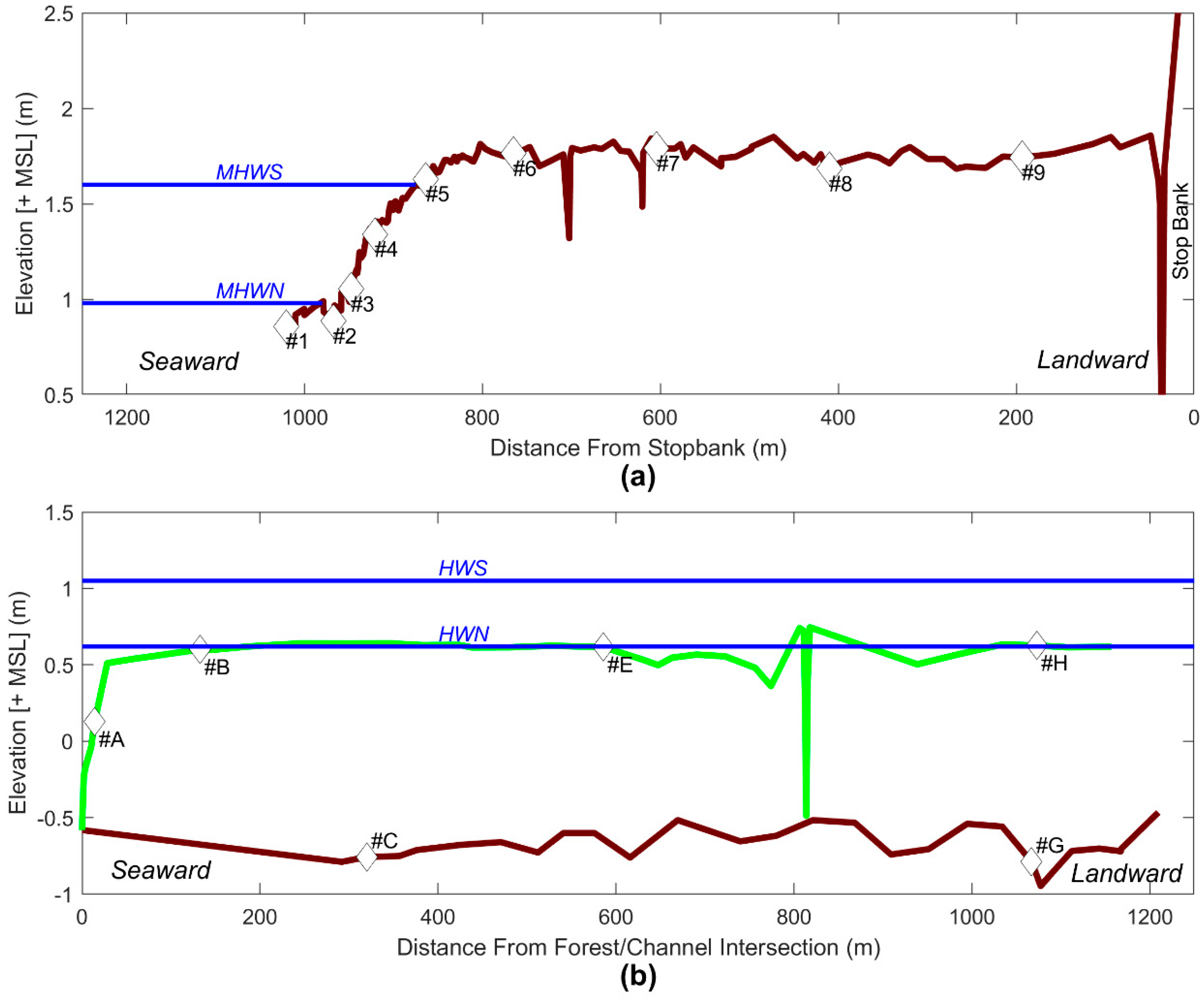
2.2. Field Data Collection
2.2.1. Vegetation Survey
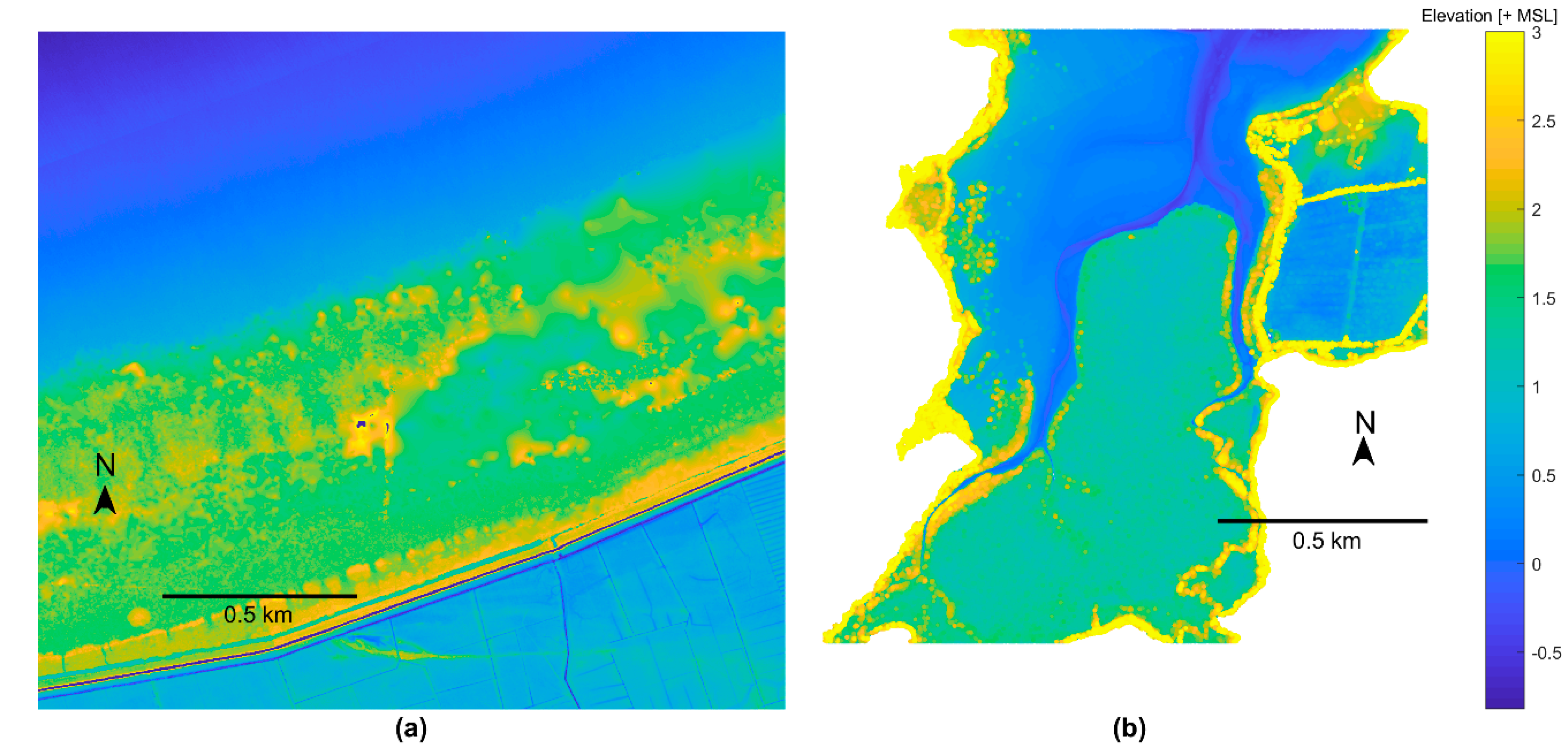
2.2.2. Bathymetry
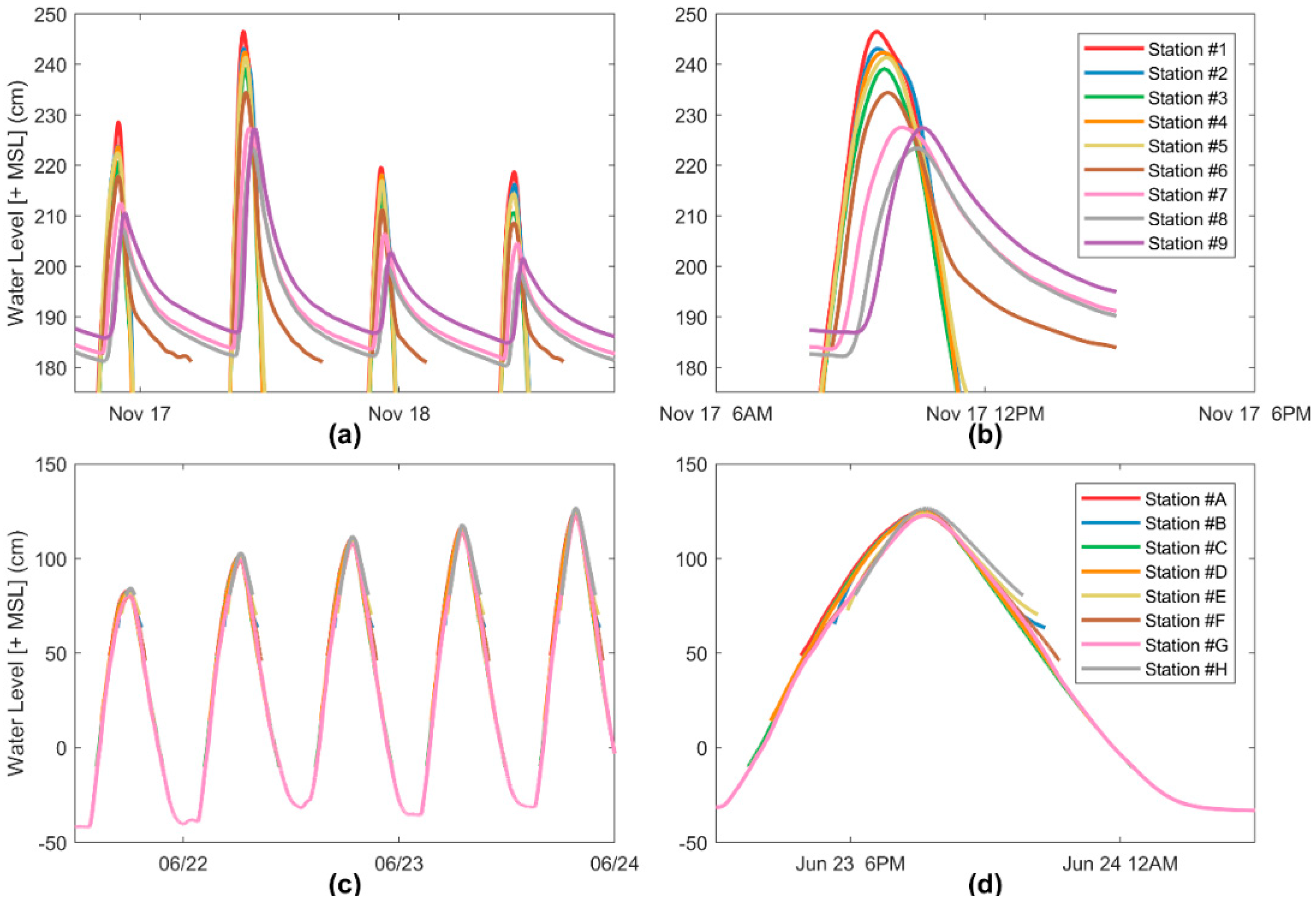
2.2.3. Water Level
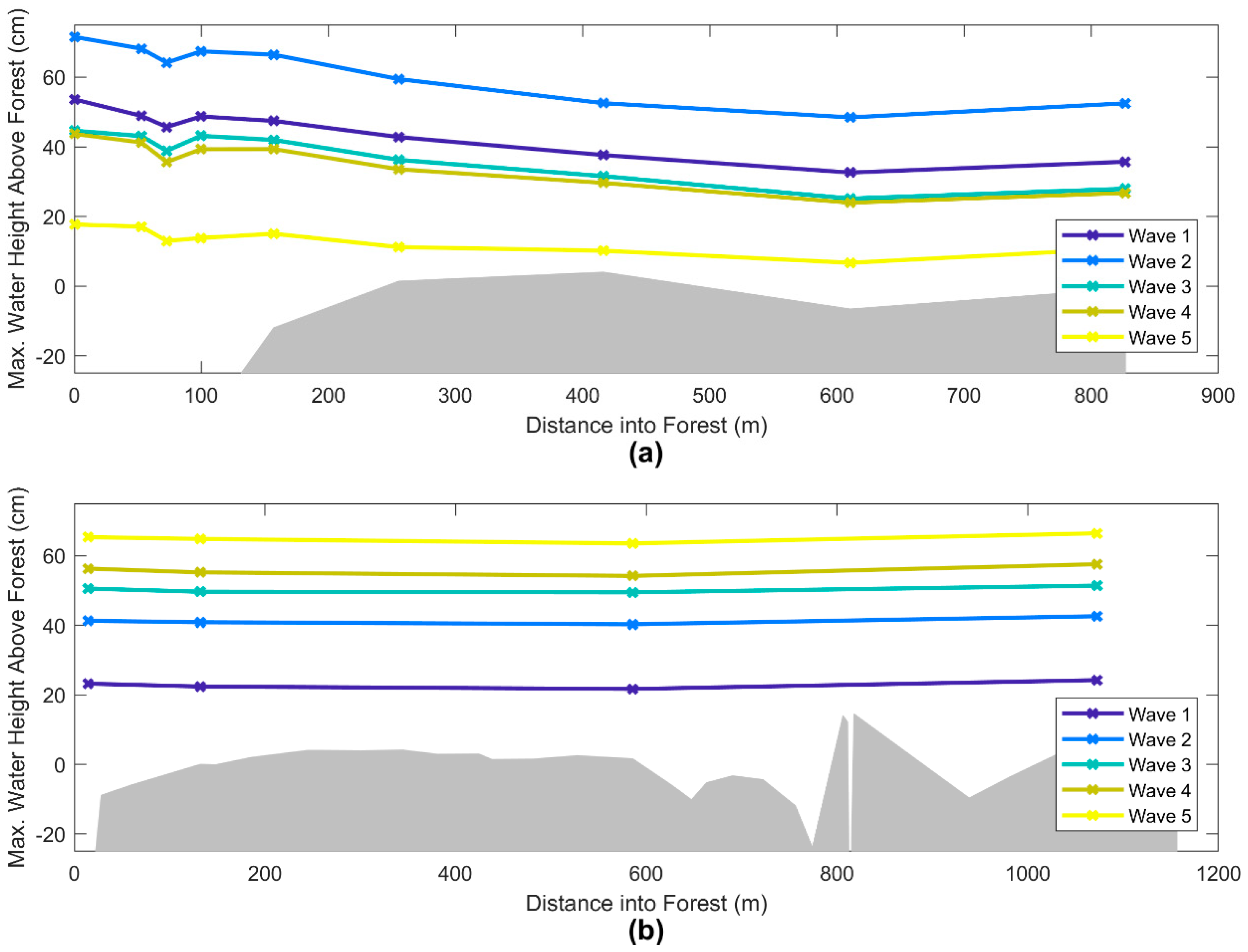
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization (FAO). The World’s Mangroves 1980–2005; Food and Agriculture Organization of the United Nations: Rome, Italy, 2007; ISBN 978-92-5-105856-5. [Google Scholar]
- McLeod, E.; Chmura, G.L.; Bouillon, S.; Salm, R.; Bjork, M.; Duarte, C.M.; Lovelock, C.E.; Schlesinger, W.H.; Silliman, B.R. A blueprint for blue carbon: Toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2. Front. Ecol. Environ. 2011, 9, 552–560. [Google Scholar] [CrossRef]
- van Maanen, B.; Coco, G.; Bryan, K.R. On the ecogeomorphological feedbacks that control tidal channel network evolution in a sandy mangrove setting. Proc. R. Soc. A 2015, 471, 20150115. [Google Scholar] [CrossRef] [PubMed]
