
Assessing the Contrasting Effects of the Exceptional 2015 Drought on the Carbon Dynamics in Two Norway Spruce Forest Ecosystems

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Station Description
2.2. Eddy Covariance and Ancillary Measurements
2.3. Data Processing and Analysis
2.3.1. Turbulent Flux Measurements
2.3.2. Soil Water Content Simulations
2.3.3. Drought Stress Determination
2.3.4. Light Response Curve Fitting
2.3.5. Piecewise Regression Analyses for the Assessment of Drought Effect
3. Results
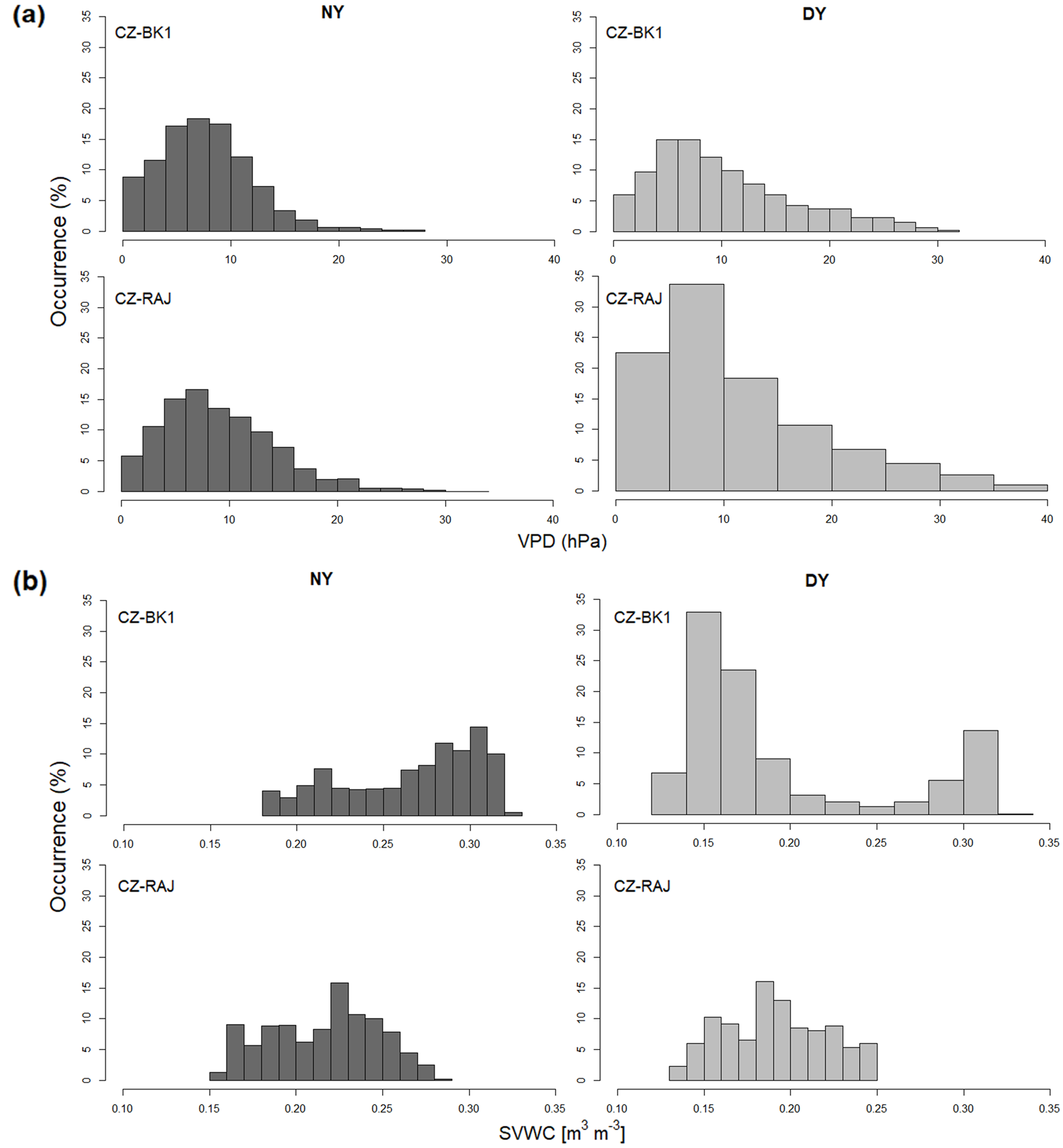
3.1. Differences in Meteorological Conditions at the Experimental Sites
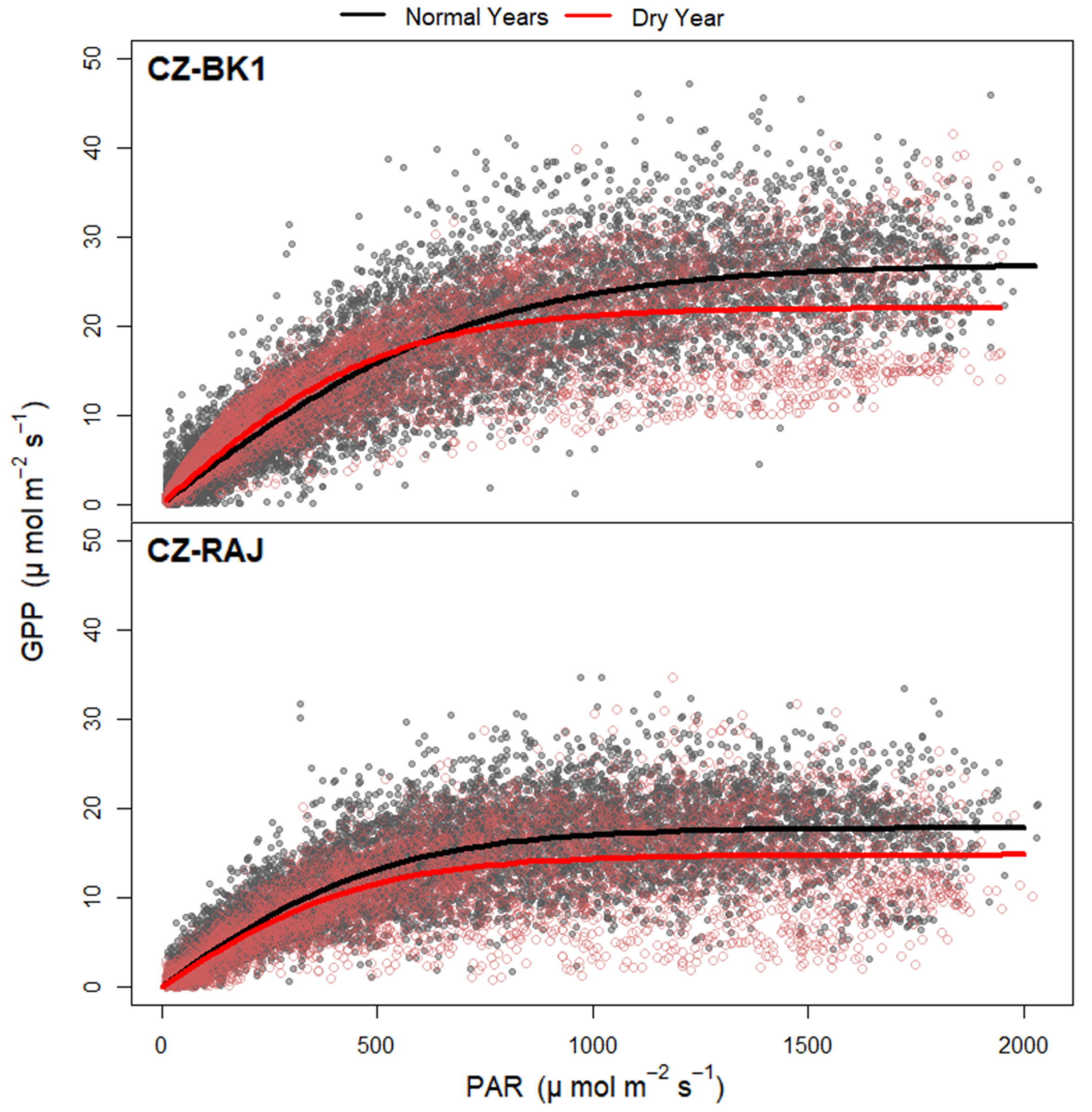
3.2. Light Response Curves of GPP at Different Climates
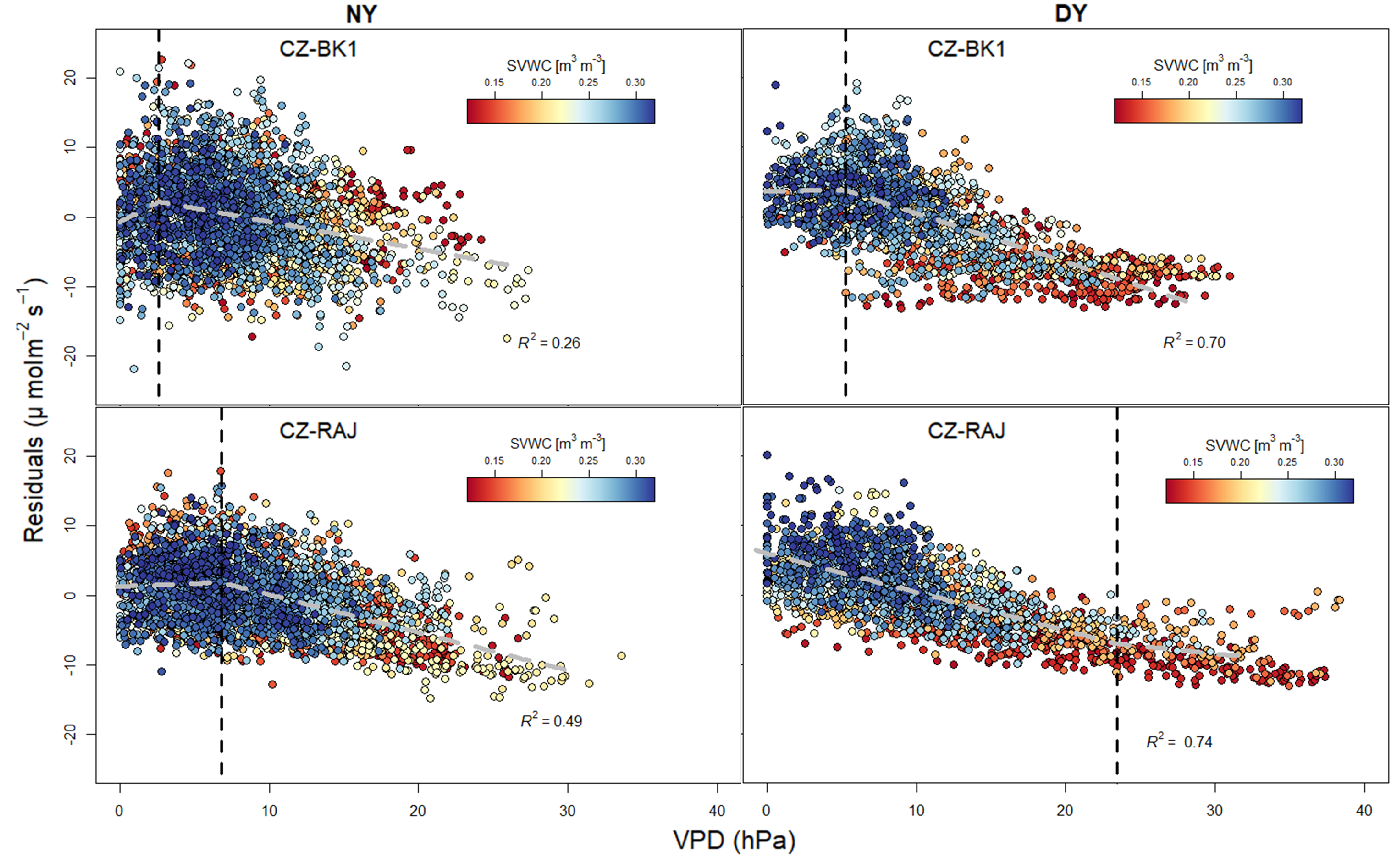
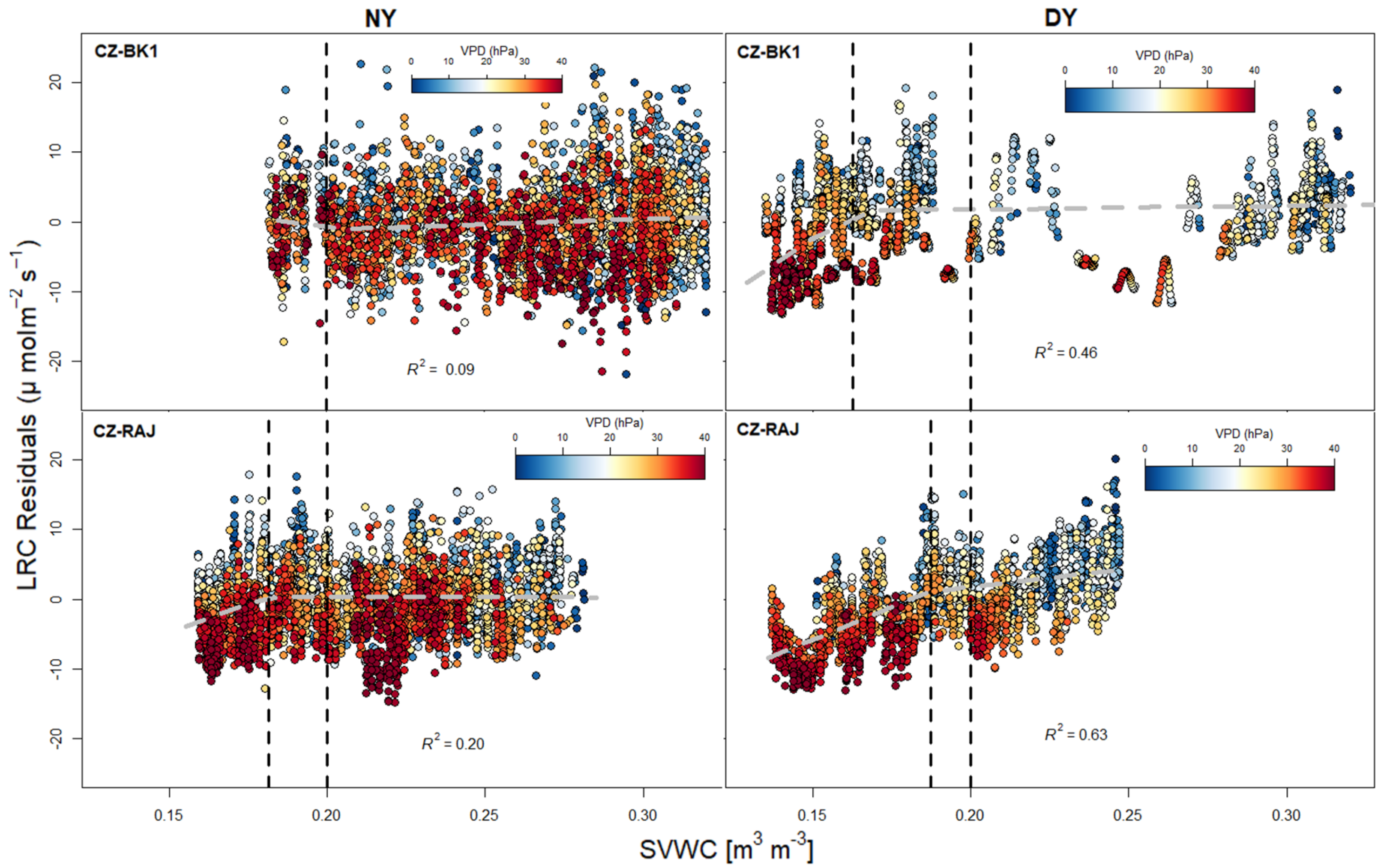
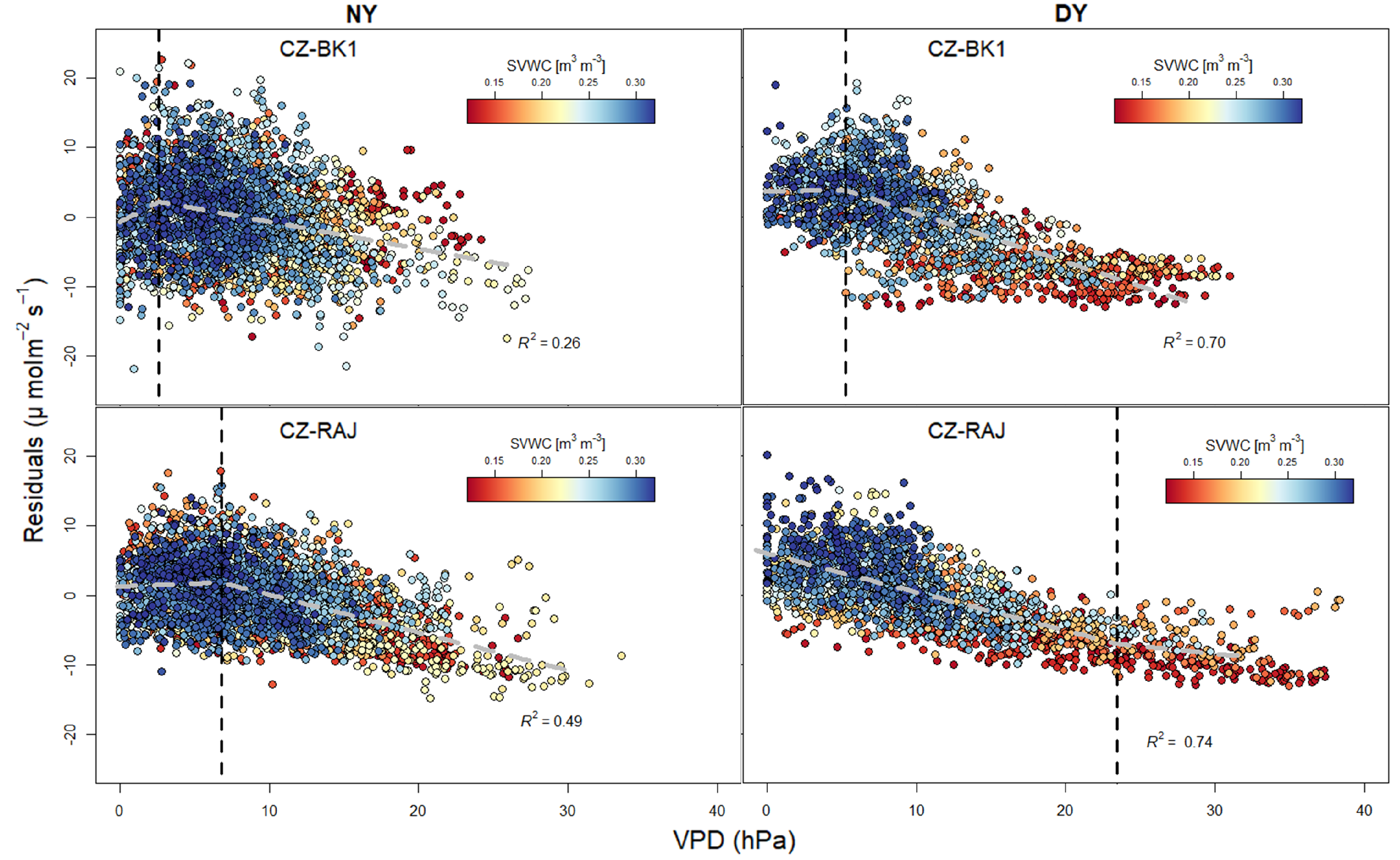
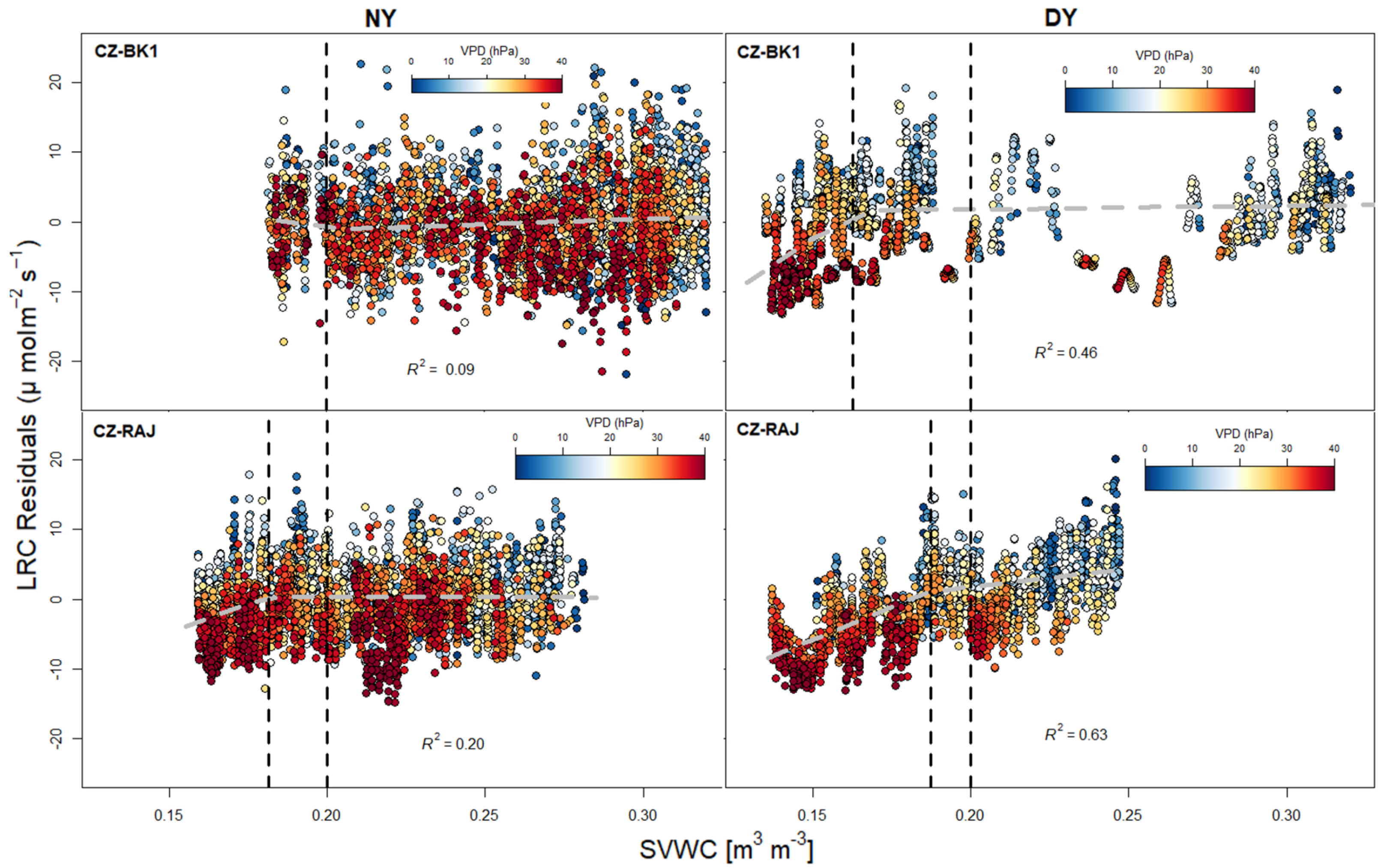
3.3. Response of Light Response Curve Residuals to VPD and SVWC at Different Climates
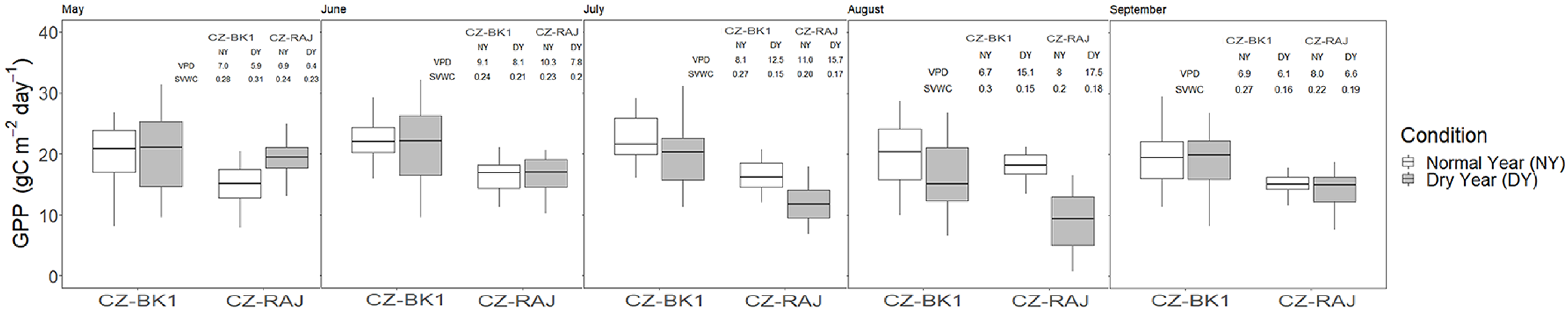
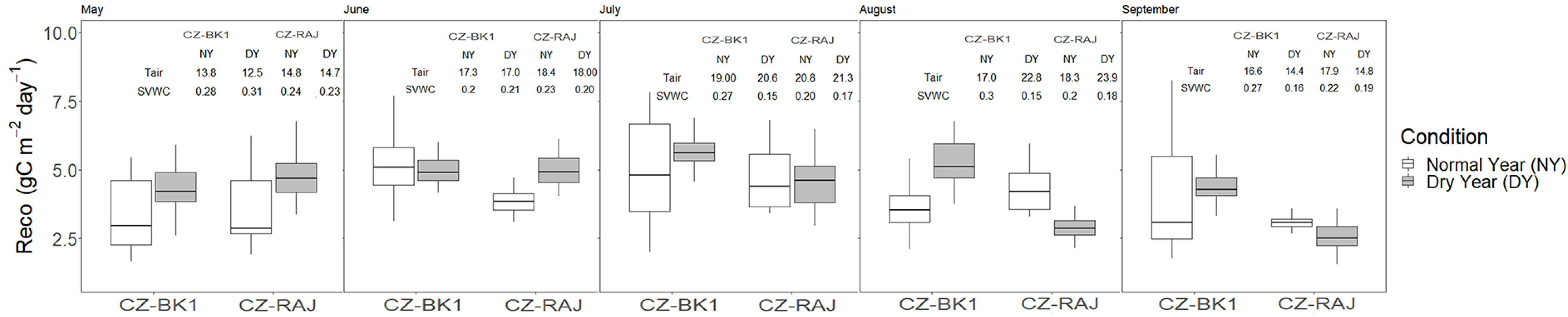
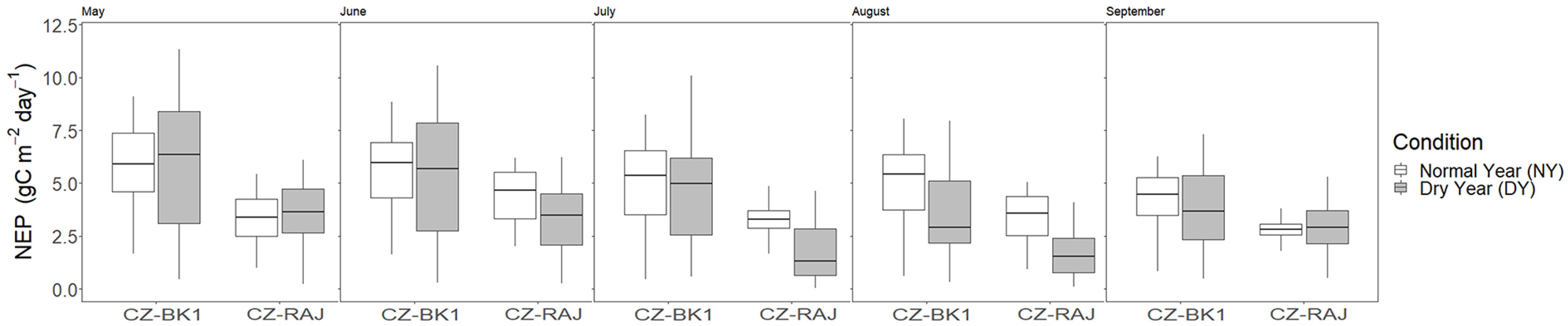
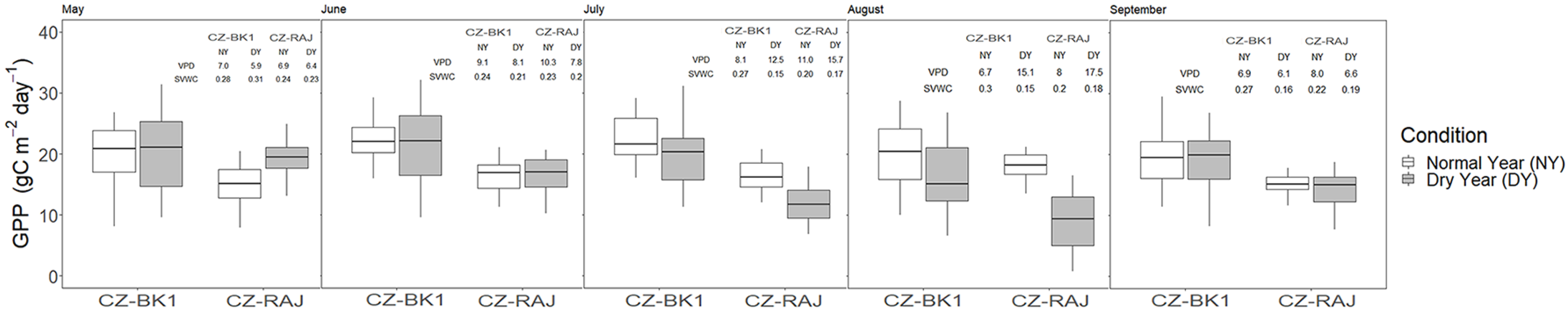
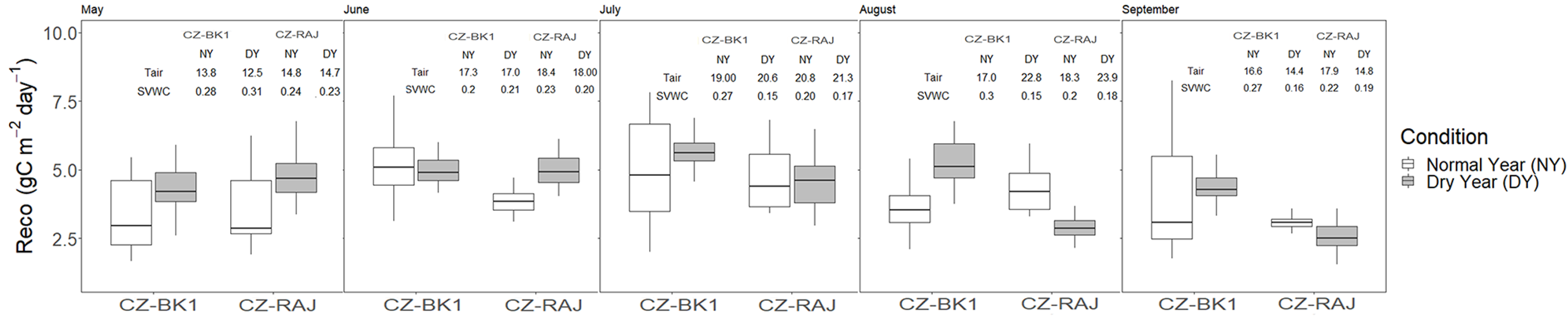
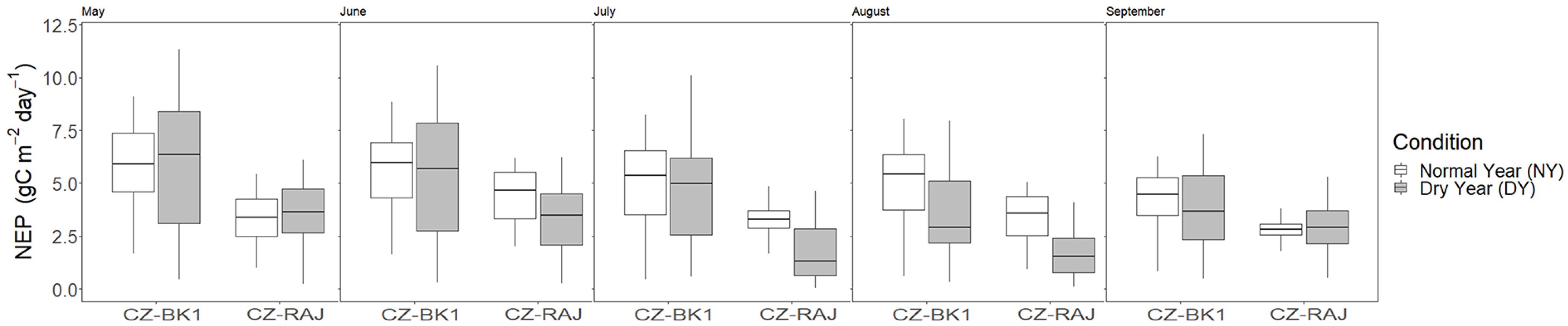
3.4. Impact of Drought on Ecosystem Carbon Fluxes at Monthly Timescale
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rossi, G.; Cancelliere, A.; Pereira, L.S. (Eds.) Tools for Drought Mitigation in Mediterranean Regions; Springer Science and Business Media: Berlin, Germany, 2003; Volume 44. [Google Scholar]
- Huntington, T.G. Evidence for intensification of the global water cycle: Review and synthesis. J. Hydrol. 2003, 319, 83–95. [Google Scholar] [CrossRef]
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [Green Version]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Gonzalez, P. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Marek, M.V.; Janouš, D.; Taufarová, K.; Havránková, K.; Pavelka, M.; Kaplan, V.; Marková, I. Carbon exchange between ecosystems and atmosphere in the Czech Republic is affected by climate factors. Environ. Pollut. 2011, 159, 1035–1039. [Google Scholar] [CrossRef]
