Abstract
In this study, a multilevel drought-induced resistance and resilience analysis (MDRRA) approach was developed to investigate the stability of vegetation in the Yellow River Basin (YRB). MDRRA was quantified by utilizing the Normalized Difference Vegetation Index (NDVI). It was applied to YRB to assess vegetation resistance and resilience to various levels of drought by utilizing precipitation and NDVI data from 2000 to 2019. The results reveal that vegetation resistance and resilience in YRB are affected by drought severity. Monthly and annual changes in SPI over the warm–temperate humid zone of the YRB show a decreasing trend, with rates of 0.001 per decade and 0.034 per decade, respectively; however, the other climatic subregions exhibit an increasing trend, with rates ranging from 0.002 per decade to 0.82 per decade. Over 77.56% of the downstream areas show increases in the annual SPI averages. Drought severity differs across subregions in the YRB. More severe drought events occur in its upper and middle reaches, while less severe ones happen in its lower reaches. As the drought severity increases, the arid and semiarid regions of the mesothermal zone exhibit a decrease in the resistance and resilience indices. MDRRA can help improve the stability and resilience of the ecosystem in the YRB.
1. Introduction
Under climate change, the Intergovernmental Panel on Climate Change (IPCC) indicated that more regions will be hit by droughts with higher frequency and severity in the future due to global warming [1]. Drought is one of the most devastating and universal extreme natural calamities, with a serious influence on water resources, ecosystems [2], the agricultural industry, and economic development [3]. For example, the increasing water stress projected for China could lead to a shift in the geographical distribution of agriculture and manufacturing [4]. Particularly, drought severity could affect the vegetation type and structure as well, which could have profound effects on vegetation growth, land degradation, and forest fires [5]. For example, increased droughts could seriously impact vegetation productivity, and persistent, severe droughts may even lead to the death of substantial vegetation [6,7]. Therefore, it is essential to conduct a comprehensive quantification of vegetation susceptibility to droughts with the purpose of predicting the consequences resulting from the influenced ecosystem functions and the associated ecosystem services under climate change.
Vegetation stability is usually assessed in terms of resistance and resilience. Resistance measures a drought’s impact on the ecosystem by gauging its ability to maintain its typical level during a drought [8]; resilience determines how quickly an ecosystem recovers to its normal state after experiencing a drought [9]. Currently, vegetation resistance and resilience at regional and global scales have been discussed in many studies through various methodologies.
For instance, the land-based ecosystem in India is comparatively delicate, with one-third of the watersheds and the majority of vegetation categories exhibiting lesser resistance to drought [10]. On the Indian Peninsula, half the catchment areas are not resistant to hydrological changes caused by climate warming [11]. In the Weihe River Basin of China, precipitation is one of the main factors influencing vegetation changes [12]. In southwestern China and the tropics, more drought occurrences have reduced forest resilience [13,14]. During the 2015–2016 El Niño event, the experimental plots in African tropical forests experienced the highest temperatures and driest conditions on record. Despite this, these extreme temperatures did not significantly reduce the carbon gains from tree growth or increase the carbon losses from tree mortality, demonstrating the resilience of African tropical forests to extreme weather anomalies [15]. Evidence based on scientific observation suggests that the sensitivity of vegetation growth to temperature has diminished in boreal regions in recent decades [16]. In terms of vegetation resilience, it is currently studied based on observational data from the sampling of tree annual rings, remote sensing, flux towers, etc. In Spain [17], the Tibetan Plateau [18], Central Asia [19], and even globally [20], researchers found that vegetation resilience was connected with vegetation types, and it often showed opposite patterns to resistance. Previous studies have concentrated on trees and used annual ring data from trees. During extreme drought events, trees allocate more photosynthetic products to roots, which further reduces radial growth. This suggests that trees’ annual ring data are not reliable indicators of primary productivity [21]. However, it has been limited to use data from remote sensing to evaluate the resilience of various vegetation types towards extreme drought. Data from remote sensing can effectively capture the spatial coherence of vegetation changes in productivity and growth [22]. Based on satellite vegetation indices, Huang et al. [23] quantified the temporal stability of biomes on a global scale and proved that broadleaf evergreen forests were more drought-tolerant. Zhang et al. [24] studied drought distribution and drought’s effect on vegetation productivity in China using PDSI and NDVI.
Previous studies have shown that vegetation’s response to drought varies spatially due to the heterogeneous severity of drought, leading to differing impacts on vegetation [25]. While the physiological mechanisms of vegetation’s response to drought are well established, the ecologically fragile Yellow River Basin (YRB) [26] lacks sufficient research on vegetation’s resistance and stability to drought. Therefore, an objective assessment of vegetation’s response to various drought disturbances is essential to enhance vegetation stability and determine its capacity to withstand drought.
The purpose of this paper was to develop a multilevel drought-induced resistance and resilience analysis (MDRRA) methodology to study vegetation stability in the Long Beach region. The MDRRA methodology identifies and categorizes drought events by calculating the Standardized Precipitation Index (SPI) and then assesses vegetation’s drought resistance and resilience based on NDVI. In this paper, the MDRRA method will be applied to assess the drought resistance and resilience of vegetation in the YRB zones using precipitation and NDVI data from 2000 to 2019. This objective entails the following: (i) the classification of the YRB based on different climatic characteristics [27]; (ii) an investigation of the spatiotemporal distribution of meteorological droughts within each climatic region; (iii) an investigation of the changing trends in drought for each climatic region; (iv) the identification of the various drought events in the basin; and (v) an assessment of ecosystem resistance and resilience to various levels of droughts.
2. Study Area and Data Collection
The YRB is located between longitude 96 and 119° E and latitude 32 and 42° N (Figure 1). Its source is the Tibetan Plateau, and it spans an area of 795,000 km2. The basin measures roughly 1900 km long from east to west and approximately 1100 km wide from north to south. Additionally, it displays notable distinctions in climatic elements across spatial extents and uneven temporal distribution throughout the annual and inter-seasonal periods. The YRB is situated in the mid-latitude zone, exhibiting significant variations in terrain and geomorphology. The YRB features varying climate zones, including arid, semi-arid, semi-humid, and humid regions. The upper reaches of the basin are mainly arid, while the central Shaanxi, Gansu, and Ningxia provinces fall under the semi-arid zone. The middle and lower reaches of the basin belong to the semi-humid zone. The spatial distribution of rainfall in the YRB varies significantly. There is a trend of greater rainfall in the south and less in the north, with a gradual decrease from the southeast to the northwest. The average annual precipitation in the YRB is approximately 476 mm. Since 2000, the area of the Yellow River Basin affected by meteorological drought has reached 528,000 square kilometers, accounting for 66.4% of the total basin area [28].
