Abstract
Forests play a pivotal role in mitigating global warming as an important carbon sink. Recent global greening trends reflect a positive influence of elevated atmospheric CO2 on terrestrial carbon uptake. However, increasingly frequent and intense drought events endanger the carbon sequestration function of forests. This review integrates previous studies across scales to identify potential global trends in forest responses to drought and elevated CO2 as well as to identify data needs in this important research field. The inconsistent responses of ecosystem respiration to drought contributes to the change of forest net CO2 exchange, which depends on the balance of opposite effects of warming and water stress on respiration. Whether CO2 fertilization can offset the effects of drought remains controversial, however, we found a potential overestimation of global CO2 fertilization effects because of increasing water stress and other limitations such as light and nutrients (N, P) as well as the possibility of photosynthetic acclimation.
1. Introduction
Forest ecosystems play a key role in the land-atmosphere system and provide wide ecological services and socio-economic benefits, including forest products, water regulation, biodiversity, soil protection, spiritual and inspirational needs, recreational activities and aesthetic value [1]. Additionally, climate is influenced and regulated by the interaction of forests and atmosphere through the exchange of energy, water, CO2 and other chemical compounds. Forests capture atmospheric CO2 and sequester it into their biomass and soil, and then release oxygen back to the atmosphere, which is the buffer against global warming. Forty-five percent of the terrestrial carbon is stored in forest biomass, organic matter and soil, which is one of the largest global carbon pools [1]. Terrestrial ecosystems take up around 3 petagrams (Pg) of carbon emitted by human activities per year and capture nearly 30% of the CO2 released by logging and the combustion of fossil fuel [2]. However, small shifts in the balance of forest ecosystem photosynthesis and respiration can lead to huge changes in the forest carbon cycle [3]. For example, elevated atmospheric CO2 promotes the rate of plant photosynthesis and enhances water use efficiency (WUE) by reducing stomatal conductance, which can offset part of carbon emissions without more water consumption [4].
The increase of plant photosynthetic rate by CO2 fertilization effects is considered one of the potential reasons for the observed global greening trends [5]. Higher WUE under elevated atmospheric CO2 makes plants more tolerant to water stress [6], however, frequent occurrence of extreme droughts and heat stress lead to the decline of forest productivity and biomass accrual by widespread tree mortality and wildfires [7]. The balance between drought-induced forest carbon loss and enhanced carbon uptake by CO2 fertilization depends on the severity and duration of droughts. The extent to which climate change affects forest carbon uptake and the interaction of drought and elevated CO2 on plant stress resistance is still unclear. We review the current understanding in the response of the forest carbon cycle to droughts and elevated CO2 and highlight a research gap in the interaction of drought and elevated CO2 on forest ecosystems. Since the global warming and severity of extreme events will continue to increase if greenhouse gas emissions are unmitigated, understanding the implications of the capacity of forests to sequester carbon under climate scenarios remains a pressing and crucial need.
2. Drought and Forest Carbon Cycle
Multiple concepts and indices of drought are applied in drought-related studies, which reflects the inconsistent indicators used to identify drought events. Climate change exacerbates the frequency, intensity and duration of extreme droughts, which directly impairs the carbon sequestration function of forest ecosystems. We clarify the definition of drought and discuss forest carbon cycle feedback to drought below.
2.1. Drought Definition
Kelly Redmond [8] generally describes drought as ‘insufficient water to meet needs’. The moisture of terrestrial ecosystems is primarily provided by precipitation, but water demand comes from numerous places, for example, plant transpiration, land surface evaporation, agricultural irrigation and urban water use [9]. Therefore, although studies related to terrestrial ecosystem responses to droughts have a long history, there is no unified or standard definition for drought. Instead, researchers have characterized and defined drought in various ways according to the impacts of drought. For instance, drought can be described as precipitation decrease, low soil moisture, low water flow and plants facing water stress compared with normal conditions [10]. Multiple drought indices have been widely used to quantitatively define drought events, such as the Standardized Precipitation Index (SPI), the Standardized Precipitation-Evapotranspiration Index (SPEI), the Palmer Drought Severity Index (PDSI) and the self-calibrate PDSI (sc-PDSI). These indices require diverse input data and reflect different concepts of drought. For example, the SPI calculation only requires the time series of precipitation but does not consider the impact of evapotranspiration. SPEI was developed to compensate for this absence, which uses the same statistical method as SPI, but the input precipitation data is replaced with the difference between precipitation and potential evapotranspiration data (which is related to air temperature). Moreover, indices which perform well in specific regions may be unsuitable for other regions with different climate patterns and topographic conditions. Eslamian et al. [11] indicate that the PDSI performs better in wide plains than other indexes, such as in the United States and Australia. The Surface Water Supply Index (SWSI) is more suitable for the less-flat mountainous region [11]. A review focused on drought definition indicates that studies often confuse drought with dry conditions [10]. They calculated the SPEI for a group of drought-related studies, which did not quantify drought with specific indices, and found that 50% of drought events in those studies were defined as a normal condition by the SPEI (−1 < SPEI < 1). This surprising result may be attributed to the fact that the drought events in these studies had a shorter duration than the timescales of calculation [10].
Inconsistent definitions of drought may result in uncertainties in assessments of drought and drought-related ecological impacts. In addition to incorporating standard information (SPEI, PDSI, etc.) and statistically standardizing the definition of drought, it is also important to consider the response of ecosystems to drought. Smith [12] defined ecosystem-related extreme events as conditions in which changes in ecosystem functions and structures (such as productivity, nutrients, species composition and population) deviate from normal variability within a certain duration and area. Moreover, responses of ecosystems to extreme events include immediate and delayed effects [13], so the lagged response that happen after extreme events should also be considered.
2.2. Forest Carbon Cycle Feedbacks to Drought
Seventy-eight percent of the changes of global gross primary production (GPP) in the past three decades are related to extreme events, while drought events account for 60–90% of these climate extremes [14]. The water limitation and high air temperature directly affect the carbon cycle of forests through plant responses to stress. Resistance strategies of plants coping with drought include changes in plants physiological traits [15]. At the leaf level, plant responses to drought are reflected in the reduced stomatal conductance and changes in photosynthetic and respiratory rates, which directly affects the carbon exchange between vegetation and the atmosphere (Figure 1). Granier et al. [16] demonstrated that soil moisture is the dominant limiting factor for forest carbon exchange in the 2003 Europe drought event because the GPP and ecosystem respiration (Re) both dropped sharply when the relative extractable soil water was lower than about 0.4. Furthermore, the ecosystem feedbacks may aggravate the severity of drought because: (i) The evaporative cooling effect is reduced due to stomatal closure, so the high leaf temperature may cause heat stress; (ii) high-light stress aggravated by drought leads to the occurrence of photoinhibition or even photodamage; (iii) high temperature, high radiation and rising vapor pressure deficit (VPD) exacerbate drought by higher evapotranspiration [17].
