
Comparative Analysis of MBNL1 Antibodies: Characterization of Recognition Sites and Detection of RNA Foci Colocalization


Abstract
1. Introduction
2. Materials and Methods
2.1. Plasmid Construction
2.2. Cell Culture
2.3. Transfection
2.4. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.5. Reverse Transcription-PCR (RT-PCR)
2.6. Fluorescence In Situ Hybridization (FISH)
2.7. Immunofluorescence Staining
2.8. Immunoprecipitation
3. Results
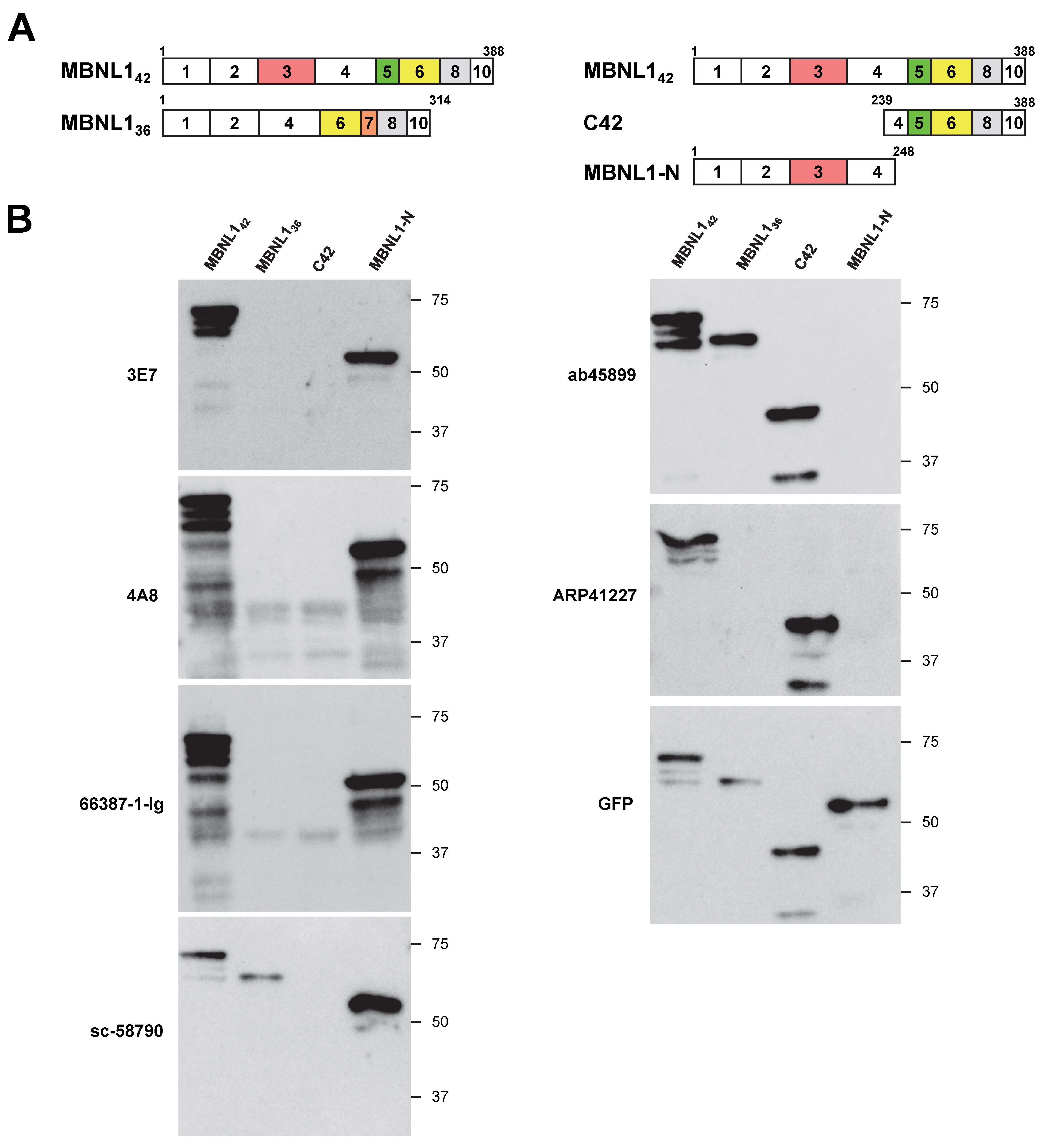
3.1. Differential Detection of MBNL1 by Various Antibodies