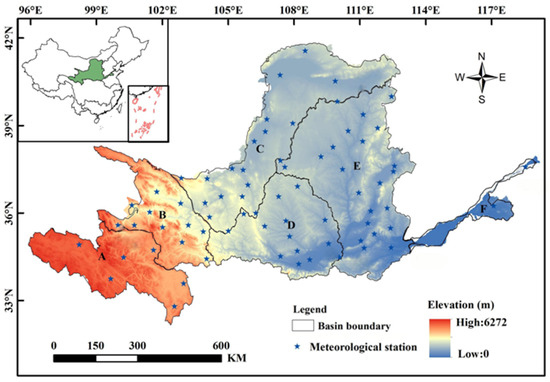
Figure 1.
Geographic location and zoning of the YRB. Zone A is the alpine belt semi-arid and semi-humid region; Zone B is cold–temperate belt semi-arid and semi-humid region; Zone C is the middle-temperate belt arid region; Zone D represents the warm–temperate belt semi-arid region; Zone E represents warm–temperate belt semi-arid and semi-humid region; and Zone F represents the warm–temperate zone humid region.
To systematically study drought conditions in different regions of the Yellow River Basin, this paper divides the entire basin into six sub-regions, considering the diverse climate across the basin [27]. The divisions are shown in Figure 1. Region A (cold semi-arid and semi-humid zone) covers the source area of the Yellow River; Region B is a transitional zone with a cold–temperate climate, classified as a semi-arid and semi-humid climate, serving as a transition between the plateau and the temperate climate zones; Region C is located on the Inner Mongolia Plateau, characterized as a temperate arid and semi-arid zone. The Ningxia–Mongolia section of the Yellow River flows through the Loess Plateau River valley basin and the river valley plain, situated in Region D, which has a warm–temperate semi-arid climate and is the largest tributary of the Yangtze River and the Weihe River Basin, characterized by a temperate continental and semi-arid climate. Region E (warm–temperate, semi-arid, and semi-humid zone) has a temperate continental climate, with semi-arid and semi-humid characteristics. Region F (warm–temperate humid zone) is located in the lower reaches of the Yangtze River, within the North China Plain, and it features a temperate monsoon climate and is classified as a humid zone.
The NDVI data used in this study were obtained from the SPOT/VEGETATION NDVI satellite remote sensing dataset, which is part of the China Long Time Series Vegetation Index (NDVI) dataset provided by the Center for Resource and Environmental Science and Data of the Chinese Academy of Sciences (https://www.resdc.cn/data.aspx?DATAID=254 (accessed on 10 April 2021)). The dataset includes China Monthly Vegetation Index (NDVI) spatial distribution data. In this paper, NDVI data from January 2000 to December 2019 were used, and pixels with an average NDVI of lower than 0.1 in non-vegetated areas, such as snow and water bodies, were excluded to ensure the accuracy and consistency of the study object. The referenced meteorological data for this research were monthly precipitation data for the YRB from January 2000 to December 2019, obtained from the National Meteorological Information Center website (http://data.cma.cn/en (accessed on 10 April 2024)).
3. Multilevel Drought-Induced Resistance and Resilience Analysis
In this study, MDRRA was developed based on the SPI and the Mann–Kendall trend test. In the first part, the SPI was calculated based on precipitation data from 2000 to 2019. The calculation was carried out for each sub-region of the Yellow River Basin (YRB) to systematically determine the period of drought. By applying the SPI method, each drought event was determined to occur in a specific time range in each region. The second part analyzed the SPI trends and classifications according to different drought severities. Finally, vegetation drought resistance and resilience in the YRB were measured using annual average NDVI data from 2000 to 2019. In the past, there has been a lack of adequate research on vegetation drought resistance and stability in the ecologically fragile YRB. To address this issue, MDRRA objectively assesses the response of vegetation to different levels of drought-disturbance characteristics. This assessment is essential for improving vegetation stability and determining its potential to withstand drought occurrence. Figure 2 illustrates the technical roadmap of this study.
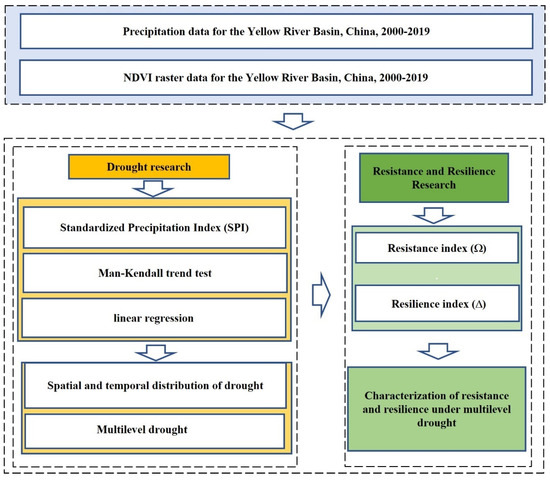
Figure 2.
Technology roadmap.
For the assessment of drought events, McKee et al. [29] first proposed the use of the SPI to describe drought conditions in Colorado. The SPI is a meteorological drought assessment indicator that characterizes the probability of precipitation occurring in a given period of time. The SPI is suitable for drought monitoring and assessment on time scales of more than one month [5]. Precipitation usually follows a skewed distribution. In precipitation analysis and drought monitoring and evaluation, the Γ distribution is often used to describe the changes in precipitation, and then the cumulative probability is standardized to obtain the SPI. The Γ distribution is a continuous probability distribution commonly used to model precipitation and other non-negative data. By using the Γ distribution, the cumulative frequency distribution of precipitation is transformed into a standard normal distribution, allowing for the comparison of precipitation conditions across different times and locations. The SPI describes multiple time scales, ranging from 1 to 48 months, allowing for a comprehensive analysis of temporal patterns. The 1-month-time-scale SPI is calculated by fitting a gamma probability distribution to the values of the precipitation series for the same month in different years. The cumulative frequency distribution of precipitation is transformed to a standard normal distribution using an equal probability transformation. Finally, the SPI for that month is calculated separately. The calculation of the SPI at various time scales necessitates cumulating rainfall values within a specified time frame. For example, to determine the annual SPI for a given year, all monthly rainfalls for that year are first summed to obtain the total annual rainfall. The annual total rainfall data for multiple years are then fitted to a gamma probability distribution, the cumulative probabilities are converted to a standard normal distribution, and the SPI for that year is determined. The steps for calculating the SPI are as follows: Let X represent the monthly precipitation time series, and let denote the accumulated monthly precipitation series over a time scale, w, where w = 1, 3, 6, …, and represents the accumulated monthly precipitation series for a specific month, with mon = 1, 2, 3, …, 12. The formula for calculating the SPI can be expressed as follows:
where F denotes the Gamma distribution function, and represents the inverse of the standard normal distribution function.
The SPI has the characteristic of characterizing moisture gains and losses on long time scales, and the classification classes of the SPI are shown in Table 1.

Table 1.
SPI drought-class classification.
To comprehend the spatial and chronological features of drought in the YRB, this study utilized the Mann–Kendall trend test, along with the linear regression approach for computation.