- Lal, R.; Smith, P.; Jungkunst, H.F.; Mitsch, W.J.; Lehmann, J.; Nair, P.R.; Skorupa, A.L. The carbon sequestration potential of terrestrial ecosystems. J. Soil Water Conserv. 2018, 73, 145A–152A. [Google Scholar] [CrossRef] [Green Version]
- Novick, K.A.; Ficklin, D.L.; Stoy, P.C.; Williams, C.A.; Bohrer, G.; Oishi, A.C.; Papuga, S.A.; Blanken, P.D.; Noormets, A.; Sulman, B.N.; et al. The increasing importance of atmospheric demand for ecosystem water and carbon fluxes. Nat. Clim. Chang. 2016, 6, 1023–1027. [Google Scholar] [CrossRef] [Green Version]
- Kowalska, N.; Šigut, L.; Stojanović, M.; Fischer, M.; Kyselova, I.; Pavelka, M. Analysis of floodplain forest sensitivity to drought. Philos. Trans. R. Soc. B 2020, 375, 20190518. [Google Scholar] [CrossRef]
- Beer, C.; Reichstein, M.; Tomelleri, E.; Ciais, P.; Jung, M.; Carvalhais, N.; Rödenbeck, C.; Arain, M.A.; Baldocchi, D.; Bonan, G.B.; et al. Terrestrial gross carbon dioxide uptake: Global distribution and covariation with climate. Science 2010, 329, 834–838. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [Green Version]
- Cox, P.M.; Pearson, D.; Booth, B.B.; Friedlingstein, P.; Huntingford, C.; Jones, C.D.; Luke, C.M. Sensitivity of tropical carbon to climate change constrained by carbon dioxide variability. Nature 2013, 494, 341. [Google Scholar] [CrossRef]
- Le Quéré, C.; Andres, R.J.; Boden, T.; Conway, T.; Houghton, R.A.; House, J.I.; Marland, G.; Peters, G.P.; Van der Werf, G.; Ahlström, A.; et al. The global carbon budget 1959–2011. Earth Syst. Sci. Data 2013, 5, 165–185. [Google Scholar] [CrossRef] [Green Version]
- Krejza, J.; Cienciala, E.; Světlík, J.; Bellan, M.; Noyer, E.; Horáček, P.; Štěpánek, P.; Marek, M.V. Evidence of climate-induced stress of Norway spruce along elevation gradient preceding the current dieback in Central Europe. Trees 2020, 35, 103–119. [Google Scholar] [CrossRef]
- Zeng, N.; Qian, H.; Roedenbeck, C.; Heimann, M. Impact of 1998–2002 midlatitude drought and warming on terrestrial ecosystem and the global carbon cycle. Geophys. Res. Lett. 2005, 32. [Google Scholar] [CrossRef] [Green Version]