- Giri, C.; Ochieng, E.; Tieszen, L.L.; Zhu, Z.; Singh, A.; Loveland, T.; Masek, J.; Duke, N. Status and distribution of mangrove forests of the world using earth observation satellite data. Glob. Ecol. Biogeogr. 2011, 20, 154–159. [Google Scholar] [CrossRef]
- Wahl, T.; Haigh, I.D.; Nicholls, R.J.; Arns, A.; Dangendorf, S.; Hinkel, J.; Slangen, A.B.A. Understanding extreme sea levels for broad-scale coastal impact and adaptation analysis. Nat. Commun. 2017, 8, 16075. [Google Scholar] [CrossRef] [PubMed]
- Kroeker, K.J.; Reguero, B.G.; Rittelmeyer, P.; Beckd, M.W. Ecosystem Service and Coastal Engineering Tools for Coastal Protection and Risk Reduction. In Managing Coasts with Natural Solutions; World Bank: Washington, DC, USA, 2016. [Google Scholar]
- Small, C.; Nicholls, R.J. A global analysis of human settlement in coastal zones. J. Coast. Res. 2003, 19, 584–599. [Google Scholar]
- Plant, N.G.; Griggs, G.B. Interactions between nearshore processes and beach morphology near a seawall. J. Coast. Res. 1992, 8, 183–200. [Google Scholar]