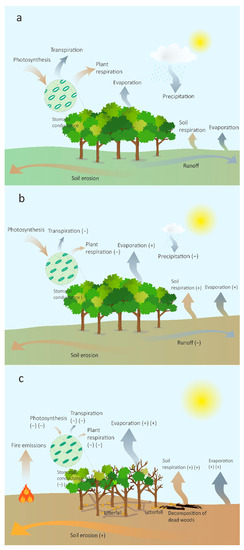
Figure 1.
Forest carbon and water fluxes change as the intensity of drought increases. (a) Before drought events. (b) Moderate drought without tree mortality. (c) Severe drought with large biomass loss. Orange arrows show the carbon flux; blue arrows show the water flux. The magnitude of fluxes is indicated by the size of arrows. Numbers of (+) and (−) indicate that the degree of fluxes increase and decrease, respectively. Plant respiration includes leaf and stem respiration; soil respiration includes autotrophic soil respiration (roots) and heterotrophic respiration (litterfall, microbial community, soil organic matter, etc.).
Forest carbon uptake and sequestration can rapidly decrease after widespread tree mortality [18], and the impacts of the decline in forest net primary productivity (NPP) could last for decades. Moreover, the decomposition process of the remnant biomass and underground dead roots releases stored carbon back to the atmosphere (Figure 1c), which can transform forests from carbon sinks into sources. Tree mortality is usually induced by the combination of multiple driving factors such as climate stress, diseases related to pests and pathogens, the stand life histories and fires [19]. Drought may serve as an inducer of the death of trees that are already under stress. The complexity of these drought-induced death mechanisms of trees makes the monitoring of tree mortality challenging. McDowell et al. [20] proposed three hypotheses about the mechanisms of drought-induced plant die-off: (i) Hydraulic failure; (ii) carbon starvation, where water limitation-induced stomatal closure blocks carbon uptake; and (iii) biotic attack, where changes in populations of pests and pathogens under continuous warming lead to excessive tree damage and mortality. Hydraulic failure is considered to be the main factor leading to plant mortality related to drought [21,22]. The hydraulic failure hypothesis states that low soil water potential and soil hydraulic conductivity lead to an increase in the vapor pressure difference between leaves and atmosphere and the rising demand for evaporation. The xylem conduits and rhizosphere are filled with air so the plant hydraulic system cannot transport water from the rhizosphere to leaves resulting in plant tissues desiccation and death [20]. Drought-induced tree mortality directly influences the land-atmosphere interaction because of the loss of vegetation function of exchanging water, energy and carbon.
Literature reporting forest carbon cycle responses to drought events in the last two decades is summarized in Table 1. The main causes of forest carbon budget variations related to drought are summarized below. First, increased tree mortality induced by drought explains large forest carbon storage loss in North America [17,23,24,25] and the Amazon [26,27,28]. As an example, in the 2010 Amazon drought event, an estimated 2.2 Pg C storage was impacted [27] with a wider impact area than the 2005 event (1.6 Pg C [26]). This impact was mainly derived from the temporary suspension of biomass growth and tree mortality, which reversed the forest from a net carbon sink to source [27].
Second, studies from North America and Europe suggest drought duration and timing are crucial factors that affect the dynamics of forest carbon balance between gross primary productivity and ecosystem respiration [29,30,31]. For example, the increase of forest carbon uptake in a warm spring is eliminated by subsequent summer drought [29,30,31,32], and early warming-induced consumption of soil moisture may intensify the water stress and heating in summer [30]. In addition, the sensitivity of carbon flux to warming and drought varies among vegetation types [31,33]. Alaskan deciduous forests showed a much higher net carbon uptake during the growing season than evergreen forest because of leaf area increase in warm spring, which enlarged the difference of response to summer drought between deciduous forests and evergreen forests [33]. This result indicates a bigger contribution of CO2 uptake by deciduous forests in the northern hemisphere.
Third, the response of ecosystem respiration is expected to contribute to the uncertainty of the net CO2 exchange, because if Re and GPP decrease with a similar magnitude, the net primary productivity is expected to be unaffected by drought. The magnitude and direction of Re variation depend on duration and severity of drought and regional characteristics (such as soil moisture and vegetation types). Von Buttlar et al. [34] global study suggests that the direction of ecosystem respiration change depends on whether heat stress coincides with drought because heat and drought have the opposite impacts on Re. Re will keep stable if a warming-induced increase in Re is offset by water limitation. For example, ecosystem respiration in Alaskan forests increased during the 2004 summer drought because warming-caused increases in respiration exceeded the effects of limitations such as soil moisture and oxygen content [33]. Moreover, the European hot drought in 2003 led to a reduction in ecosystem respiration (autotrophic and heterotrophic respiration decreased) [16,35,36]. Ciais et al. [35] suggest that less carbon assimilation led to plant respiration decrease, and the negative effect of higher water stress on heterotrophic respiration exceed the positive effects of warming on it. Similar results were found in Lindroth et al. [37], they suggest that most forests in their study experienced a decline of net ecosystem productivity (decrease both in GPP and Re) in the 2018 summer drought across Northern Europe, and the forest carbon balance change was largely explained by decreased heterotrophic respiration and lack of precipitation.

Table 1.
Summary of forest carbon cycle responses to drought events in recent two decades.
Table 1.