- Dai, A. Increasing drought under global warming in observations and models. Nat. Clim. Chang. 2013, 3, 52. [Google Scholar] [CrossRef]
- Reichstein, M.; Bahn, M.; Ciais, P.; Frank, D.; Mahecha, M.D.; Seneviratne, S.I.; Zscheischler, J.; Beer, C.; Buchmann, N.; Frank, D.C.; et al. Climate extremes and the carbon cycle. Nature 2013, 500, 287. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, W.; Zhou, W. Assessment of future drought in Southwest China based on CMIP5 multimodel projections. Adv. Atmos. Sci. 2014, 31, 1035–1050. [Google Scholar] [CrossRef]
- Xie, Z.; Wang, L.; Jia, B.; Yuan, X. Measuring and modeling the impact of a severe drought on terrestrial ecosystem CO2 and water fluxes in a subtropical forest. J. Geophys. Res. Biogeosci. 2016, 121, 2576–2587. [Google Scholar] [CrossRef] [Green Version]
- Zang, C.; Hartl-Meier, C.; Dittmar, C.; Rothe, A.; Menzel, A. Patterns of drought tolerance in major European temperate forest trees: Climatic drivers and levels of variability. Glob. Chang. Biol. 2014, 20, 3767–3779. [Google Scholar] [CrossRef]
- De Boeck, H.J.; Verbeeck, H. Drought-associated changes in climate and their relevance for ecosystem experiments and models. Biogeosciences 2011, 8, 1121–1130. [Google Scholar] [CrossRef] [Green Version]
- Hui, D.; Deng, Q.; Tian, H.; Luo, Y. Climate change and carbon sequestration in forest ecosystems. In Handbook of Climate Change Mitigation and Adaptation; Chen, W.Y., Suzuki, T., Lackner, M., Eds.; Springer International Publishing: New York, NY, USA, 2017; pp. 555–594. [Google Scholar]
- Qie, L.; Lewis, S.L.; Sullivan, M.J.; Lopez-Gonzalez, G.; Pickavance, G.C.; Sunderland, T.; Ashton, P.; Hubau, W.; Salim, K.A.; Aiba, S.I.; et al. Long-term carbon sink in Borneo’s forests halted by drought and vulnerable to edge effects. Nat. Commun. 2017, 8, 1966. [Google Scholar] [CrossRef] [Green Version]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529. [Google Scholar] [CrossRef]