- Airoldi, L.; Abbiati, M.; Beck, M.W.; Hawkins, S.J.; Jonsson, P.R.; Martin, D.; Moschellad, P.S.; Sundelofg, A.; Thompsonf, R.C.; Aberg, P. An ecological perspective on the deployment and design of low-crested and other hard coastal defence structures. Coast. Eng. 2005, 52, 1073–1087. [Google Scholar] [CrossRef]
- Guannel, G.; Ruggiero, P.; Faries, J.; Arkema, K.; Pinsky, M.; Gelfenbaum, G.; Guerry, A.; Kim, C.K. Integrated modeling framework to quantify the coastal protection services supplied by vegetation. J. Geophys. Res. Ocean. 2015, 120, 324–345. [Google Scholar] [CrossRef]
- Temmerman, S.; Meire, P.; Bouma, T.J.; Herman, P.; Ysebaert, T.; De Vriend, H.J. Ecosystem-based coastal defence in the face of global change. Nature 2013, 504, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Alongi, D.M. Mangrove forests: Resilience, protection from tsunamis, and responses to global climate change. Estuar. Coast. Shelf Sci. 2008, 76, 1–13. [Google Scholar] [CrossRef]
- Henderson, S.M.; Norris, B.K.; Mullarney, J.C.; Bryan, K.R. Wave-frequency flows within a near-bed vegetation canopy. Cont. Shelf Res. 2017, 147, 91–101. [Google Scholar] [CrossRef]
- Massel, S.; Furukawa, K.; Brinkman, R. Surface wave propagation in mangrove forests. Fluid Dyn. Res. 1999, 24, 219–249. [Google Scholar] [CrossRef]
- Krauss, K.W.; Doyle, T.W.; Doyle, T.J.; Swarzenski, C.M.; From, A.S.; Day, R.H.; Conner, W.H. Water level observations in mangrove swamps during two hurricanes in Florida. Wetlands 2009, 29, 142–149. [Google Scholar] [CrossRef]
- Zhang, K.Q.; Liu, H.Q.; Li, Y.P.; Xu, H.Z.; Shen, J.; Rhome, J.; Smith, T.J. The role of mangroves in attenuating storm surges. Estuar. Coast. Shelf Sci. 2012, 102–103, 11–23. [Google Scholar] [CrossRef]
- U.S. Army Corps of Engineers. Hurricane Study for Morgan City, Louisiana and Vicinity; United States Congress Serial Set: Washington, DC, USA, 1965.