The Mann–Kendall trend test is a widely used test for hydrological trend analysis. It can be applied to complex sequences and is unsuitable for identifying trend curves [30]. The method is employed to determine the trend status and significance of various time series. The calculation formula is provided below for reference:
For a time series, x, with sample size n, at moment i (i = 1, 2, …, n − 1), there are
If all values in the sequence increase compared to the previous one, then the sequence is considered to have an upward trend. If not, the trend is considered a downward one. The test statistic of this trend is as follows:
The statistic U is defined as obeying a positive distribution, and the significant level α in this paper is taken to be 0.05, at which time |U| < Uα/2 = 1.96, the change in the series is considered to be non-significant, or else the trend of the time series is judged to be significant. is the Mann–Kendall statistic, used to measure the strength and direction of the trend in time series data.
Linear regression analysis on raster sequences uses the image element scale [31]. This analysis represents the trend of a specific pixel over time, with the NDVI serving as an example. The calculation method is outlined below:
where S is the slope of tendency. When , it means that a pixel shows an increasing trend over time, and vice versa shows a decreasing trend; t is the year, and is the NDVI value of the i year on a pixel.
Resistance is the ability of vegetation to resist external disturbances. Resilience refers to the ability of vegetation to restore itself to its pre-disturbance state after disturbance and perturbation. The resistance index () and the resilience index (Δ) are calculated as follows [32]:
where represents normal conditions, which means the NDVI without drought (SPI > −0.5), represents the NDVI value during a drought, and represents the multi-year NDVI mean of the time series. The resistance and resilience classes [25] are delineated in Table 2.
Table 2.
Resistance and resilience levels’ division.
The level of the resistance index and the resilience index represents the level of resistance and resilience of the vegetation. A higher resistance index signifies increased resistance, while a higher resilience index signifies greater recuperative ability and also suggests more stable and, thus, more resilient vegetation. At the same time, the greater the resistance, the less likely the NDVI is to change during a disturbance, and the more stable the vegetation. The higher the resilience, the quicker the recovery of NDVI after disturbance, resulting in faster vegetation recovery to its disturbed state.
4. Results
4.1. Spatial and Temporal Distribution of Drought
To provide a more precise description of the drought situation in the six sub-regions, we present an example of SPI1 analysis. This allows for the study of changes in meteorological drought characteristics within each sub-region. As Figure 3 shows, the minimum SPI value in Region A occurred in February 2015, with a value of −2.401. The minimum SPI values in Regions B and C both occurred in March 2013, with values of −2.653 and −2.304, respectively. The minimum SPI value in Region D occurred in December 2008, with a value of −2.235. The minimum SPI value in Region E occurred in May 2001, with a value of −2.313. The minimum SPI value in Region F occurred in July 2015, with a value of −2.381. We can see that the increasing and decreasing trend of SPI1 was not the same in each sub-district. Zone A through E indicated an increasing trend, suggesting an overall wetting of the watershed. Zone A had the highest growth rate, which was 0.037 per decade, while Zone D had the lowest growth rate of 0.008 per decade. The growth rates in Zones B, C, and E were 0.027/10a, 0.023/10a, and 0.011/10a, respectively. Zone F exhibited a declining trend at a rate of 0.008 per decade.
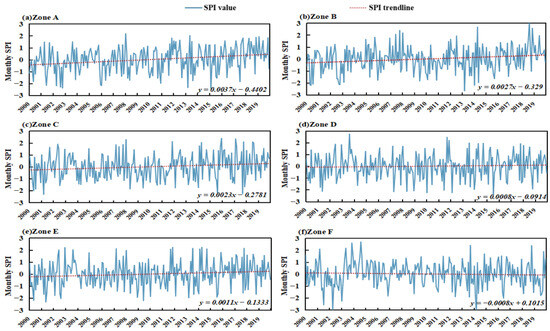
Figure 3.
Trend of monthly-scale SPI1 in the YRB by subregion, 2000–2019.
We first aggregated the monthly precipitation data for each year in each region to obtain the total annual precipitation. Then, we standardized these annual total precipitation values using a fitted gamma probability distribution to calculate the annual SPI values for each region, specifically SPI12. The outcomes for each subregion are illustrated in Figure 4. From the results, it can be seen that there was an increasing trend from A to E in the same way as the trend in the monthly scale. In Zone C, the growth rate was the highest at 0.82/10a, followed by Zone E, which had the slowest rate at 0.245/10a. Zone F displayed a decreasing trend with a rate of 0.671/10a. The SPI12 values in the YRB’s regions ranged from −3 to 2.5, with occurrences of extreme drought events. Specifically, both A and B experienced an extreme drought year in 2002. In contrast, the midstream Zones of D and E exhibited smoother characteristics, with SPI values fluctuating around 0.
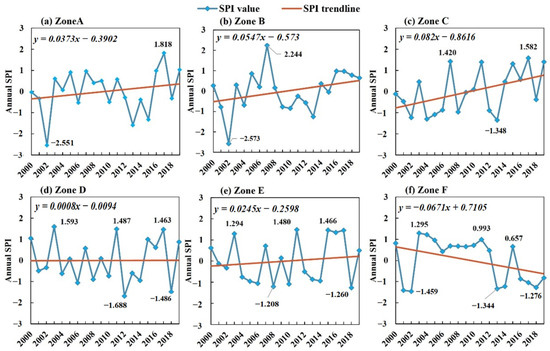
Figure 4.
Trends of annual-scale SPI12 in the YRB by subregion, 2000–2019.
The study utilized the Mann–Kendall trend test to ascertain monthly and yearly SPI trends in each basin. Table 3 details the results for each basin, whereby the monthly and yearly SPI of Zone C exhibited a significant upward trend, having passed the 95% confidence test. The monthly and yearly SPI for Zones A and B exhibited a significant upward trend and a non-significant upward trend, respectively. Notably, the SPI trend of monthly and yearly observations was consistent with the significance of the yearly SPI. Conversely, the monthly and yearly SPI for Zones D, E, and F did not pass the confidence test, suggesting no discernible trend. Interestingly, the SPI for Zones D, E, and F also did not pass the confidence test, and the monthly SPI trend mirrored that of the annual SPI significance trend.
Table 3.
Mann–Kendall trend test for monthly and annual SPI in the YRB.
A trend analysis of SPI12 in the YRB utilized one-way linear regression. As depicted in Figure 5, there was great spatial heterogeneity in the distribution of SPI12 trends across the YRB. Among them, approximately 77.56% of the regional SPI values exhibited an upward trend, indicating increased precipitation, primarily concentrated in the southeastern and central regions of the YRB. Zone B had the greatest expansion, covering 96.69% of the total watershed area, while Zone D followed with a growth of 96.63% of the total watershed area. This was followed by Zone A (96.62%), Zone B (96.54%), Zone E (94.16%), and Zone F (71.9%). In the YRB, 22.44% of the area exhibited a decreasing trend in SPI-12 values, indicating a trend toward aridification, primarily concentrated in the southeastern region of the basin. Among these, Zone F had the largest decrease in area, accounting for 28.10% of the total basin area.