Summary of forest carbon cycle responses to drought events in recent two decades.
| Location | Period | Forest Carbon Variations | Causes | Reference |
|---|---|---|---|---|
| North American boreal forest | 20th Century |
|
| Barber et al., 2000 [38] |
| Europe | 2003 |
|
| Ciais et al., 2005 [35] Reichstein, M. et al., 2007 [36] Granier et al., 2007 [16] |
| Canada’s boreal forests | 2000–2003 |
|
| Kljun et al., 2006 [29] |
| Interior Alaska | 2002–2004 |
|
| Welp et al., 2007 [33] |
| Amazon | 2005 |
|
| Phillips et al., 2009 [26] |
| Amazon | 2005, 2010 |
|
| Lewis et al., 2011 [27] |
| Aspen forests in North America | 2009–2011 |
|
| Huang and Anderegg, 2011 [17] |
| Canada’s boreal forests | 1965–2005 |
|
| Ma et al., 2012 [23] |
| Congo rainforest greenness | 2000–2012 |
|
| Zhou et al., 2014 [39] |
| Amazon | 1985–2010 |
|
| Brienen et al., 2015 [28] |
| United States | 2012 |
|
| Wolf et al., 2016 [30] |
| Texas | 2011 |
|
| Schwantes et al., 2017 [24] |
| Global | Site-dependent |
|
| von Buttlar et al., 2018 [34] |
| California | 2001–2015 |
|
| Sleeter et al., 2019 [25] |
| North American forests | Site-dependent |
|
| Xu et al., 2020 [31] |
| Northern Europe | Dry summer of 2018 |
|
| Lindroth et al., 2020 [37] |
| Site-dependent | Site-dependent |
|
| Zhang and Yuan, 2020 [40] |
| Europe | 2018 |
|
| Gourlez de la Motte et al., 2020 [41] |
| Europe | 2018 |
|
| Graf et al., 2020 [42] |
| Europe | 2018 |
|
| Smith et al., 2020 [43] |
3. Elevated CO2 Effect on Plants and Ecosystem
Increasing global CO2 emissions positively stimulate the terrestrial gross primary productivity. Previous research on plant response to elevated CO2 has been developed across a range of scales, from leaf-level physiological experiments to satellite observations and the modelling of ecosystem response. We discuss CO2 fertilization effects from the perspectives of plant physiology to ecosystem response below.
3.1. How Plants Respond to Elevated CO2
Enriched atmospheric CO2 concentration interferes with plant physiological processes by reducing plant stomatal conductance, increasing water use efficiency and simultaneously enhancing photosynthesis rate and light use efficiency [4]. However, the CO2 fertilization effect is usually provisional since plants often acclimate to long-term elevated atmospheric CO2 with lower photosynthetic capacity. Furthermore, CO2 fertilization effects on plants may also be limited by other nutrients such as nitrogen (N) and phosphorus (P) [44].
3.1.1. Stomatal Conductance
The stomatal aperture is determined by the turgor pressure of the guard cell [45]. CO2 enrichment stimulates the guard cell by changing the ion channel activities or the ion composition inside and outside the cell membrane [46]. The response of stomata to atmospheric CO2 concentration varies among species, mainly owing to genetic traits and stomatal acclimation to humidity [4]. A reduced stomatal conductance () under the enriched CO2 explains the decrease in leaf transpiration and the increase in water use efficiency, but it does not seem to limit photosynthesis. Whether reduced restricts photosynthesis is indicated by the ratio of intercellular to atmospheric CO2 concentration (Ci/Ca). The decrease in stomatal conductance appears to limit plant photosynthesis if the value of Ci/Ca in elevated CO2 is less than that in normal ambient CO2. However, Drake et al. [4] examined experiments with different species and found that there is no significant difference in Ci/Ca between plants grown in ambient CO2 and elevated CO2, which proves that the reduced does not inhibit plant photosynthesis but enhances the WUE of plants.
3.1.2. Photosynthesis
The mechanism of plant photosynthesis stimulation by elevated CO2 is mainly driven by the activity of Ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco). Rubisco catalyzes the carboxylation reaction to fix CO2 into 3-phosphoglyceric acid (3PGA) [47]. Since Rubisco has a low affinity for CO2 carboxylation and is not saturated at current CO2 concentration, elevated CO2 can stimulate and promote the carboxylation reaction [4]. However, Rubisco can also catalyze the oxygenation reaction of Ribulose-1,5-bisphosphate (RuBP) and O2, that is, the first step of photorespiration that inhibits photosynthesis by consuming light energy and releasing assimilated carbon [45]. The balance of carboxylation and oxygenation reactions depends on the ratio of O2 to CO2 concentration of the site of carboxylation. Elevated CO2 stimulates carboxylation reactions and inhibits oxygenation reaction, promoting the process of photosynthesis. However, the affinity of Rubisco for CO2 and the solubility of CO2 (relative to O2) decrease with increasing temperature, thus the increase in temperature will promote photorespiration [48]. The stimulating effect of elevated CO2 on photosynthesis increases with the rising temperature, and CO2 fertilization has a stronger effect with high temperature.
Compared with C3 plants, elevated CO2 has no direct effect on the photosynthesis of C4 plants, because C4 plants have a mechanism which can avoid photorespiration. In FACE experiments, the response of C3 plants to elevated CO2 is three times that of C4 plants [44]. Recent studies [49,50] argue that C4 plants benefit from elevated CO2 indirectly by the interaction of decreased and water stress, that is, the drought stress is mitigated by rising CO2-induced higher water use efficiency.
3.1.3. Photosynthetic Acclimation
The photosynthetic rate is accelerated by short-term CO2 increases, but plants may eventually adapt to elevated CO2 atmosphere in the long-term. Plants grown in long-term elevated CO2 will experience photosynthetic down-regulation, accompanied by higher concentrations of carbohydrates, lower concentrations of soluble proteins and Rubisco than those grown in the normal ambient atmosphere [4]. The stomatal acclimation to elevated CO2 shows a significant downtrend in stomatal conductance, which is also associated with photosynthetic acclimation [45,51]. Nevertheless, the decline in is not considered the dominant reason for the down-regulation of photosynthesis because there is no obvious difference in Ci/Ca between plants grown in elevated and ambient CO2 [4,44]. The mechanism of acclimation of photosynthesis and stomata behavior is not completely understood, but there are several common possibilities. First, plants are unable to use extra carbohydrates produced by CO2-stimulated photosynthesis, which then trigger the feedback mechanism that inhibits carbohydrate production at the source [4,45]. Second, the content and activity of Rubisco decrease under elevated CO2 [4,52]. While there is no widespread agreement that photosynthesis will acclimate to elevated CO2, it is hasty to consider photosynthetic acclimation as the eventual result of the plant response to elevated CO2, especially at ecosystem scale.