- McIvor, A.; Spencer, T.; Möller, I.; Spalding, M. Storm surge reduction by mangroves. In Natural Coastal Protection Series: Report 2; The Nature Conservancy and Wetlands International: Cambridge, UK, 2012. [Google Scholar]
- Chen, R.; Twilley, R.R. A gap dynamic model of mangrove forest development along gradients of soil salinity and nutrient resources. J. Ecol. 1998, 86, 37–51. [Google Scholar] [CrossRef]
- Mullarney, J.C.; Henderson, S.M. Flows Within Marine Vegetaion Canopies in Advances. In Coastal Hydraulics; Panchange, V., Kaihatu, J., Eds.; World Scientific Publishing: Singapore, 2018; pp. 1–46. [Google Scholar]
- Nepf, H. Hydrodynamics of vegetated channels. J. Hydraul. Res. 2012, 50, 262–279. [Google Scholar] [CrossRef]
- Folkard, A.M. Vegetated flows in their environmental context: A review. Proc. Inst. Civ. Eng.-Eng. Comput. Mech. 2011, 164, 3–24. [Google Scholar] [CrossRef]
- Horstman, E.M.; Lundquist, C.J.; Bryan, K.R.; Bulmer, R.H.; Mullarney, J.C.; Stokes, D.J. The dynamics of expanding mangroves in New Zealand. In Threats to Mangrove Forests; Springer: New York, NY, USA, 2018; pp. 23–51. [Google Scholar] [CrossRef]
- Swales, A.; Bentley, S.J.; Lovelock, C.; Bell, R.G. Sediment processes and mangrove-habitat expansion on a rapidly-prograding muddy coast, New Zealand. In Proceedings of the Coastal Sediments'07, Processing of the Sixth International Symposium on Coastal Engineering and Science of Coastal Sediment Process, New Orleans, LA, USA, 13–17 May 2007. [Google Scholar]
- Swales, A.; Bentley, S.J.; Lovelock, C.E. Mangrove-forest evolution in a sediment-rich estuarine system: Opportunists or agents of geomorphic change? Earth Surf. Proc. Landf. 2015, 40, 1672–1687. [Google Scholar] [CrossRef]
- Heath, R. A review of the physical oceanography of the seas around New Zealand—1982. N. Z. J. Mar. Freshw. Res. 1985, 19, 79–124. [Google Scholar] [CrossRef]
- Tay, H.W.; Bryan, K.R.; de Lange, W.P.; Pilditch, C.A. The hydrodynamics of the southern basin of Tauranga Harbour. N. Z. J. Mar. Freshw. Res. 2013, 47, 249–274. [Google Scholar] [CrossRef]
- Healy, T.R.; Cole, R.; de Lange, W. Geomorphology and ecology of New Zealand shallow estuaries and shorelines. In Estuarine Shores; John Wiley and Sons: New York, NY, USA, 1996. [Google Scholar]
- Park, S.G. Aspects of Mangrove Distribution and Abundance in Tauranga Harbour; Environment Bay of Plenty: Whakatane, New Zealand, 2004.