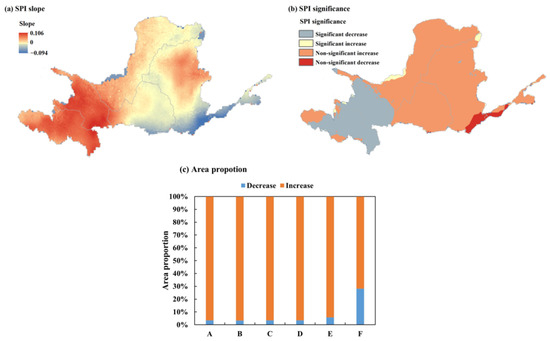
Figure 5.
SPI12 (a) slope, (b) significance, and (c) area proportion in the YRB by subregion, 2000–2019.
4.2. Selection of Years with Different Degrees of Drought
The years with varying degrees of drought in each subregion are displayed in Table 4, based on the annually calculated SPI12 values. The SPI values were interpolated spatially using the inverse distance weighting method [33]. The resulting percentage of areas with corresponding degrees of aridity is displayed in Figure 6. During the period of 2000–2019, there was a prevalence of light and moderate drought years. To choose the representative years and prepare for the next phase of resistance and resilience calculation, we assessed the SPI12 value and the percentage of areas affected by light and moderate droughts. The remaining years with severe and extreme droughts were determined based on SPI12 value.
Table 4.
Distribution of drought years by subregion in the YRB.
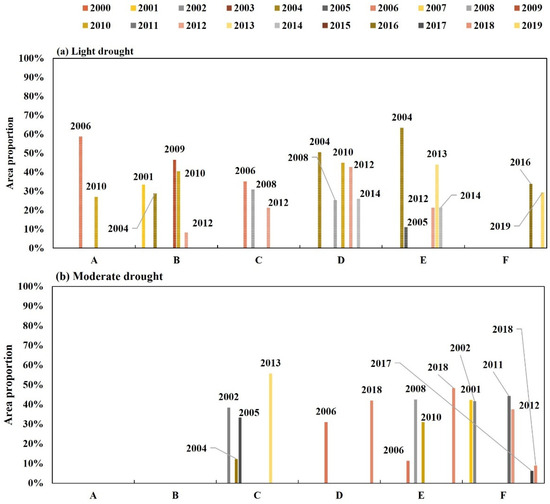
Figure 6.
Percentage of area in the YRB with (a) light and (b) moderate drought in each subregion, 2000–2019.
The droughts that occurred in Zones A and B of the YRB in 2002 proved to be the most severe within the 2000–2019 timeframe. The lowest SPI12 values for Zones A and B were recorded in 2002 at −2.55 and −2.57, respectively. In the remaining zones, there were no years with extreme drought. The years of severe drought in Zone A and Zone D were 2013 and 2012, respectively, with corresponding SPI12 values of −1.59 and −1.69. No severe drought years occurred in the remaining subzones. The years of light drought in Zone A were 2006 and 2010, with SPI12 values of −0.53 and −0.50, respectively. In those years, the proportion of the light drought Zone to the total Zone was 59% and 27%, respectively. Therefore, 2006 was selected as the representative year of light drought in Zone A. The SPI12 values for Zone B during light drought years ranged from −0.57 to −0.84. The lowest SPI12 value, recorded in 2010, was −0.84, while −0.78 was observed in 2001. The area experiencing mild drought reached a peak of 46% in 2009, with a subsequent decrease to 40% in 2010. Based on an analysis of SPI12 values and the percentage of affected land, 2010 was determined to be a representative year for mild drought in Zone B. The years of mild drought in Zone C were 2006, 2008, and 2012, with SPI values of −0.87, −0.97, and −0.90, respectively. The light drought area accounted for 35%, 31%, and 21% of the total area in the aforementioned years. Therefore, 2008 was chosen as the year that best represented the occurrence of light drought in Zone C. The SPI12 values for moderate drought years in Zone C ranged from −1.09 to −1.34. The highest SPI12 value was recorded in 2013, with −1.30 in 2004 being the second highest. In terms of mild drought, the highest-area share was observed in 2013 (56%) followed by 38% in 2002. Given the SPI12 values and the area share, 2013 was identified as the representative year for moderate drought in Zone C. The years with light drought conditions in Area D were identified based on SPI values. Specifically, SPI12 values of −0.62 (2004), −0.89 (2008), −0.73 (2010), −0.60 (2013), and −0.95 (2014) corresponded to 50%, 25%, 45%, 43%, and 26% of the area, respectively. 2010 was ultimately selected as the representative year for light drought. The SPI12 value and area share of moderate drought were greatest in Zone D in 2018. Consequently, 2018 was determined to be the year that best represented moderate drought in Area D. The year 2005 had the highest SPI12 value of −0.96, while 2014 and 2013 had SPI values of −0.34 and −0.87, respectively. The areas affected by light drought in these years were 11%, 21%, and 44%, respectively. Thus, 2013 was chosen as the representative year for light drought in Zone E. In 2018, Zone E exhibited the largest SPI12 and area percentage in moderate drought. Therefore, 2018 served as the representative year for moderate drought in Zone E. In 2016, Zone F exhibited the highest SPI12 value and the largest percentage of the Zone affected by light drought, with values of −0.87 and 34%, respectively. Therefore, this year was selected as the representative year for light drought in Zone F. Similarly, the year 2002 served as the exemplar for moderate drought in Zone F, with a SPI12 value of −1.46, the highest among moderate droughts, and a coverage rate of 42%.
In summary, Zone A had representative years of light (2006), moderate (2015), severe (2013), and extreme (2002) drought. Zone B had representative years of light (2010), moderate (2013), and extreme (2002) drought but no representative year of severe drought. Zone C had representative years of light (2008) and moderate (2013) drought but no representative years of severe or extreme drought. In Zone D, representative years for light, moderate, and severe drought were 2010, 2018, and 2012, respectively. No extreme drought-representative years were observed. In Zone E, the representative years for drought were 2013 and 2018, respectively, with no severe or extreme drought representative years. Zone F experienced droughts in 2016 and 2002, with no severe or extreme representative years.