3.1.4. Nutrient Availability
Rubisco requires a large amount of N investment and the activity of Rubisco play a key role in determining elevated CO2 effects on photosynthesis rate. Therefore, the strength of CO2 fertilization will be greatly reduced when other nutrients such as N are limited. There have been many experiments and studies supporting the hypothesis that CO2 fertilization is limited by N availability [53,54,55]. When the sink strength of plants is limited due to N restriction, CO2-stimulated excess photosynthate is more likely to trigger the feedback mechanism of plants that reduces products of photosynthesis from the source, thus leading to an earlier occurrence of photosynthetic acclimation [56]. The process of P affecting forest carbon uptake is different from that of N. Ellsworth et al. [57] provide the evidence from FACE experiments that P potentially limits the effect of CO2 fertilization, although their experiment is limited to forests consisting of single species. Terrer et al.’s [58] data analysis of 138 elevated CO2 experiments shows that the strength of elevated CO2 effect on 25% of global vegetation is primary constrained by P availability.
3.2. Ecosystem Responses to Elevated CO2
Plant physiological response to elevated CO2 at the leaf level has been supported by short-term individual experiments, however, extrapolation of these conclusions from individuals to ecosystems requires multiple data across scales. Over the past decades, research of plant responses to elevated CO2 scaled up and increased in complexity. Large-scale open-top chambers (OTC) and FACE experiments with an open-air condition reflect plant responses to elevated CO2 at the field level and extend the timescale of research, however, the incorrect data, experimental deficiencies (such as the unnatural increase of CO2) and unsatisfactory future prediction needs still challenge the understanding of global-scale forest responses to climate change.
Satellite observations reveal the change in global vegetation on a scale of decades and there is a documented upward trend on vegetation cover in the northern hemisphere over the past two decades [59]. Recent evidence [60,61,62,63] indicates that in recent decades, this trend is probably attributed to a combined effect of the elevated CO2, nutrients, water supply, solar radiation and socio-economic factors (such as land use change and reforestation). For example, according to Nemani et al. [60], global land NPP experienced a climate-induced increase between 1982 and 1999, which is probably due to solar radiation increase rather than CO2 fertilization in the Amazon rainforest (accounting for 42% of total global NPP increase). Hickler et al. [64] analyze the drivers of the Sahel greening trend through ecosystem modelling and the output suggests that precipitation is the main cause of increased vegetation cover while CO2 has only a small contribution. However, CO2 fertilization is considered more obvious in areas where water is the primary limitation for vegetation growth because the increased WUE under elevated CO2 mitigates water stress [5]. Donohue et al. [5] indicate that the prediction of CO2-stimulated increase in maximum foliage cover is consistent with satellite observations, which suggests that CO2 does play a key role in warm and arid area greening. Ukkola et al. [65] showed consistent results with Donohue et al., and they indicated that the WUE of vegetation in Australia increased over the past decades due to CO2 fertilization. A new method, which scales up from leaf and canopy level, was developed by Ueyama et al. [66] to isolate elevated CO2 effects from other effects and quantify the magnitude of vegetation response globally, and their results supported the effects of CO2 enrichment on GPP increase and transpiration decrease.
Process-based models that involve mechanical process descriptions are feasibly to isolate global forest response to CO2 fertilization from other drivers on a longer time scale (decades). Piao et al.’s [67] modelling studies based on ORCHIDEE suggest that 49% of the increase of leaf area index (LAI) in growing season in the northern hemisphere is contributed by atmospheric CO2 (temperature and precipitation account for 31% and 13%, respectively). Ten carbon cycle models used for the IPCC were evaluated through statistical functions by Piao et al. [68] and their results show that every increase of 100 ppm CO2 concentration contributes to 5–20% simulated NPP increase, which slightly exceeds the results from FACE experiments (13%). However, Wang et al. [69] indicated a decline in CO2 fertilization effects from 1982 to 2015 based on observation datasets, which related to increased nutrient (N, P) limitations and water limitation. Their results suggest an overestimation of ecosystem response to elevated CO2 by carbon cycle models, possibly associated to constraint factors, which restrict the ability of ecosystem responses, such as nutrient limitation, photosynthetic acclimation and soil moisture deficit.
4. Interaction of Drought and CO2 Fertilization
Plant responses to elevated CO2 and warming is reflected in the increase of photosynthetic rates and water use efficiency. Recent studies [49,50,70,71] suggest that the CO2 fertilization effect on WUE partly eliminates water limitation during drought events, however, few reports have given explicit quantitative magnitude of the WUE effect. The extent to which extreme events such as drought that negatively affect the ability of vegetation to sequester carbon, and the degree to which the impact of drought can be offset by rising CO2 remains unresolved and debatable. The ecosystem categories and drought intensity and duration are considered to determine the effects of higher water use efficiency on drought resistance. Figure 1 shows the fluxes of carbon and water under elevated CO2 in different levels of drought intensity, exhibiting the responses of forests to diverse droughts.
Reduced stomatal conductance under elevated CO2 may be able to limit water consumption at leaf level (Figure 1b), but the increased evapotranspirating leaf area attributed to rising biomass stimulated by elevated CO2 may offset the effect of CO2-induced stomatal response [72]. The sensitivity coefficient of ecosystem evapotranspiration (ET) to atmospheric CO2 reflects whether biomass growth or lower stomatal conductance dominates ecosystem ET [65]. Ecosystem evapotranspiration response to elevated CO2 may lead to soil moisture and atmospheric humidity variations, which is classified as an indirect response of ecosystems to elevated CO2 [49]. This response varies greatly among ecosystems and species, such that there is not yet a consistent pattern that can describe it.
Evidence from gas exchange chamber experiments suggest that elevated CO2 benefits to water efficiency will initially help resist desiccation. This benefit will gradually disappear under continued drought and heat, as VPD-induced stomatal closure will contribute to loss of CO2 assimilation and increased respiration with leaf temperature leading to CO2 loss (Figure 1c) [6]. The increase in air temperature and desiccation may reverse the CO2-induced WUE increase. Reichstein et al. [73] suggest that the potential possibilities of stomatal closure led to ecosystem water use efficiency decreasing during drought. Although warming and elevated CO2 are expected to benefit plant growth, warming-induced water stress will restrict the positive effects of CO2 fertilization on photosynthesis [70]. Yuan et al. [74] indicate that the terrestrial GPP in the late 1990s experienced a continuous large-scale decline, while the vapor pressure deficit increased sharply during the same period, which offset the effect of CO2 fertilization. Silva et al. [70] analyzed temperate and boreal forests in Ontario, Canada, by isotopic and dendrochronological methods, and showed that rising CO2 did increase the water use efficiency, but forest tree growth experienced a widespread decline due to water limitation induced by warming, which suggests that the increase in WUE cannot offset the growth decline under warm drought. A positive case that CO2 fertilization and warming reverses the negative effect of water limitation was found by Morgan et al. [50], who conducted FACE experiments in a semi-arid grassland with C4 grasses. However, their experiments only suggest the effective response of WUE to resist desiccation in grassland and did not cover other vegetation types. Furthermore, drought-induced tree mortality is an essential driver of forest carbon loss. Duan et al. [75] indicated that high temperature accelerates the process of plant hydraulic failure and death and the mitigative effect of elevated CO2 is negligible, but their study is limited to tree seedling mortality.