- Nepf, H. Vegetated flow dynamics. In The Ecogeomorphology of Tidal Marshes; American Geophysical Union: Washington, DC, USA, 2004; pp. 137–163. [Google Scholar]
- Chen, Y.N.; Li, Y.; Cai, T.L.; Thompson, C.; Li, Y. A comparison of biohydrodynamic interaction within mangrove and saltmarsh boundaries. Earth Surf. Proc. Landf. 2016, 41, 1967–1979. [Google Scholar] [CrossRef]
- Norris, B.K.; Mullarney, J.C.; Bryan, K.R.; Henderson, S.M. The effect of pneumatophore density on turbulence: A field study in a Sonneratia-dominated mangrove forest, Vietnam. Cont. Shelf Res. 2017, 147, 114–127. [Google Scholar] [CrossRef]
- Horstman, E.; Bryan, K.; Mullarney, J.; Pilditch, C.; Eager, C. Are flow-vegetation interactions well represented by mimics? A case study of mangrove pneumatophores. Adv. Water Resour. 2018, 111, 360–371. [Google Scholar] [CrossRef]
- Mullarney, J.C.; Henderson, S.M.; Reyns, J.A.; Norris, B.K.; Bryan, K.R. Spatially varying drag within a wave-exposed mangrove forest and on the adjacent tidal flat. Cont. Shelf Res. 2017, 147, 102–113. [Google Scholar] [CrossRef]
- Green, J.C. Comparison of blockage factors in modelling the resistance of channels containing submerged macrophytes. River Res. Appl. 2005, 21, 671–686. [Google Scholar] [CrossRef]
- Horstman, E.M.; Dohmen-Janssen, C.M.; Bouma, T.J.; Hulscher, S.J.M.H. Tidal-scale flow routing and sedimentation in mangrove forests: Combining field data and numerical modelling. Geomorphology 2015, 228, 244–262. [Google Scholar] [CrossRef]
- Mazda, Y.; Kanazawa, N.; Wolanski, E. Tidal asymmetry in mangrove creeks. Hydrobiologia 1995, 295, 51–58. [Google Scholar] [CrossRef]
- Mei, C.C.; Chan, I.C.; Liu, P.L.F.; Huang, Z.; Zhang, W. Long waves through emergent coastal vegetation. J. Fluid Mech. 2011, 687, 461–491. [Google Scholar] [CrossRef]
- Bryan, K.R.; Nardin, W.; Mullarney, J.C.; Fagherazzi, S. The role of cross-shore tidal dynamics in controlling intertidal sediment exchange in mangroves in Cù Lao Dung, Vietnam. Cont. Shelf Res. 2017, 147, 128–143. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Instrument | Sampling | Temperature Dependence | Depth Measurement Uncertainty | |
|---|---|---|---|---|---|
| Regime | Details | Correction Applied | (cm) | ||
| #1 | Nortek Aquadopp | Burst | 212 samples at 8 Hz every 15 min | Y | <10 |
| #2 | Nortek Vector | Burst | 7.5 min sampling at 16 Hz every 15 min | Y | <5 |
| #3 | Nortek Aquadopp | Burst | 212 samples at 8 Hz every 15 min | Y | <10 |
| #4 | Nortek Aquadopp | Burst | 212 samples at 8 Hz every 15 min | Y | <10 |
| #5 | RBR Concerto | Continuous | 4 Hz | N | <1 |
| #6 | RBR Duet | Continuous | 8 Hz | N | <1 |
| #7 | Solinst Levelogger | Continuous | 1/60 Hz | N | <1 |
| #8 | Solinst Levelogger | Continuous | 1/60 Hz | N | <1 |
| #9 | Solinst Levelogger | Continuous | 1/60 Hz | N | <1 |
| Weather Station | Continuous | 1/5 min | N | <1 | |
| Station | Instrument | Sampling | Temperature Dependence | Depth Measurement Uncertainty | |
|---|---|---|---|---|---|
| Regime | Details | Correction Applied | (cm) | ||