4.3. Correlation of SPI and NDVI across Regions
Figure 7 illustrates the spatial distribution of the correlation between the NDVI and the multi-scale SPI, where SPI1 and SPI12 represent the correlations at 1-month and 12-month time scales, respectively. The results reveal significant spatial heterogeneity within the study area. The spatial distribution of the SPI and NDVI correlations at both 1-month and 12-month time scales was relatively consistent, with total correlation coefficients ranging from −0.84 to 0.97. In Region A, SPI1 and SPI12 exhibit a generally positive correlation with the NDVI, though negative correlations are primarily concentrated in the central and southwestern parts. The correlation coefficients between the SPI1 and NDVI range from −0.84 to 0.85, while, for SPI12, they range from −0.74 to 0.94, with positive-correlation areas constituting 56% and 76% of Region A, respectively. In Region B, SPI12 has the highest positive correlation coefficient of 0.97 across the Yellow River Basin, with the positive correlation areas of SPI1 and SPI12 covering 82% and 86% of Region B, respectively. In Region C, SPI1 and SPI12 show predominantly positive correlations with the NDVI, with positive correlation areas covering 96% and 97% of Region C, respectively. The correlation coefficients range from −0.73 to 0.80 for SPI1 and −0.70 to 0.96 for SPI12. In Region D, the southern part showed negative correlations between SPI1, SPI12, and the NDVI, but overall, SPI1 and SPI12 exhibited positive correlations with the NDVI, with correlation coefficients ranging from −0.73 to 0.78 (SPI1) and −0.81 to 0.89 (SPI12) and positive correlation areas making up 86% and 88% of Region D, respectively. In Region E, SPI1 and SPI12 generally showed positive correlations with the NDVI, with coefficients ranging from −0.71 to 0.76 for SPI1 and −0.78 to 0.93 for SPI12 and positive correlation areas constituting 95% of Region E for both SPI1 and SPI12. In Region F, SPI1 and SPI12 also exhibited overall positive correlations with the NDVI, with coefficients ranging from −0.73 to 0.70 for SPI1 and −0.63 to 0.91 for SPI12 and positive correlation areas covering 69% and 76% of Region F, respectively.
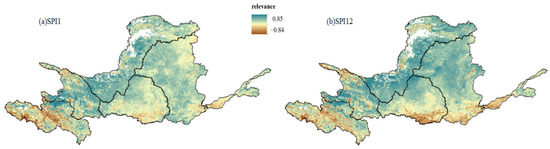
Figure 7.
Correlation of subregions with NDVI for (a) SPI1 and (b) SPI12 in the YRB, 2000–2019.
4.4. Resistance and Resilience under Different Drought Levels
Ecosystem resistance and resilience are two essential characteristics that reflect the steady state of ecosystems under climate change. Resistance in ecosystems is associated with characteristics such as vegetation types and attributes connected to drought stress, including the degree of drought, duration, and post-drought environmental conditions [34]. This paper classifies different levels of drought years based on the annual SPI index. It calculates the resistance and resilience of vegetation to varying degrees of drought in each subregion of the YRB, with results presented in Figure 7, Figure 8, Figure 9 and Figure 10.
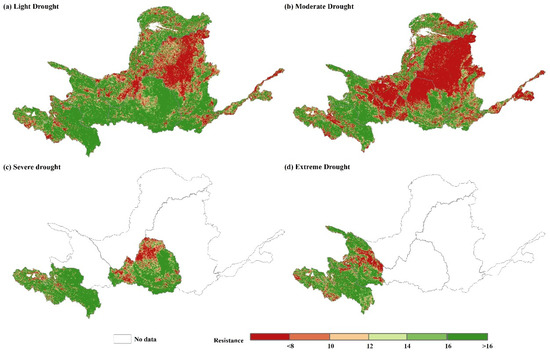
Figure 8.
Spatial distribution of resistance under (a) light drought, (b) moderate drought, (c) severe drought, and (d) extreme drought in the YRB, 2000–2019.
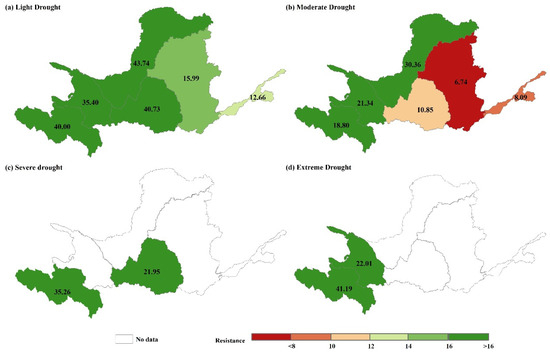
Figure 9.
Mean values of resistance under (a) light drought, (b) moderate drought, (c) severe drought, and (d) extreme drought in the YRB, 2000–2019.
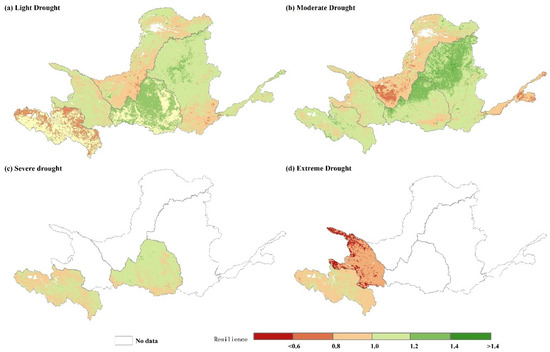
Figure 10.
Spatial distribution of resilience under (a) light drought, (b) moderate drought, (c) severe drought, and (d) extreme drought in the YRB, 2000–2019.
Figure 8 and Figure 9 display the spatial distribution and average resistance values to various degrees of drought in each subregion of the YRB between 2000 and 2019. The spatial distribution of resistance in the basin is heterogeneous, with weak resistance to light and moderate drought in the central and downstream areas and higher resistance in the southwestern and southeastern parts. Area A experienced four degrees of drought, Area B experienced light, moderate, and extreme drought, and Area D experienced light, moderate, and severe drought, respectively. Figure 6c,d indicate that the southeastern part of Zone A has high resistance to severe and extreme drought, while the central part of the region has Zones with low resistance to severe drought. Additionally, there are Zones with low resistance to extreme drought in the southeastern part of Zone A. Resistance to severe drought was lower in the northwestern part of Zone D and in the central part of Zone B for extreme drought. The mean resistance to varying levels of drought was higher in Zone A, ranging from 18.80 to 41.19, with the greatest resistance shown towards extreme drought and the lowest resistance displayed towards moderate drought. Resistance to light, moderate, and severe drought was higher in Zone B compared to Zone A, albeit marginally lower. The mean values of resistance ranged from 21.34 to 35.40, with the maximum resistance to light drought and the minimum resistance to extreme drought. Resistance to drought in Zone C decreased as the drought severity increased. The mean resistance value decreased from 43.74 under a light drought to 30.36 under a moderate drought. The resistance to varying degrees of drought in Zone D significantly varied compared to other subzones. The resistance to light and severe droughts was higher, at 40.73 and 21.95, respectively, while resistance to moderate drought was weaker, only showing a medium level of resistance at 10.85. The resistance variability in Zones E and F resembled that of Zone C. Zone E demonstrated a shift from high resistance to light drought (15.99) to low resistance to moderate drought (6.74), whereas Zone F demonstrated a change from medium resistance to light drought (12.66) to low resistance to moderate drought (8.09).