- Granier, A.; Reichstein, M.; Breda, N.; Janssens, I.A.; Falge, E.; Ciais, P.; Grünwald, T.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; et al. Evidence for soil water control on carbon and water dynamics in European forests during the extremely dry year: 2003. Agric. For. Meteorol. 2007, 143, 123–145. [Google Scholar] [CrossRef]
- Trnka, M.; Hlavinka, P.; Možný, M.; Semerádová, D.; Štěpánek, P.; Balek, J.; Bartošová, L.; Zahradníček, P.; Bláhová, M.; Skalak, P.; et al. Czech Drought Monitor System for Monitoring and Forecasting Agricultural Drought and Drought Impacts. Int. J. Climatol. 2020, 40, 5941–5958. [Google Scholar] [CrossRef]
- Orth, R.; Seneviratne, S.I. Introduction of a simple-model-based land surface dataset for Europe. Environ. Res. Lett. 2015, 10, 0442012. [Google Scholar]
- Orth, R.; Zscheischler, J.; Seneviratne, S.I. Record dry summer in 2015 challenges precipitation projections in Central Europe. Sci. Rep. 2016, 6, 28334. [Google Scholar] [CrossRef] [PubMed]
- Van Lanen, H.A.; Laaha, G.; Kingston, D.G.; Gauster, T.; Ionita, M.; Vidal, J.P.; Vlnas, R.; Tallaksen, L.M.; Stahl, K.; Hannaford, J.; et al. Hydrology needed to manage droughts: The 2015 European case. Hydrol. Process. 2016, 30, 3097–3104. [Google Scholar] [CrossRef] [Green Version]
- Ionita, M.; Tallaksen, L.M.; Kingston, D.G.; Stagge, J.H.; Laaha, G.; Van Lanen, H.A.; Scholz, P.; Chelcea, S.M.; Haslinger, K. The European 2015 drought from a climatological perspective. Hydrol. Earth Syst. Sci. 2017, 21, 1397–1419. [Google Scholar] [CrossRef] [Green Version]
- Krupková, L.; Havránková, K.; Krejza, J.; Sedlák, P.; Marek, M.V. Impact of water scarcity on spruce and beech forests. J. For. Res. 2018, 30, 899–909. [Google Scholar] [CrossRef]
- McGloin, R.; Šigut, L.; Havránková, K.; Dušek, J.; Pavelka, M.; Sedlák, P. Energy balance closure at a variety of ecosystems in Central Europe with contrasting topographies. Agric. For. Meteorol. 2018, 248, 418–431. [Google Scholar] [CrossRef]
- Kenderes, K.; Mihók, B.; Standovár, T. Thirty years of gap dynamics in a Central European beech forest reserve. Forestry 2008, 81, 111–123. [Google Scholar] [CrossRef]