Therefore, the combined effects of drought and CO2 fertilization depend on the forest’s ability to resist water limitation, which leads to whether the biomass growth is stimulated by atmospheric CO2 or lost because of drought-induced tree mortality [18]. There is a concept of the “tipping point” for tropical forests, which indicates that forests will lose their storage carbon sharply when the global daily maximum temperature over the year increases to 32.3 °C, especially in drier forests where water is the primary limitation [18].
5. Conclusions and Outlook
Although we have recognized that drought events and CO2 fertilization have critical impacts on the carbon cycle of forests, it is still challenging to figure out the complex, comprehensive effects of climate change on vegetation.
We reviewed the current understanding of vegetation response to drought and elevated CO2 from leaf to ecosystem scales. In conclusion, plants initiate drought resistance strategies in arid and warm environments, which likely change the carbon flux of forests. Extreme drought massively killing trees is one of the main reasons for forest ecosystem carbon loss. Elevated CO2 favoring photosynthesis and water use efficiency of plants in a short period of time has been confirmed by a large number of experimental studies, but the effect of CO2 fertilization is limited by other constraint factors such as light, water and nutrients (N, P). Additionally, on a long-term scale, plants have the possibility of acclimating to warming and elevated CO2 with a lower net photosynthetic assimilation. The underlying mechanism has not been explained clearly. There is great instability and unpredictability in the physiological mechanisms of vegetation adaptation and mortality under drought [76]. This instability stems from the drought resistance of trees and elevated CO2 effects on WUE. However, the problem of how far CO2 fertilization can offset the effects of drought has not been resolved. Whether and to what extent the improved water use efficiency by CO2 fertilization can withstand drought damage remains unknown.
Understanding the interaction effects of rising CO2 and drought is crucial for looking forward to future forest ecosystems growing in a drier and warmer atmosphere. Process-based models are important tools for predicting the future interaction of forest ecosystems and atmosphere, however, accurately extrapolating the conclusions derived from individual experiment reports to large-scale ecosystem models requires more comprehensive observational data and process descriptions. There have been studies that provide information that helps improve model simulations, for example, different vegetation type responses to drought [31,33,42] and N, P limitation [53,54,55,57,58]. Applying that information to model building may help reduce the uncertainty of models. We recommend considering the uncertainties from the following perspectives:
- Including observational data across different scales. Flux tower measurements, space-time data from large-scale long-term experiments, observation of climate change and vegetation change recorded by remote sensing.
- Different responses to drought among vegetation types and changing plant species composition of ecosystems over time.
- Consideration of constraint factors. Restrictions of nutritional elements (N, P), water and light on CO2 fertilization.
- Effect of photosynthetic acclimation (down-regulation of photosynthetic capacity).
- Disturbances. The complexity of the multiple disturbance interactions associated with drought (fires, insects and pathogens).
Overall, a better understanding of mechanisms and scale-up methods are key issues in current research on forest strategies to combat climate change. However, it is worth noting that forests cannot infinitely resist climate change, and the containment of greenhouse gases emissions is the fundamental and necessary way of mitigating global warming and protecting forest ecosystem functions.
Author Contributions
Writing—original draft preparation, J.-L.X. and W.-Y.S.; writing—review and editing, F.Z. and W.-Y.S.; help and discussion, F.Z., Q.-L.H., Y.-X.Y., X.H. and W.-Y.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of China (Nos. 41975114 and 41830648).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bonan, G. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef]
- Canadell, J.G.; Raupach, M.R. Managing forests for climate change mitigation. Science 2008, 320, 1456–1457. [Google Scholar] [CrossRef] [PubMed]
- Pregitzer, K.S.; Euskirchen, E.S. Carbon cycling and storage in world forests: Biome patterns related to forest age. Glob. Chang. Biol. 2004, 10, 2052–2077. [Google Scholar] [CrossRef]
- Drake, B.G.; Gonzàlez-Meler, M.A.; Long, S.P. More efficient plants: A consequence of rising atmospheric CO2? Annu. Rev. Plant Biol. 1997, 48, 609–639. [Google Scholar] [CrossRef]
- Donohue, R.J.; Roderick, M.L.; McVicar, T.R.; Farquhar, G.D. Impact of CO2 fertilization on maximum foliage cover across the globe’s warm, arid environments. Geophys. Res. Lett. 2013, 40, 3031–3035. [Google Scholar] [CrossRef]
- Birami, B.; Nagele, T.; Gattmann, M.; Preisler, Y.; Gast, A.; Arneth, A.; Ruehr, N.K. Hot drought reduces the effects of elevated CO2 on tree water-use efficiency and carbon metabolism. New Phytol. 2020, 226, 1607–1621. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Redmond, K.T. The depiction of drought: A commentary. Bull. Am. Meteorol. Soc. 2002, 83, 1143–1148. [Google Scholar] [CrossRef]