| #A | RBR Solo | Continuous | 8 Hz | N | <1 |
| #B | RBR Solo | Continuous | Every 2 min Average 1 min of data sampled at 4 Hz | N | <1 |
| #C | RBR Duet | Continuous | 8 Hz | N | <1 |
| #D | RBR Duet | Continuous | 8 Hz | N | <1 |
| #E | RBR Solo | Continuous | Every 2 min Average 1 min of data sampled at 4 Hz | N | <1 |
| #F | RBR Solo | Continuous | 8 Hz | N | <1 |
| #G | RBR Solo | Continuous | 8 Hz | N | <1 |
| #H | RBR Solo | Continuous | 8 Hz | N | <1 |
| Weather Station | Continuous | 1/5 min | N | <1 | |
| Location | Trees | Pneumatophores | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Description | Distance from Fringe | Density | Height | Diameter | Frontal Area | Density | Height | Diameter | Frontal Area |
| (m) | (# per m2) | (m) | (cm) | (m−1) | (# per m2) | (cm) | (cm) | (m−1) | |
| Mudflat | −49 | 0.1 | 0.6 | 1.6 | 0.00 | 0 | 0 | 0 | 0 |
| Fringe (Front) | 0 | 0.1 | 2.3 | 6.4 | 0.01 | 126 | 25.4 (8.5) | 0.6 (0.2) | 0.76 |
| Fringe (Back) | 22 | 1.0 | 1.9 (1.1) | 4.8 (5.4) | 0.05 | 385 | 22.9 (9.1) | 0.6 (0.2) | 2.31 |
| Forest | 36 | 1.0 | 2.9 (0.3) | 5.8 (3.1) | 0.06 | 386 | 17.0 (6.9) | 0.5 (0.2) | 1.93 |
| Forest | 114 | 6.4 | 2.3 (0.2) | 3.0 (0.7) | 0.19 | 322 | 12.5 (5.6) | 0.6 (0.2) | 1.93 |
| Forest | 166 | 11.4 | 1.5 (0.3) | 2.8 (1.2) | 0.32 | 357 | 7.7 (4.4) | 0.6 (0.2) | 2.14 |
| Forest | 262 | 12.2 | 1.2 (0.1) | 1.9 (0.4) | 0.23 | 481 | 8.3 (4.1) | 0.6 (0.1) | 2.89 |
| Forest | 376 | 6.4 | 3.1 (0.1) | 3.6 (1.1) | 0.23 | 342 | 8.7 (5.4) | 0.5 (0.1) | 1.71 |
| Forest | 476 | 1.4 | 3.8 (0.3) | 4.2 (1.1) | 0.06 | 330 | 10.5 (5.5) | 0.6 (0.1) | 1.98 |
| Forest | 561 | 3.6 | 1.6 (0.2) | 3.9 (2.2) | 0.14 | 570 | 12.0 (6.1) | 0.5 (0.1) | 2.85 |
| Forest | 667 | 5 | 2.6 (0.3) | 3.8 (0.6) | 0.19 | 482 | 11.0 (4.7) | 0.5 (0.1) | 2.41 |
| Location | Trees | Pneumatophores | |||||
|---|---|---|---|---|---|---|---|
| Transect | Distance from Fringe | Canopy Height | Frontal Area * | Density | Height | Diameter | Frontal Area |
| (m) | (m) | (m−1) | (# per m2) | (cm) | (cm) | (m−1) | |
| Along-shore | 0 | 0 | 0.26 | 40 | 1.5 (0.5) | 0.3 (0.2) | 0.12 |
| Along-shore | 5 | 0.97 | 0.26 | 164 | 1.7 (0.5) | 0.3 (0.2) | 0.49 |
| Along-shore | 15 | 0.48 | 0.26 | 296 | 1.1 (0.4) | 0.4 (0.2) | 1.18 |
| Along-shore | 25 | 0.41 | 0.26 | 308 | 1.1 (0.5) | 0.5 (0.2) | 1.54 |
| Along-shore | 35 | 0.36 | 0.26 | 296 | 0.6 (0.5) | 0.4 (0.2) | 1.18 |
| Along-shore | 45 | 0.38 | 0.26 | 252 | 1 (0.6) | 0.4 (0.2) | 1.00 |
| Along-shore | 55 | 0.365 | 0.26 | 248 | 6 (3.7) | 0.5 (0.2) | 1.24 |
| Along-shore | 65 | 0.34 | 0.26 | 356 | 8.7 (4.2) | 0.6 (0.1) | 2.14 |
| Along-shore | 75 | 0.37 | 0.26 | 196 | 10.8 (2.8) | 0.6 (0.2) | 1.18 |
| Along-shore | 85 | 0.29 | 0.25 | 244 | 7 (2.6) | 0.6 (0.1) | 1.46 |
| Along-shore | 95 | 0.23 | 0.26 | 240 | 5.1 (2.4) | 0.6 (0.1) | 1.44 |
| Along-shore | 105 | 0.26 | 0.26 | 216 | 10.3 (4.1) | 0.5 (0.2) | 1.08 |
| Along-shore | 115 | 0.27 | 0.25 | 172 | 4.3 (1.8) | 0.6 (0.2) | 1.03 |
| Along-shore | 125 | 0.22 | 0.26 | 172 | 5.3 (3.2) | 0.5 (0.2) | 0.86 |
| Along-shore | 135 | 0.29 | 0.26 | 264 | 6.9 (3.6) | 0.5 (0.1) | 1.32 |