- Nadezhdina, N.; Urban, J.; Čermák, J.; Nadezhdin, V.; Kantor, P. Comparative study of long-term water uptake of Norway spruce and Douglas-fir in Moravian upland. J. Hydrol. Hydromech. 2014, 62, 1–6. [Google Scholar] [CrossRef]
- Kmet, J.; Ditmarová, L.; Kurjak, D.; Priwitzer, T. Physiological response of Norway spruce foliage in the drought vegetation period 2009. Beskydy 2011, 4, 109–118. [Google Scholar]
- Aubinet, M.; Vesala, T.; Papale, D. (Eds.) Eddy Covariance: A Practical Huide to Measurement and Data Analysis; Springer Science and Business Media: Berlin, Germany, 2012. [Google Scholar]
- Baldocchi, D.D. Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems: Past, present and future. Glob. Chang. Biol. 2003, 9, 479–492. [Google Scholar] [CrossRef] [Green Version]
- Sedlák, P.; Aubinet, M.; Heinesch, B.; Janouš, D.; Pavelka, M.; Potužníková, K.; Yernaux, M. Night-time airflow in a forest canopy near a mountain crest. Agric. For. Meteorol. 2010, 150, 736–744. [Google Scholar] [CrossRef]
- Urban, O.; Klem, K.; Holišová, P.; Šigut, L.; Šprtová, M.; Teslová-Navrátilová, P.; Zitová, M.; Špunda, V.; Marek, M.V.; Grace, J. Impact of elevated CO2 concentration on dynamics of leaf photosynthesis in Fagus sylvatica is modulated by sky conditions. Environ. Pollut. 2014, 185, 271–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteith, J.; Unsworth, M. Principles of Environmental Physics: Plants, Animals, and the Atmosphere; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Foken, T.; Leuning, R.; Oncley, S.R.; Mauder, M.; Aubinet, M. Corrections and data quality control. In Eddy Covariance; Springer: Dordrecht, The Netherlands, 2012; pp. 85–131. [Google Scholar]
- Wilczak, J.M.; Oncley, S.P.; Stage, S.A. Sonic anemometer tilt correction algorithms. Bound. Layer Meteorol. 2001, 99, 127–150. [Google Scholar] [CrossRef]
- Ibrom, A.; Dellwik, E.; Larsen, S.E.; Pilegaard, K.I.M. On the use of the Webb-Pearman-Leuning theory for closed-path eddy correlation measurements. Tellus B Chem. Phys. Meteorol. 2007, 59, 937–946. [Google Scholar] [CrossRef]
- Horst, T.W.; Lenschow, D.H. Attenuation of scalar fluxes measured with spatially-displaced sensors. Bound. Layer Meteorol. 2009, 130, 275–300. [Google Scholar] [CrossRef]