- Ault, T.R. On the essentials of drought in a changing climate. Science 2020, 368, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Slette, I.J.; Post, A.K.; Awad, M.; Even, T.; Punzalan, A.; Williams, S.; Smith, M.D.; Knapp, A.K. How ecologists define drought, and why we should do better. Glob. Chang. Biol. 2019, 25, 3193–3200. [Google Scholar] [CrossRef]
- Eslamian, S.; Ostad-Ali-Askari, K.; Singh, V.P. A review of drought indices. Int. J. Constr. Res. Civ. Eng. 2017, 3, 48–66. [Google Scholar]
- Smith, M.D. An ecological perspective on extreme climatic events: A synthetic definition and framework to guide future research. J. Ecol. 2011, 99, 656–663. [Google Scholar] [CrossRef]
- Reichstein, M.; Bahn, M.; Ciais, P.; Frank, D.; Mahecha, M.D.; Seneviratne, S.I.; Zscheischler, J.; Beer, C.; Buchmann, N.; Frank, D.C.; et al. Climate extremes and the carbon cycle. Nature 2013, 500, 287–295. [Google Scholar] [CrossRef]
- Zscheischler, J.; Mahecha, M.D.; Von Buttlar, J.; Harmeling, S.; Jung, M.; Rammig, A.; Randerson, J.T.; Schölkopf, B.; Seneviratne, S.I.; Tomelleri, E.; et al. A few extreme events dominate global interannual variability in gross primary production. Environ. Res. Lett. 2014, 9, 035001. [Google Scholar] [CrossRef]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Granier, A.; Reichstein, M.; Bréda, N.; Janssens, I.A.; Falge, E.; Ciais, P.; Grünwald, T.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; et al. Evidence for soil water control on carbon and water dynamics in European forests during the extremely dry year: 2003. Agric. For. Meteorol. 2007, 143, 123–145. [Google Scholar] [CrossRef]
- Huang, C.Y.; Anderegg, W.R.L. Large drought-induced aboveground live biomass losses in southern Rocky Mountain aspen forests. Glob. Chang. Biol. 2011, 18, 1016–1027. [Google Scholar] [CrossRef]
- Pennisi, E. Tropical forests store carbon despite warming. Science 2020, 368, 813. [Google Scholar] [CrossRef] [PubMed]
- Anderegg, W.R.L.; Trugman, A.T.; Badgley, G.; Anderson, C.M.; Bartuska, A.; Ciais, P.; Cullenward, D.; Field, C.B.; Freeman, J.; Goetz, S.J.; et al. Climate-driven risks to the climate mitigation potential of forests. Science 2020, 368. [Google Scholar] [CrossRef]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Choat, B. Predicting thresholds of drought-induced mortality in woody plant species. Tree Physiol. 2013, 33, 669–671. [Google Scholar] [CrossRef]
- Anderegg, W.R.; Berry, J.A.; Smith, D.D.; Sperry, J.S.; Anderegg, L.D.; Field, C.B. The roles of hydraulic and carbon stress in a widespread climate-induced forest die-off. Proc. Natl. Acad. Sci. USA 2012, 109, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Peng, C.; Zhu, Q.; Chen, H.; Yu, G.; Li, W.; Zhou, X.; Wang, W.; Zhang, W. Regional drought-induced reduction in the biomass carbon sink of Canada’s boreal forests. Proc. Natl. Acad. Sci. USA 2012, 109, 2423–2427. [Google Scholar] [CrossRef]
- Schwantes, A.M.; Swenson, J.J.; Gonzalez-Roglich, M.; Johnson, D.M.; Domec, J.C.; Jackson, R.B. Measuring canopy loss and climatic thresholds from an extreme drought along a fivefold precipitation gradient across Texas. Glob. Chang. Biol. 2017, 23, 5120–5135. [Google Scholar] [CrossRef] [PubMed]
- Sleeter, B.M.; Marvin, D.C.; Cameron, D.R.; Selmants, P.C.; Westerling, A.L.; Kreitler, J.; Daniel, C.J.; Liu, J.; Wilson, T.S. Effects of 21st-century climate, land use, and disturbances on ecosystem carbon balance in California. Glob. Chang. Biol. 2019, 25. [Google Scholar] [CrossRef]
- Phillips, O.L.; Aragão, L.E.; Lewis, S.L.; Fisher, J.B.; Lloyd, J.; López-González, G.; Malhi, Y.; Monteagudo, A.; Peacock, J.; Quesada, C.A. Drought sensitivity of the Amazon rainforest. Science 2009, 323, 1344–1347. [Google Scholar] [CrossRef]
- Lewis, S.L.; Brando, P.M.; Phillips, O.L.; Van der Heijden, G.M.; Nepstad, D. The 2010 amazon drought. Science 2011, 331, 554. [Google Scholar] [CrossRef]
- Brienen, R.J.; Phillips, O.L.; Feldpausch, T.R.; Gloor, E.; Baker, T.R.; Lloyd, J.; Lopez-Gonzalez, G.; Monteagudo-Mendoza, A.; Malhi, Y.; Lewis, S.L.; et al. Long-term decline of the Amazon carbon sink. Nature 2015, 519, 344–348. [Google Scholar] [CrossRef]
- Kljun, N.; Black, T.A.; Griffis, T.J.; Barr, A.G.; Gaumont-Guay, D.; Morgenstern, K.; McCaughey, J.H.; Nesic, Z. Response of net ecosystem productivity of three boreal forest stands to drought. Ecosystems 2007, 10, 1039–1055. [Google Scholar] [CrossRef]
- Wolf, S.; Keenan, T.F.; Fisher, J.B.; Baldocchi, D.D.; Desai, A.R.; Richardson, A.D.; Scott, R.L.; Law, B.E.; Litvak, M.E.; Brunsell, N.A.; et al. Warm spring reduced carbon cycle impact of the 2012 US summer drought. Proc. Natl. Acad. Sci. USA 2016, 113, 5880–5885. [Google Scholar] [CrossRef]
- Xu, B.; Arain, M.A.; Black, T.A.; Law, B.E.; Pastorello, G.Z.; Chu, H.S. Seasonal variability of forest sensitivity to heat and drought stresses: A synthesis based on carbon fluxes from North American forest ecosystems. Glob. Chang. Biol. 2020, 26. [Google Scholar] [CrossRef]
- Angert, A.; Biraud, S.; Bonfils, C.; Henning, C.C.; Buermann, W.; Pinzon, J.; Tucker, C.J.; Fung, I. Drier summers cancel out the CO2 uptake enhancement induced by warmer springs. Proc. Natl. Acad. Sci. USA 2005, 102, 10823–10827. [Google Scholar] [CrossRef] [PubMed]
- Welp, L.R.; Randerson, J.T.; Liu, H.P. The sensitivity of carbon fluxes to spring warming and summer drought depends on plant functional type in boreal forest ecosystems. Agric. For. Meteorol. 2007, 147, 172–185. [Google Scholar] [CrossRef]