| Along-shore | 145 | 0.26 | 0.25 | 248 | 7.4 (3.5) | 0.5 (0.2) | 1.24 |
| Along-shore | 155 | 0.23 | 0.25 | 364 | 6.3 (3.7) | 0.6 (0.2) | 2.18 |
| Along-shore | 165 | 0.23 | 0.25 | 292 | 6.1 (3.3) | 0.6 (0.2) | 1.75 |
| Along-shore | 175 | 0.22 | 0.25 | 252 | 6.2 (2.1) | 0.7 (0.2) | 1.76 |
| Along-shore | 185 | 0.2 | 0.25 | 172 | 6.3 (2.5) | 0.7 (0.1) | 1.20 |
| Along-shore | 195 | 0.2 | 0.26 | 200 | 13.6 (7.1) | 0.6 (0.2) | 1.20 |
| Along-shore | 205 | 0.32 | 0.26 | 212 | 5.4 (3.1) | 0.5 (0.1) | 1.06 |
| Along-shore | 215 | 0.3 | 0.26 | 224 | 11.2 (2.9) | 0.6 (0.1) | 1.34 |
| Along-shore | 225 | 0.24 | 0.26 | 192 | 7 (6.0) | 0.5 (0.1) | 0.96 |
| Along-shore | 200 | 0.27 | 0.26 | 276 | 11.7 (2.8) | 0.6 (0.2) | 1.66 |
| Along-shore | 190 | 0.29 | 0.26 | 324 | 3.7 (2.5) | 0.6 (0.2) | 1.94 |
| Cross-shore | 180 | 0.33 | 0.26 | 344 | 8.2 (6.3) | 0.6 (0.2) | 2.06 |
| Cross-shore | 170 | 0.52 | 0.26 | 192 | 11.7 (8.0) | 0.6 (0.1) | 1.15 |
| Cross-shore | 160 | 0.36 | 0.26 | 340 | 14.2 (7.1) | 0.6 (0.1) | 2.04 |
| Cross-shore | 150 | 0.35 | 0.26 | 356 | 8.5 (5.2) | 0.6 (0.1) | 2.14 |
| Cross-shore | 140 | 0.47 | 0.26 | 348 | 12.9 (7.3) | 0.6 (0.2) | 2.09 |
| Cross-shore | 130 | 0.54 | 0.26 | 396 | 10.2 (3.9) | 0.6 (0.2) | 2.38 |
| Cross-shore | 110 | 0.59 | 0.26 | 404 | 15.1 (7.0) | 0.6 (0.2) | 2.42 |
| Cross-shore | 100 | 0.63 | 0.26 | 536 | 12.7 (4.3) | 0.6 (0.1) | 3.22 |
| Cross-shore | 90 | 0.64 | 0.26 | 400 | 10.7 (2.5) | 0.5 (0.1) | 2.00 |
| Cross-shore | 80 | 0.71 | 0.26 | 552 | 8.1 (4.0) | 0.5 (0.1) | 2.76 |
| Cross-shore | 70 | 0.93 | 0.26 | 568 | 7.4 (2.7) | 0.5 (0.1) | 2.84 |
| Cross-shore | 60 | 0.55 | 0.26 | 404 | 8.8 (4.1) | 0.6 (0.2) | 2.42 |
| Cross-shore | 50 | 0.56 | 0.26 | 464 | 15.9 (8.6) | 0.6 (0.1) | 2.78 |
| Cross-shore | 40 | 0.42 | 0.26 | 408 | 7.6 (6.1) | 0.5 (0.1) | 2.04 |
| Cross-shore | 30 | 0.44 | 0.26 | 444 | 8.9 (4.2) | 0.6 (0.1) | 2.66 |
| Cross-shore | 20 | 0.72 | 0.25 | 308 | 10.5 (5.0) | 0.5 (0.1) | 1.54 |
| Cross-shore | 10 | 1.3 | 0.26 | 432 | 5.4 (2.8) | 0.5 (0.1) | 2.16 |
| Cross-shore | 0 | 0.97 | 0.26 | 268 | 12.8 (5.3) | 0.5 (0.1) | 1.34 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montgomery, J.M.; Bryan, K.R.; Horstman, E.M.; Mullarney, J.C. Attenuation of Tides and Surges by Mangroves: Contrasting Case Studies from New Zealand. Water 2018, 10, 1119. https://doi.org/10.3390/w10091119
Montgomery JM, Bryan KR, Horstman EM, Mullarney JC. Attenuation of Tides and Surges by Mangroves: Contrasting Case Studies from New Zealand. Water. 2018; 10(9):1119. https://doi.org/10.3390/w10091119
Chicago/Turabian StyleMontgomery, John M., Karin R. Bryan, Erik M. Horstman, and Julia C. Mullarney. 2018. "Attenuation of Tides and Surges by Mangroves: Contrasting Case Studies from New Zealand" Water 10, no. 9: 1119. https://doi.org/10.3390/w10091119
APA StyleMontgomery, J. M., Bryan, K. R., Horstman, E. M., & Mullarney, J. C. (2018). Attenuation of Tides and Surges by Mangroves: Contrasting Case Studies from New Zealand. Water, 10(9), 1119. https://doi.org/10.3390/w10091119