- Moncrieff, J.B.; Jarvis, P.G.; Valentini, R. Canopy fluxes. In Methods in Ecosystem Science; Springer: New York, NY, USA, 2000; pp. 161–180. [Google Scholar]
- Papale, D.; Reichstein, M.; Aubinet, M.; Canfora, E.; Bernhofer, C.; Kutsch, W.; Longdoz, B.; Rambal, S.; Valentini, R.; Vesala, T.; et al. Towards a standardized processing of Net Ecosystem Exchange measured with eddy covariance technique: Algorithms and uncertainty estimation. Biogeosciences 2006, 3, 571–583. [Google Scholar] [CrossRef] [Green Version]
- Reichstein, M.; Falge, E.; Baldocchi, D.; Papale, D.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; Buchmann, N.; Gilmanov, T.; Granier, A.; et al. On the separation of net ecosystem exchange into assimilation and ecosystem respiration: Review and improved algorithm. Glob. Chang. Biol. 2005, 11, 1424–1439. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Wutzler, T.; Lucas-Moffat, A.; Migliavacca, M.; Knauer, J.; Sickel, K.; Šigut, L.; Menzer, O.; Reichstein, M. Basic and extensible post-processing of eddy covariance flux data with REddyProc. Biogeosciences 2018, 15, 5015–5030. [Google Scholar] [CrossRef] [Green Version]
- Lasslop, G.; Reichstein, M.; Papale, D.; Richardson, A.D.; Arneth, A.; Barr, A.; Stoy, P.; Wohlfahrt, G. Separation of net ecosystem exchange into assimilation and respiration using a light response curve approach: Critical issues and global evaluation. Glob. Chang. Biol. 2010, 16, 187–208. [Google Scholar] [CrossRef] [Green Version]
- Fischer, M.; Zenone, T.; Trnka, M.; Orság, M.; Montagnani, L.; Ward, E.J.; Tripathi, A.M.; Hlavinka, P.; Seufert, G.; Žalud, Z.; et al. Water requirements of short rotation poplar coppice: Experimental and modelling analyses across Europe. Agric. For. Meteorol. 2018, 250, 343–360. [Google Scholar] [CrossRef]
- Hartig, F.; Minunno, F.; Paul, S. BayesianTools: General-Purpose MCMC and SMC Samplers and Tools for Bayesian Statistics. R Package Version 0.1.6. 2019. Available online: https://CRAN.R-project.org/package=BayesianTools (accessed on 20 March 2021).