Figure 10 and Figure 11 depict the spatial distribution and mean values of resilience to various levels of drought in the YRB for each subregion between 2000 and 2019. The resilience distribution is spatially heterogeneous, with higher resilience observed in areas of Zone D post a light drought. In contrast, lower resilience was noted more in the eastern part of Zone A and the southern part of Zone E. Areas with higher resilience following a moderate drought are predominantly concentrated in the northern regions of Zones C and D, whereas the southwestern part of Zone C and the northern area of Zone F exhibit lower resilience. The spatial distribution of resilience displays comparable heterogeneity. After a severe drought, resilience is lower in Zone A, with the exception of the central region, and in the southern part of Zone D. In the aftermath of an even more severe drought, resilience is diminished in the northern and northeastern parts of Zone B. In terms of mean values, the resilience of the six subzones varied between medium and low (0.88–1.16) following exposure to various levels of drought. Similarly, Zone A demonstrated medium resilience after experiencing different degrees of drought, with the lowest resilience recorded at 0.97 during the most severe drought. Furthermore, resilience values during light, medium, and severe drought were 1.02, 1.06, and 1.00, respectively. Zone B exhibited equivalent resilience after experiencing light and moderate drought, maintaining a medium-grade resilience (1.03). However, after an extreme drought, the resilience decreased to 1.00. Zone C encountered a slight reduction in resilience when subjected to increasing drought severity, with a decrease from 0.99 to 0.94, while maintaining medium-grade resilience. Zone D experienced less of a change in resilience after light, moderate, and severe droughts, with resilience increasing to 1.11 and then decreasing to 1.05 after a moderate drought. In Zone E, resilience increased from 1.07 to 1.16 with increasing drought severity, while the opposite occurred in Zone F, with resilience decreasing from 1.06 in the moderate rating to 0.88 in the low rating.
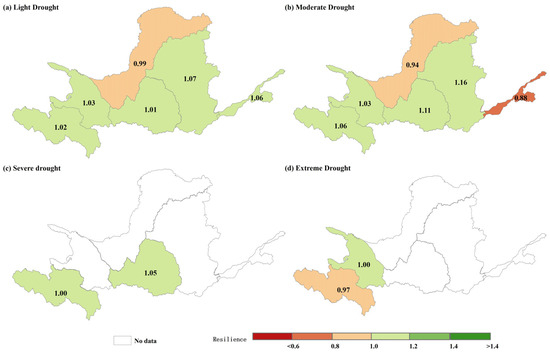
Figure 11.
Mean values of resilience under (a) light drought, (b) moderate drought, (c) severe drought, and (d) extreme drought in the YRB, 2000–2019.
5. Discussion
According to the study of Zhou et al. [35], based on the SPEI index, extreme droughts, severe droughts, and moderate droughts are mostly distributed in the northwest region of the YRB, showing a decreasing trend from the northwest region to the southeast region. This is consistent with the results calculated in this paper. The results calculated in this paper show that the years of a mild drought in each region of the Yellow River Basin are the highest, followed by moderate drought, and the years of a severe drought and extreme drought are significantly less than the other two types of droughts. Studies have found that each region of the Yellow River Basin is dominated by mild droughts, with a frequency of 14.91% to 16.23%. The frequency of moderate droughts is the second highest. In contrast, the incidence of the other two droughts by severity is relatively low, especially extreme droughts, with an incidence of about 1%. The results are consistent with the results of this study [36]. Extreme climatic events such as droughts reduce vegetation’s primary productivity by reducing the water availability and net ecosystem carbon dioxide exchange, and the resulting drought stress can further reduce plant viability [37]. However, this study has revealed a positive correlation between the degree of drought and the drought resistance of vegetation in individual sub-areas. This suggests that an increase in the degree of drought led to an increase in the drought resistance of vegetation. The resistance of plants to drought could be attributed to a major factor, namely their ability to adapt to poor hydrothermal conditions [38]. Liu et al. [32] also showed that, despite the occurrence of drought, the gap between the atmospheric temperature and the optimum temperature of vegetation prevents drought from negatively affecting ecosystem photosynthesis and that the effects of drought on plants are complex and involve a certain degree of uncertainty.
Since resilience is a measure of the ability of an ecosystem to return to its normal state, the ability of an ecosystem to recover from disturbance to favorable conditions for vegetation growth becomes a determinant of the spatial pattern of vegetation resilience. In areas with higher precipitation, the probability of soil moisture being replenished in a shorter period of time is higher, and ecosystems have a higher capacity for vegetation resilience. For hot and dry areas, high temperatures inhibit vegetation growth, resulting in lower vegetation resilience in these areas. It is possible that, despite the worsening drought, its resilience will not diminish. Herbaceous plants in semi-arid or arid regions have shallow root systems that restrict their access to stable groundwater sources. This makes them more susceptible to precipitation and even minor rewetting events after droughts. Wang et al. [39] found that drought resilience efficiency in the inland northwest coastal areas of China is relatively high. The same results were observed in arid and semi-arid regions of Central Asia [40].
The level of resilience was not consistently high in regions with high resistance, owing to the unstable correlation between stability components. Resistance may be either positively or negatively linked to resilience in this context [41]. Under normal circumstances, the ecosystem’s function fluctuates within a certain range. The size of the fluctuation reflects the ecosystem’s changes over time. The larger the fluctuation, the smaller the temporal stability. The ecosystem responds to disturbance and the function declines, resulting in deviation. When subjected to the same disturbance, the larger the deviation value, the weaker the ability to resist external intrusion. Conversely, the greater the resistance, the less susceptible the ecosystem is. If the recovery rate is slower and the ecosystem is less resilient, it takes longer to return to the original state after experiencing the same type of disturbance. According to ecology and evolutionary theory, ecosystems can gradually adjust to unfavorable conditions [42]. The ability to adapt renders species, communities, habitats, or ecosystems more resilient while impeding their resistance. Such adaptability increases the likelihood of survival and relative stability under future climatic conditions.
6. Conclusions
The conclusions drawn from this study are as follows.
In the YRB, the SPI1 and SPI12 in Zone F showed a non-significant decreasing trend, whereas other climatic zones exhibited an increasing trend in both SPI1 and SPI12. Additionally, the increasing trend in SPI12 covered 77.56% of the total area of the YRB.
Different parts of the YRB experienced varying levels of drought. The upper and middle climatic areas underwent more severe drought, with the year 2002 having extreme cases. The lower portions of the river had milder droughts, including mild and moderate drought years.
In all study areas, SPI1 and SPI12 showed a generally positive correlation with the NDVI. The C zone had the largest proportion of positive correlation, with the area where SPI1 and SPI12 were positively correlated with the NDVI accounting for 96% and 97% of the total area, respectively. In contrast, the A zone had the smallest proportion of positive correlation, but the area where SPI1 and SPI12 were positively correlated with the NDVI still exceeded 50% of the total area.