- Von Buttlar, J.; Zscheischler, J.; Rammig, A.; Sippel, S.; Reichstein, M.; Knohl, A.; Jung, M.; Menzer, O.; Arain, M.A.; Buchmann, N.; et al. Impacts of droughts and extreme-temperature events on gross primary production and ecosystem respiration: A systematic assessment across ecosystems and climate zones. Biogeosciences 2018, 15. [Google Scholar] [CrossRef]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Reichstein, M.; Ciais, P.; Papale, D.; Valentini, R.; Running, S.; Viovy, N.; Cramer, W.; Granier, A.; Ogee, J.; Allard, V.; et al. Reduction of ecosystem productivity and respiration during the European summer 2003 climate anomaly: A joint flux tower, remote sensing and modelling analysis. Glob. Chang. Biol. 2007, 13, 634–651. [Google Scholar] [CrossRef]
- Lindroth, A.; Holst, J.; Linderson, M.-L.; Aurela, M.; Biermann, T.; Heliasz, M.; Chi, J.; Ibrom, A.; Kolari, P.; Klemedtsson, L.; et al. Effects of drought and meteorological forcing on carbon and water fluxes in Nordic forests during the dry summer of 2018. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375. [Google Scholar] [CrossRef] [PubMed]
- Barber, V.A.; Juday, G.P.; Finney, B. Reduced growth of Alaskan white spruce in the twentieth century from temperature-induced drought stress. Nature 2000, 405, 668–673. [Google Scholar] [CrossRef]
- Zhou, L.; Tian, Y.; Myneni, R.B.; Ciais, P.; Saatchi, S.; Liu, Y.Y.; Piao, S.; Chen, H.; Vermote, E.F.; Song, C.; et al. Widespread decline of Congo rainforest greenness in the past decade. Nature 2014, 509, 86–90. [Google Scholar] [CrossRef]
- Zhang, M.; Yuan, X. Rapid reduction in ecosystem productivity caused by flash droughts based on decade-long FLUXNET observations. Hydrol. Earth Syst. Sci. 2020, 24, 5579–5593. [Google Scholar] [CrossRef]
- Gourlez de la Motte, L.; Beauclaire, Q.; Heinesch, B.; Cuntz, M.; Foltynova, L.; Sigut, L.; Kowalska, N.; Manca, G.; Ballarin, I.G.; Vincke, C.; et al. Non-stomatal processes reduce gross primary productivity in temperate forest ecosystems during severe edaphic drought. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375. [Google Scholar] [CrossRef] [PubMed]
- Graf, A.; Klosterhalfen, A.; Arriga, N.; Bernhofer, C.; Bogena, H.; Bornet, F.; Brueggemann, N.; Bruemmer, C.; Buchmann, N.; Chi, J.; et al. Altered energy partitioning across terrestrial ecosystems in the European drought year 2018. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375. [Google Scholar] [CrossRef]
- Smith, N.E.; Kooijmans, L.M.J.; Koren, G.; Van Schaik, E.; Van der Woude, A.M.; Wanders, N.; Ramonet, M.; Xueref-Remy, I.; Siebicke, L.; Manca, G.; et al. Spring enhancement and summer reduction in carbon uptake during the 2018 drought in northwestern Europe. Philos. Trans. R. Soc. B Biol. Sci 2020, 375. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Long, S.P. What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol. 2005, 165, 351–371. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef]
- Assmann, S.M. The cellular basis of guard cell sensing of rising CO2. Plant Cell Environ. 1999, 22, 629–637. [Google Scholar] [CrossRef]
- Mohr, H.; Schopfer, P. The leaf as a photosynthetic system. In Plant Physiology; Springer: Berlin/Heidelberg, Germany, 1995; pp. 225–243. [Google Scholar]
- Long, S.P. Modification of the response of photosynthetic productivity to rising temperature by atmospheric CO2 concentrations: Has its importance been underestimated? Plant Cell Environ. 1991, 14, 729–739. [Google Scholar] [CrossRef]
- Morgan, J.A.; Pataki, D.E.; Korner, C.; Clark, H.; Del Grosso, S.J.; Grunzweig, J.M.; Knapp, A.K.; Mosier, A.R.; Newton, P.C.; Niklaus, P.A.; et al. Water relations in grassland and desert ecosystems exposed to elevated atmospheric CO2. Oecologia 2004, 140, 11–25. [Google Scholar] [CrossRef]
- Morgan, J.A.; LeCain, D.R.; Pendall, E.; Blumenthal, D.M.; Kimball, B.A.; Carrillo, Y.; Williams, D.G.; Heisler-White, J.; Dijkstra, F.A.; West, M. C4 grasses prosper as carbon dioxide eliminates desiccation in warmed semi-arid grassland. Nature 2011, 476, 202–205. [Google Scholar] [CrossRef]
- Xu, D.-Q.; Gifford, R.M.; Chow, W. Photosynthetic acclimation in pea and soybean to high atmospheric CO2 partial pressure. Plant Physiol. 1994, 106, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, J.R.; Patterson, A.E.; Rodrigues, W.P.; Campostrini, E.; Griffin, K.L. Photosynthetic acclimation to elevated CO2 combined with partial rootzone drying results in improved water use efficiency, drought tolerance and leaf carbon balance of grapevines. Environ. Exp. Bot. 2017, 134, 82–95. [Google Scholar] [CrossRef]
- Norby, R.J.; Warren, J.M.; Iversen, C.M.; Medlyn, B.E.; McMurtrie, R.E. CO2 enhancement of forest productivity constrained by limited nitrogen availability. Proc. Natl. Acad. Sci. USA 2010, 107, 19368–19373. [Google Scholar] [CrossRef]
- Reich, P.B.; Hobbie, S.E.; Lee, T.; Ellsworth, D.S.; West, J.B.; Tilman, D.; Knops, J.M.; Naeem, S.; Trost, J. Nitrogen limitation constrains sustainability of ecosystem response to CO2. Nature 2006, 440, 922–925. [Google Scholar] [CrossRef]
- Reich, P.B.; Hobbie, S.E.; Lee, T.D. Plant growth enhancement by elevated CO2 eliminated by joint water and nitrogen limitation. Nat. Geosci. 2014, 7, 920–924. [Google Scholar] [CrossRef]