- Gelman, A.; Rubin, D.B. Inference from iterative simulation using multiple sequences. Stat. Sci. 1992, 7, 457–472. [Google Scholar] [CrossRef]
- Brooks, S.P.; Gelman, A. General methods for monitoring convergence of iterative simulations. J. Comput. Graph. Stat. 1998, 7, 434–455. [Google Scholar]
- Jackson, R.B.; Canadell, J.; Ehleringer, J.R.; Mooney, H.A.; Sala, O.E.; Schulze, E.D. A global analysis of root distributions for terrestrial biomes. Oecologia 1996, 108, 389–411. [Google Scholar] [CrossRef] [PubMed]
- Oren, R.; Sperry, J.S.; Katul, G.G.; Pataki, D.E.; Ewers, B.E.; Phillips, N.; Scháfer, K.V.R. Survey and synthesis of intra-and interspecific variation in stomatal sensitivity to vapour pressure deficit. Plant Cell Environ. 1999, 22, 1515–1526. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.K.; Singh, V.P. A review of drought concepts. J. Hydrol. 2010, 391, 202–216. [Google Scholar] [CrossRef]
- Beguería, S.; Vicente-Serrano, S.M.; Reig, F.; Latorre, B. Standardized precipitation evapotranspiration index (SPEI) revisited: Parameter fitting, evapotranspiration models, tools, datasets and drought monitoring. Int. J. Climatol. 2014, 34, 3001–3023. [Google Scholar] [CrossRef] [Green Version]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A Multiscalar Drought Index Sensitive to Global Warming: The Standardized Precipitation Evapotranspiration Index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef] [Green Version]
- Beguería, S.; Vicente-Serrano, S.M. SPEI: Calculation of the Standardised Precipitation-Evapotranspiration Index. R Package Version 1.7. 2017. Available online: https://CRAN.R-project.org/package=SPEI (accessed on 13 January 2021).
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop evapotranspiration—Guidelines for computing crop water requirements. FAO Irrig. Drain. Pap. 1998, 56, 290. [Google Scholar]
- Moffat, A.M. A New Methodology to Interpret High Resolution Measurements of Net Carbon Fluxes between Terrestrial Ecosystems and the Atmosphere. Ph.D. Thesis, Friedrich-Schiller-Universität Jena, Fakultät für Mathematik und Informatik, Jena, Germany, 2012. [Google Scholar]
- Migliavacca, M.; Reichstein, M.; Richardson, A.D.; Colombo, R.; Sutton, M.A.; Lasslop, G.; Tomelleri, E.; Wohlfahrt, G.; Carvalhais, N.; Cescatti, A.; et al. Semiempirical modeling of abiotic and biotic factors controlling ecosystem respiration across eddy covariance sites. Glob. Chang. Biol. 2011, 17, 390–409. [Google Scholar] [CrossRef]
- Muggeo, V.M. Estimating regression models with unknown break-points. Stat. Med. 2003, 22, 3055–3071. [Google Scholar] [CrossRef]
- Davies, R.B. Hypothesis testing when a nuisance parameter is present only under the alternative: Linear model case. Biometrika 2002, 89, 484–489. [Google Scholar] [CrossRef]
- Reichstein, M.; Ciais, P.; Papale, D.; Valentini, R.; Running, S.; Viovy, N.; Cramer, W.; Granier, A.; Ogee, J.; Allard, V.; et al. Reduction of ecosystem productivity and respiration during the European summer 2003 climate anomaly: A joint flux tower, remote sensing and modelling analysis. Glob. Chang. Biol. 2007, 13, 634–651. [Google Scholar] [CrossRef]
- Jassal, R.S.; Black, T.A.; Chen, B.; Roy, R.; Nesic, Z.; Spittlehouse, D.L.; Trofymow, J.A. N2O emissions and carbon sequestration in a nitrogen-fertilized Douglas fir stand. J. Geophys. Res. Biogeosci. 2008, 113. [Google Scholar] [CrossRef] [Green Version]
- Mahecha, M.D.; Reichstein, M.; Carvalhais, N.; Lasslop, G.; Lange, H.; Seneviratne, S.I.; Vargas, R.; Ammann, C.; Arain, M.A.; Cescatti, A.; et al. Global convergence in the temperature sensitivity of respiration at ecosystem level. Science 2010, 329, 838–840. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.H.; Jansson, P.E.; Kolari, P. Modeling seasonal course of carbon fluxes and evapotranspiration in response to low temperature and moisture in a boreal Scots pine ecosystem. Ecol. Model. 2011, 222, 3103–3119. [Google Scholar] [CrossRef]
- Frank, D.; Reichstein, M.; Bahn, M.; Thonicke, K.; Frank, D.; Mahecha, M.D.; Smith, P.; Van der Velde, M.; Vicca, S.; Babst, F.; et al. Effects of climate extremes on the terrestrial carbon cycle: Concepts, processes and potential future impacts. Glob. Chang. Biol. 2015, 21, 2861–2880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Gorsel, E.; Wolf, S.; Cleverly, J.; Isaac, P.; Haverd, V.; Ewenz, C.; Arndt, S.; Beringer, J.; Resco de Dios, V.; Evans, B.J.; et al. Carbon uptake and water use in woodlands and forests in southern Australia during an extreme heat wave event in the “angry Summer” of 2012/2013. Biogeosciences 2016, 13, 5947–5964. [Google Scholar] [CrossRef] [Green Version]
- von Buttlar, J.; Zscheischler, J.; Rammig, A.; Sippel, S.; Reichstein, M.; Knohl, A.; Jung, M.; Menzer, O.; Arain, M.A.; Buchmann, N.; et al. Impacts of droughts and extreme-temperature events on gross primary production and ecosystem respiration: A systematic assessment across ecosystems and climate zones. Biogeosciences 2018, 15, 1293–1318. [Google Scholar] [CrossRef] [Green Version]
- Ruehr, N.K.; Law, B.E.; Quandt, D.; Williams, M. Effects of heat and drought on carbon and water dynamics in a regenerating semi-arid pine forest: A combined experimental and modeling approach. Biogeosciences 2014, 11, 4139–4156. [Google Scholar] [CrossRef] [Green Version]
- Sulman, B.N.; Roman, D.T.; Yi, K.; Wang, L.; Phillips, R.P.; Novick, K.A. High atmospheric demand for water can limit forest carbon uptake and transpiration as severely as dry soil. Geophys. Res. Lett. 2016, 43, 9686–9695. [Google Scholar] [CrossRef]
- Sheffield, J.; Wood, E.F.; Roderick, M.L. Little change in global drought over the past 60 years. Nature 2012, 491, 435–438. [Google Scholar] [CrossRef] [PubMed]
- Potopová, V.; Boroneant, C.; Možný, M.; Soukup, J. Driving role of snow cover on soil moisture and drought development during the growing season in the Czech Republic. Int. J. Climatol. 2016, 36, 3741–3758. [Google Scholar] [CrossRef] [Green Version]