Increased drought intensity leads to decreased resistance and resilience in the arid and semi-arid zones of the mesothermal regions and the humid zones of the warm–temperate regions.
This study focused solely on the impact of meteorological droughts on vegetation, and it did not analyze soil droughts. During periods of low precipitation, plants are likely to grow normally if the soil is not water-stressed. Further research is needed to investigate the impact of soil droughts on vegetation growth. Additionally, the study did not consider the delayed consequences of long-term droughts, such as drought-induced tree mortality and wildfires, as well as the delayed and cumulative effects of droughts on vegetation. Further research is also needed on the effects of soil, topography, geology, and socio-economic development on droughts. Although this study used NDVI information to assess vegetation resistance and resilience, this method has limitations without field-reference data. Relying solely on NDVI data may not fully reflect the actual impact of droughts on vegetation, as the NDVI only provides remote sensing information about vegetation conditions, and it cannot directly measure physiological states or groundwater conditions. Therefore, combining field-observation data with NDVI remote sensing information will help more comprehensively and accurately assess vegetation responses to drought.
In the future, this type of research can be further improved by incorporating the concept of “recovery” to gain a comprehensive understanding of the concepts related to the vegetation NDVI. Recovery refers to the ability of vegetation to return to a normal state after experiencing extreme weather events such as droughts. Future studies could assess recovery by monitoring the rate and magnitude of NDVI value changes after the end of a drought event. For example, analyzing the time required for NDVI values to return to normal levels and evaluating the growth magnitude of the NDVI during the recovery period can provide a detailed understanding of recovery-time differences and degrees among different regions and vegetation types. Additionally, studying key factors influencing recovery, such as soil types, water availability, vegetation types, and human interference, can help identify which factors play a crucial role in promoting or hindering vegetation recovery.
Author Contributions
J.F.: data curation, formal analysis, and writing—original draft. W.Z., and F.X.: data curation and formal analysis. X.Z., and W.D.: conceptualization and methodology. C.W., S.W., Y.Z., and D.W.: investigation, validation, and supervision. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (grant number 52209013), the Department of Education of Hebei Province (grant number ZD2022085), and Fundamental Research Funds for the Central Universities (Grant NO. 300102293508).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets analyzed during the current study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sun, Y. Impact of human activities on climate system: An interpretation of Chapter III of WGI report of IPCC AR6. Transac. Atmos. Sci. 2021, 44, 654–657. [Google Scholar]
- Fang, L.; Wang, L.; Chen, W.; Sun, J.; Cao, Q.; Wang, S.; Wang, L. Identifying the impacts of natural and human factors on ecosystem service in the Yangtze and Yellow River Basins. J. Clean. Prod. 2021, 314, 127995. [Google Scholar] [CrossRef]
- Fan, J.; Wei, S.; Liu, D.; Qin, T.; Xu, F.; Wu, C.; Liu, G.; Cheng, Y. Impact of ENSO events on meteorological drought in the Weihe River basin, China. Front. Earth Sci. 2023, 11, 1093632. [Google Scholar] [CrossRef]
- Liu, M.; Zhou, X.; Huang, G.; Li, Y. The increasing water stress projected for China could shift the agriculture and manufacturing industry geographically. Commun. Earth Environ. 2024, 5, 396. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.; Chen, Y.; Liu, M.; Gao, L. Flood/drought event identification using an effective indicator based on the correlations between multiple time scales of the Standardized Precipitation Index and river discharge. Theor. Appl. Climatol. 2017, 128, 159–168. [Google Scholar] [CrossRef]
- Li, Y.; Dong, G.; Xue, H.; Zheng, Y.; Lian, Y. Effects of multi time scale meteorological drought on vegetation in the Yellow River Basin from 1982 to 2020. J. Soil. Water Conserv. 2024, 38, 187–196. [Google Scholar]
- Fan, J.; Wei, S.; Liu, G.; Zhou, X.; Li, Y.; Wu, C.; Xu, F. Response Time of Vegetation to Drought in Weihe River Basin, China. Atmosphere 2023, 14, 938. [Google Scholar] [CrossRef]
- Van Ruijven, J.; Berendse, F. Diversity enhances community recovery, but not resistance, after drought. J. Ecol. 2010, 98, 81–86. [Google Scholar] [CrossRef]
- Ivits, E.; Horion, S.; Erhard, M.; Fensholt, R. Assessing European ecosystem stability to drought in the vegetation growing season. Glob. Chang. Biol. 2016, 25, 1131–1143. [Google Scholar] [CrossRef]
- Jha, S.; Das, J.; Sharma, A.; Hazra, B.; Goyal, M.K. Probabilistic evaluation of vegetation drought likelihood and its implications to resilience across India. Glob. Planet. Chang. 2019, 176, 23–35. [Google Scholar] [CrossRef]
- Sinha, J.; Sharma, A.; Khan, M.; Goyal, M.K. Assessment of the impacts of climatic variability and anthropogenic stress on hydrologic resilience to warming shifts in Peninsular India. Sci. Rep. 2018, 8, 13833. [Google Scholar] [CrossRef]
- Fan, J.; Wang, D.; Zhao, Y.; Zhou, X.; Cheng, Y.; Xu, F.; Wei, S.; Liu, H. Spatiotemporal geographically weighted regression analysis for runoff variations in the Weihe River Basin. J. Environ. Manag. 2024, 366, 121908. [Google Scholar] [CrossRef] [PubMed]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef]
- Verbesselt, J.; Umlauf, N.; Hirota, M.; Holmgren, M.; Van Nes, E.H.; Herold, M.; Zeileis, A.; Scheffer, M. Remotely sensed resilience of tropical forests. Nat. Clim. Chang. 2016, 6, 1028–1031. [Google Scholar] [CrossRef]