- Rogers, A.; Fischer, B.U.; Bryant, J.; Frehner, M.; Blum, H.; Raines, C.A.; Long, S.P. Acclimation of Photosynthesis to Elevated CO2 under Low-Nitrogen Nutrition Is Affected by the Capacity for Assimilate Utilization. Perennial Ryegrass under Free-Air CO2 Enrichment. Plant Physiol. 1998, 118, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Ellsworth, D.S.; Anderson, I.C.; Crous, K.Y.; Cooke, J.; Drake, J.E.; Gherlenda, A.N.; Gimeno, T.E.; Macdonald, C.A.; Medlyn, B.E.; Powell, J.R.; et al. Elevated CO2 does not increase eucalypt forest productivity on a low-phosphorus soil. Nat. Clim. Chang. 2017, 7, 279–282. [Google Scholar] [CrossRef]
- Terrer, C.; Jackson, R.B.; Prentice, I.C.; Keenan, T.F.; Kaiser, C.; Vicca, S.; Fisher, J.B.; Reich, P.B.; Stocker, B.D.; Hungate, B.A.; et al. Nitrogen and phosphorus constrain the CO2 fertilization of global plant biomass. Nat. Clim. Chang. 2019, 9, 684–689. [Google Scholar] [CrossRef]
- Zhou, L.; Tucker, C.J.; Kaufmann, R.K.; Slayback, D.; Shabanov, N.V.; Myneni, R.B. Variations in northern vegetation activity inferred from satellite data of vegetation index during 1981 to 1999. J. Geophys. Res. Atmos. 2001, 106, 20069–20083. [Google Scholar] [CrossRef]
- Nemani, R.R.; Keeling, C.D.; Hashimoto, H.; Jolly, W.M.; Piper, S.C.; Tucker, C.J.; Myneni, R.B.; Running, S.W. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 2003, 300, 1560–1563. [Google Scholar] [CrossRef] [PubMed]
- Donohue, R.J.; McVicar, T.R.; Roderick, M.L. Climate-related trends in Australian vegetation cover as inferred from satellite observations, 1981–2006. Glob. Chang. Biol. 2009, 15, 1025–1039. [Google Scholar] [CrossRef]
- Fensholt, R.; Langanke, T.; Rasmussen, K.; Reenberg, A.; Prince, S.D.; Tucker, C.; Scholes, R.J.; Le, Q.B.; Bondeau, A.; Eastman, R.; et al. Greenness in semi-arid areas across the globe 1981–2007—An Earth Observing Satellite based analysis of trends and drivers. Remote Sens. Environ. 2012, 121, 144–158. [Google Scholar] [CrossRef]
- Keenan, T.F.; Prentice, I.C.; Canadell, J.G.; Williams, C.A.; Wang, H.; Raupach, M.; Collatz, G.J. Recent pause in the growth rate of atmospheric CO2 due to enhanced terrestrial carbon uptake. Nat. Commun. 2016, 7, 13428. [Google Scholar] [CrossRef]
- Hickler, T.; Eklundh, L.; Seaquist, J.W.; Smith, B.; Ardö, J.; Olsson, L.; Sykes, M.T.; Sjöström, M. Precipitation controls Sahel greening trend. Geophys. Res. Lett. 2005, 32. [Google Scholar] [CrossRef]
- Ukkola, A.M.; Prentice, I.C.; Keenan, T.F.; Van Dijk, A.I.J.M.; Viney, N.R.; Myneni, R.B.; Bi, J. Reduced streamflow in water-stressed climates consistent with CO2 effects on vegetation. Nat. Clim. Chang. 2016, 6, 75–78. [Google Scholar] [CrossRef]
- Ueyama, M.; Ichii, K.; Kobayashi, H.; Kumagai, T.O.; Beringer, J.; Merbold, L.; Euskirchen, E.S.; Hirano, T.; Marchesini, L.B.; Baldocchi, D.; et al. Inferring CO2 fertilization effect based on global monitoring land-atmosphere exchange with a theoretical model. Environ. Res. Lett. 2020, 15, 084009. [Google Scholar] [CrossRef]
- Piao, S.; Friedlingstein, P.; Ciais, P.; Zhou, L.; Chen, A. Effect of climate and CO2 changes on the greening of the Northern Hemisphere over the past two decades. Geophys. Res. Lett. 2006, 33. [Google Scholar] [CrossRef]
- Piao, S.; Sitch, S.; Ciais, P.; Friedlingstein, P.; Peylin, P.; Wang, X.; Ahlstrom, A.; Anav, A.; Canadell, J.G.; Cong, N.; et al. Evaluation of terrestrial carbon cycle models for their response to climate variability and to CO2 trends. Glob. Chang. Biol. 2013, 19. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, Y.; Ju, W.; Chen, J.M.; Ciais, P.; Cescatti, A.; Sardans, J.; Janssens, I.A.; Wu, M.; Berry, J.A.; et al. Recent global decline of CO2 fertilization effects on vegetation photosynthesis. Science 2020, 370, 1295–1300. [Google Scholar] [CrossRef]
- Silva, L.C.; Anand, M.; Leithead, M.D. Recent widespread tree growth decline despite increasing atmospheric CO2. PLoS ONE 2010, 5, e11543. [Google Scholar] [CrossRef] [PubMed]
- Sangines de Carcer, P.; Vitasse, Y.; Penuelas, J.; Jassey, V.E.J.; Buttler, A.; Signarbieux, C. Vapor-pressure deficit and extreme climatic variables limit tree growth. Glob. Chang. Biol. 2018, 24, 1108–1122. [Google Scholar] [CrossRef]
- Piao, S.; Friedlingstein, P.; Ciais, P.; De Noblet-Ducoudré, N.; Labat, D.; Zaehle, S. Changes in climate and land use have a larger direct impact than rising CO2 on global river runoff trends. Proc. Natl. Acad. Sci. USA 2007, 104, 15242–15247. [Google Scholar] [CrossRef]
- Reichstein, M.; Tenhunen, J.D.; Roupsard, O.; Ourcival, J.M.; Rambal, S.; Miglietta, F.; Peressotti, A.; Pecchiari, M.; Tirone, G.; Valentini, R. Severe drought effects on ecosystem CO2 and H2O fluxes at three Mediterranean evergreen sites: Revision of current hypotheses? Glob. Chang. Biol. 2002, 8, 999–1017. [Google Scholar] [CrossRef]
- Yuan, W.; Zheng, Y.; Piao, S.; Ciais, P.; Lombardozzi, D.; Wang, Y.; Ryu, Y.; Chen, G.; Dong, W.; Hu, Z. Increased atmospheric vapor pressure deficit reduces global vegetation growth. Sci. Adv. 2019, 5, eaax1396. [Google Scholar] [CrossRef]
- Duan, H.; Duursma, R.A.; Huang, G.; Smith, R.A.; Choat, B.; O’Grady, A.P.; Tissue, D.T. Elevated [CO2] does not ameliorate the negative effects of elevated temperature on drought-induced mortality in Eucalyptus radiata seedlings. Plant Cell Environ. 2014, 37. [Google Scholar] [CrossRef] [PubMed]
- Jump, A.S.; Ruiz-Benito, P.; Greenwood, S.; Allen, C.D.; Kitzberger, T.; Fensham, R.; Martínez-Vilalta, J.; Lloret, F. Structural overshoot of tree growth with climate variability and the global spectrum of drought-induced forest dieback. Glob. Chang. Biol. 2017, 23, 3742–3757. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).