- Laaha, G.; Gauster, T.; Tallaksen, L.M.; Vidal, J.P.; Stahl, K.; Prudhomme, C.; Heudorfer, B.; Vlnas, R.; Ionita, M.; Van Lanen, H.A.; et al. The European 2015 drought from a hydrological perspective. Hydrol. Earth Syst. Sci. 2017, 21, 3001–3024. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H.; Schütze, G.; Biber, P. Drought can favour the growth of small in relation to tall trees in mature stands of Norway spruce and European beech. For. Ecosyst. 2018, 5, 20. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Name | CZ-BK1 | CZ-RAJ |
|---|---|---|
| Location | Moravian-Silesian Beskids Mountains | Drahany Highland |
| Coordinates | 493008 N, 183213 E | 492637 N, 164148 E |
| Elevation (in m a.s.l) | 875 | 625 |
| Topography | Mountainous (13 slope with SSW exposure, located close to a mountain ridge) | Hilly (5 slope with NNE exposure) |
| Ecosystem Type | Coniferous evergreen forest | Coniferous evergreen forest |
| Prevailing species | Norway Spruce (Picea abies (L.) H.Karst.) | Norway Spruce (Picea abies (L.) H.Karst.) |
| Canopy height (m) | 16 (mean, as of 2015) | 33 (mean, as of 2015) |
| Stand age (years) | 35 (as of 2016) | 113 (as of 2016) |
| Mean annual air temperature (May–September; C) | 7.2 (2014–2016) | 8.3 (2014–2016) |
| Mean annual precipitation (May–September; mm) | 1143 (2014–2016) | 610 (2014–2016) |
| Mean annual reference evapotranspiration (May–September; mm) | 569 (2014–2016) | 649 (2014–2016) |
| Soil type | Haplic and Entic Podzol | Modal Cambisol oligotrophic |
| Max and min fetch (m) | 717 (WNW) and 115 (ENE) | 697 (SSW) and 96 (NNW) |
| References | [31,37,38] | [31] |
| Site Name | CZ-BK1 | CZ-RAJ |
|---|---|---|
| Ultrasonic Anemometer | ||
| Instrument | Gill HS-50 ultrasonic anemometer, Gill Instruments, Hampshire, U.K. | Gill R3-100 ultrasonic anemometer, Gill Instruments, Hampshire, U.K. but later changed to Gill HS-50 on 5 June 2015 |
| Gas Analyser | ||
| Instrument | LI-7200 enclosed gas analyser, LI-COR, Lincoln, U.S.A. | Initially LI-7000 (IRG-0226) closed-path gas analyser, LI-COR, Lincoln, U.S.A. but later changed to LI-7200 on 5 June 2015 |
| Measurement Height for the Eddy covariance Set-up (m) | Initially 20.5, but later changed to 25 m on 7 June 2016 | 41 |
| Air Temperature and Humidity Profile | ||
| Instrument | EMS33 temperature and humidity sensor (EMS, Brno, CZ) | EMS33 temperature and humidity sensor (EMS, Brno, CZ) |
| Measurement Height (m) | 2.0, 7.6, 12.6, 13.5, 14.3, 14.8, 15.4, 16.5, 18.7 | 2.0, 11.0, 23.0, 29.0, 35.0, 42.0 |
| Net Radiation | ||
| Instrument | CNR1 Net Radiometer | CNR1 Net Radiometer |
| Measurement Height (m) | 22 | 42 |
| Soil Moisture | ||
| Instrument | CS616 (Campbell Scientific, Inc., Logan, UT, U.S.A.) | CS616 (Campbell Scientific, Inc., Logan, UT, U.S.A.) |
| Measurement Depths (m) | 0.05, 0.1, 0.22, 0.34, 0.42 | 0.05, 0.1, 0.2, 0.5, 0.8 |
| Drought/ Wet Severity | SPEI Value |
|---|---|
| Extremely Wet | ≥2.00 |
| Severely Wet | 1.50–1.99 |
| Moderately Wet | 1.00–1.49 |
| Near Normal | −0.99–(0.99) |
| Moderate Drought | −1.00–(−1.49) |
| Severe Drought | −1.50–(−1.99) |
| Extreme Drought | ≤−2.00 |
| YEARS | CZ-BK1 | CZ-RAJ | ||
|---|---|---|---|---|
| SPEI VALUE | CLASS | SPEI VALUE | CLASS | |
| 2014 | 0.94 | Near Normal | 0.56 | Near Normal |
| 2015 | −1.75 | Severe Drought | −1.55 | Severe Drought |
| 2016 | −0.20 | Near Normal | −0.87 | Near Normal |
| YEARS | CZ-BK1 | CZ-RAJ | ||||
|---|---|---|---|---|---|---|
| VPD (hPa) | Tair (C) | SVWC (m3 −3) | VPD (hPa) | Tair (C) | SVWC (m3 −3) | |
| 2014 | 7.6 | 13.4 | 0.28 | 8.7 | 14.9 | 0.21 |
| 2015 | 10.2 | 14.6 | 0.19 | 11.2 | 15.8 | 0.19 |
| 2016 | 7.7 | 14.4 | 0.25 | 9.3 | 16.0 | 0.22 |
| Variants | CZ-BK1 | CZ-RAJ | ||
|---|---|---|---|---|
| Years with Normal Conditions (2014 & 2016) | Dry Year (2015) | Years with Normal Conditions (2014 & 2016) | Dry Year (2015) | |
| [mol (CO2) mol−1 (phot.)] | 0.0383 ± 0.0003 | 0.0446 ± 0.0006 | 0.0338 ± 0.0003 | 0.0312 ± 0.0006 |
| GPPmax[mol m−2 s−1] | 26.91 ± 0.14 | 22.03 ± 0.13 | 17.77 ± 0.08 | 14.75 ± 0.12 |
| R2 | 0.88 | 0.78 | 0.78 | 0.73 |
| Variants | CZ-BK1 | CZ-RAJ | ||
|---|---|---|---|---|
| Years with Normal Conditions (2014 & 2016) | Dry Year (2015) | Years with Normal Conditions (2014 & 2016) | Dry Year (2015) | |
| VPD [hPa] | 2.6 | 5.3 | 6.8 | 23.5 |
| Slope before breakpoint in VPD | 1.12 ± 0.30 | 0.02 ± 0.12 | 0.08 ± 0.05 | −0.55 ± 0.01 |
| Slope after breakpoint in VPD | −0.39 ± 0.02 | −0.70 ± 0.02 | −0.54 ± 0.02 | −0.22 ± 0.06 |
| SVWC [m3 m−3] | 0.20 | 0.16 | 0.18 | 0.19 |
| Slope before breakpoint in SVWC | −85.49 ± 68.37 | 305.61 ± 23.43 | 164.61 ± 25.14 | 187.22 ± 8.78 |
| Slope after breakpoint in SVWC | 14.75 ± 2.60 | 5.03 ± 2.51 | −1.27 ± 2.84 | 52.16 ± 6.24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mensah, C.; Šigut, L.; Fischer, M.; Foltýnová, L.; Jocher, G.; Acosta, M.; Kowalska, N.; Kokrda, L.; Pavelka, M.; Marshall, J.D.; et al. Assessing the Contrasting Effects of the Exceptional 2015 Drought on the Carbon Dynamics in Two Norway Spruce Forest Ecosystems. Atmosphere 2021, 12, 988. https://doi.org/10.3390/atmos12080988
Mensah C, Šigut L, Fischer M, Foltýnová L, Jocher G, Acosta M, Kowalska N, Kokrda L, Pavelka M, Marshall JD, et al. Assessing the Contrasting Effects of the Exceptional 2015 Drought on the Carbon Dynamics in Two Norway Spruce Forest Ecosystems. Atmosphere. 2021; 12(8):988. https://doi.org/10.3390/atmos12080988
Chicago/Turabian StyleMensah, Caleb, Ladislav Šigut, Milan Fischer, Lenka Foltýnová, Georg Jocher, Manuel Acosta, Natalia Kowalska, Lukáš Kokrda, Marian Pavelka, John David Marshall, and et al. 2021. "Assessing the Contrasting Effects of the Exceptional 2015 Drought on the Carbon Dynamics in Two Norway Spruce Forest Ecosystems" Atmosphere 12, no. 8: 988. https://doi.org/10.3390/atmos12080988
APA StyleMensah, C., Šigut, L., Fischer, M., Foltýnová, L., Jocher, G., Acosta, M., Kowalska, N., Kokrda, L., Pavelka, M., Marshall, J. D., Nyantakyi, E. K., & Marek, M. V. (2021). Assessing the Contrasting Effects of the Exceptional 2015 Drought on the Carbon Dynamics in Two Norway Spruce Forest Ecosystems. Atmosphere, 12(8), 988. https://doi.org/10.3390/atmos12080988