- Bennett, A.C.; Dargie, G.C.; Cuni-Sanchez, A.; Tshibamba Mukendi, J.; Hubau, W.; Mukinzi, J.M.; Phillips, O.L.; Malhi, Y.; Sullivan, M.J.P.; Cooper, D.L.M. Resistance of African tropical forests to an extreme climate anomaly. Proc. Natl. Acad. Sci. USA 2021, 118, e2003169118. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Nan, H.; Huntingford, C.; Ciais, P.; Friedlingstein, P.; Sitch, S.; Peng, S.; Ahlström, A.; Canadell, J.G.; Cong, N. Evidence for a weakening relationship between interannual temperature variability and northern vegetation activity. Nat. Commun. 2014, 5, 5018. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J.; Vicente-Serrano, S.M.; Sánchez-Salguero, R.; Gutiérrez, E.; de Luis, M.; Sangüesa-Barreda, G.; Novak, K.; Rozas, V.; Tíscar, P.A.; et al. Forest resilience to drought varies across biomes. Glob. Chang. Biol. 2018, 24, 2143–2158. [Google Scholar] [CrossRef]
- Fang, O.; Zhang, Q. Tree resilience to drought increases in the Tibetan Plateau. Glob. Chang. Biol. 2018, 25, 245–253. [Google Scholar] [CrossRef]
- Fan, X.; Hao, X.; Hao, H.; Zhang, J.; Li, Y. Comprehensive Assessment Indicator of Ecosystem Resilience in Central Asia. Water 2019, 13, 124. [Google Scholar] [CrossRef]
- Li, X.; Piao, S.; Wang, K.; Wang, X.; Wang, T.; Ciais, P.; Chen, A.; Lian, X.; Peng, S.; Peñuelas, J. Temporal trade-off between gymnosperm resistance and resilience increases forest sensitivity to extreme drought. Nat. Ecol. Evol. 2020, 4, 1075–1083. [Google Scholar] [CrossRef]
- Kannenberg, S.A.; Novick, K.A.; Alexander, M.R.; Maxwell, J.T.; Moore, D.J.P.; Phillips, R.P.; Anderegg, W.R.L. Linking drought legacy effects across scales: From leaves to tree rings to ecosystems. Glob. Chang. Biol. 2019, 25, 2978–2992. [Google Scholar] [CrossRef] [PubMed]
- Keyserlingk, J.V.; Hoop, M.D.; Mayor, A.G.; Dekker, S.C.; Rietkerk, M.; Foerster, S. Resilience of vegetation to drought: Studying the effect of grazing in a Mediterranean rangeland using satellite time series. Remote Sens. Environ. 2021, 255, 112270. [Google Scholar] [CrossRef]
- Huang, K.; Xia, J. High ecosystem stability of evergreen broadleaf forests under severe droughts. Glob. Chang. Biol. 2019, 25, 3494–3503. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xiao, J.; Zhou, Y.; Zheng, Y.; Li, J.; Xiao, H. Drought events and their effects on vegetation productivity in China. Ecosphere 2016, 7, e01591. [Google Scholar] [CrossRef]
- Shi, X.; Chen, F.; Ding, H.; Li, Y.; Shi, M. Quantifying Vegetation Stability under Drought in the Middle Reaches of Yellow River Basin, China. Forests 2022, 13, 1138. [Google Scholar] [CrossRef]
- Li, J.; Sun, W.; Li, M.; Meng, L. Coupling coordination degree of production, living and ecological spaces and its influencing factors in the Yellow River Basin. J. Clean. Prod. 2021, 298, 126803. [Google Scholar] [CrossRef]
- Li, Y. Basin Drought Assessment—Propagation—Driving—Prediction under Climate and Land Use Change. Ph.D. Thesis, Xi’an university of Technology, Xian, China, 2018. [Google Scholar]
- Fang, H. Research on the Evolution Law of Drought in the Yellow River Basin at Multi-Temporal and Spatial Scales. Ph.D. Thesis, Hebei University of Engineering, Handan, China, 2014. [Google Scholar]
- McKee, T.B.; Doesken, N.J.; Kleist, J. The Relationship of Drought Frequency and Duration to Time Scales. In Proceedings of the 8th Conference on Applied Climatology, Anaheim, CA, USA, 17–22 January 1993; Volume 17, pp. 179–183. [Google Scholar]
- Agbo, E.P.; Nkajoe, U.; Edet, C.O. Comparison of Mann–Kendall and Şen’s innovative trend method for climatic parameters over Nigeria’s climatic zones. Clim. Dyn. 2023, 60, 3385–3401. [Google Scholar] [CrossRef]
- Yuan, L.; Jiang, W.; Shen, W.; Liu, Y.; Wang, W.; Tao, L.; Zheng, H.; Liu, X. The spatiotemporal variations of vegetation cover in the Yellow River Basin from 2000 to 2010. Acta Ecol. Sin. 2013, 33, 7798–7806. [Google Scholar]
- Liu, Y.; Li, X.; Wang, X.; Wang, S.; Qian, Z. Response of vegetation stability to extreme drought in the core area of the China-Laos transportation corridor, 2001–2018. J. Ecol. 2021, 41, 2537–2547. [Google Scholar]
- Kong, D.; Zhang, Q.; Gu, X.; Wang, Y.; Li, H. Vegetation responses to drought at different time scales in China. Acta Ecol. Sin. 2016, 36, 7908–7979. [Google Scholar]
- Liu, Y.; You, C.; Zhang, Y.; Chen, S.; Zhang, Z.; Li, J.; Wu, Y. Resistance and resilience of grasslands to drought detected by SIF in inner Mongolia, China. Agric. For. Meteorol. 2021, 308, 108567. [Google Scholar] [CrossRef]
- Zhou, Z.; Zou, Y.; Zhang, L.; Wang, Z. Drought Characteristics and Causes in the Yellow River Basin from 1951 to 2013 Based on the SPEI Index. Inn. Mong. Sci. Technol. Econ. 2023, 12, 78–84. [Google Scholar]
- Jin, L.; Chen, S.; Liu, M. Multiscale Spatiotemporal Dynamics of Drought within the Yellow River Basin (YRB): An Examination of Regional Variability and Trends. Water 2024, 16, 791. [Google Scholar] [CrossRef]
- McDowell, N.G.; Sapes, G.; Pivovaroff, A.; Adams, H.D.; Allen, C.D.; Anderegg, W.R.L.; Arend, M.; Breshears, D.D.; Brodribb, T.; Choat, B. Mechanisms of woody-plant mortality under rising drought, CO2 and vapour pressure deficit. Nat. Rev. Earth Environ. 2022, 3, 294–308. [Google Scholar] [CrossRef]
- Huang, W.; Wang, W.; Cao, M.; Fu, G.; Xia, J.; Wang, Z.; Li, Z. Local climate and biodiversity affect the stability of China’s grasslands in response to drought. Sci. Total Environ. 2021, 768, 145482. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Gao, X.; Xu, T.; Xue, H.; He, W. Spatial-temporal evolution mechanism and efficiency evaluation of drought resilience system in China. J. Clean. Prod. 2023, 428, 139298. [Google Scholar] [CrossRef]
- Bai, J.; Shi, H.; Yu, Q.; Xie, Z.; Li, L.; Luo, G.; Jin, N.; Jun, L. Satellite-observed vegetation stability in response to changes in climate and total water storage in Central Asia. Sci. Total Environ. 2019, 659, 862–871. [Google Scholar] [CrossRef]
- Orians, G.H. Diversity, Stability and Maturity in Natural Ecosystems. In Unifying Concepts in Ecology; Springer: Berlin/Heidelberg, Germany, 1975; pp. 139–150. [Google Scholar]
- Xin, Z.; Wei, Z.; Hong, C. Resistance and Resilience Stability of Ecosystems. Biol. Teach. 2014, 39, 4–5. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).