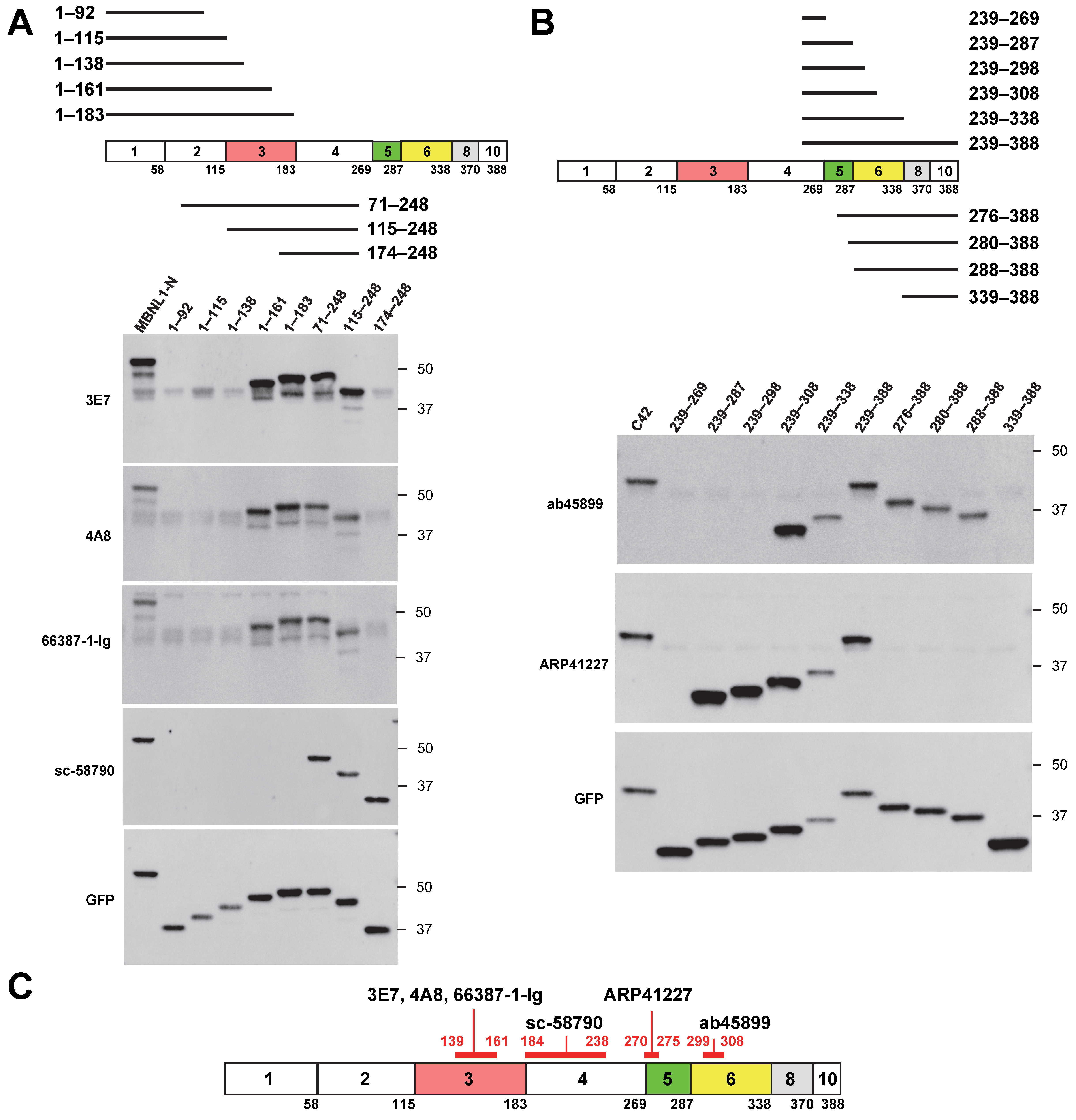
3.2. Mapping of MBNL1 Antibody Recognition Sites Using Deletion Mutants
3.3. Comparison of Antibody Reactivity to MBNL Paralogs
3.4. Endogenous Expression Patterns of MBNL1
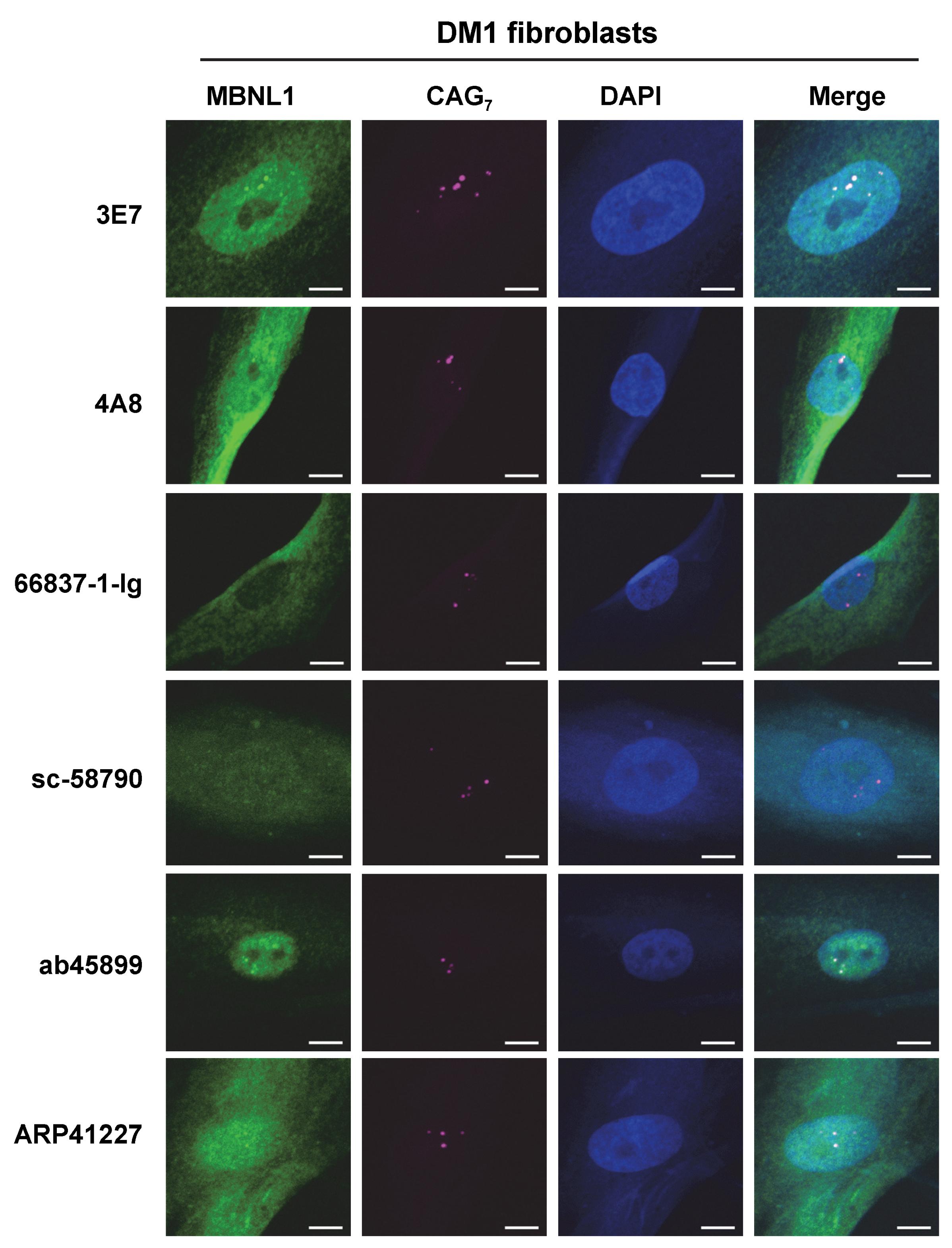
3.5. Detection of RNA Foci
3.6. Detection of MBNL1 by Immunoprecipitation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| DM | Myotonic dystrophy |
| DM1 | Myotonic dystrophy type 1 |
| DM2 | Myotonic dystrophy type 2 |
| EGFP | Enhanced green fluorescent protein |
| FISH | Fluorescence In Situ Hybridization |
| IF | Immunofluorescence |
| MBNL1 | Muscleblind-like 1 |
| ZnF | Zinc finger |
References
- Hildebrandt, R.P.; Moss, K.R.; Janusz-Kaminska, A.; Knudson, L.A.; Denes, L.T.; Saxena, T.; Boggupalli, D.P.; Li, Z.; Lin, K.; Bassell, G.J.; et al. Muscleblind-like proteins use modular domains to localize RNAs by riding kinesins and docking to membranes. Nat. Commun. 2023, 14, 3427. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.H.; Charlet, B.N.; Poulos, M.G.; Singh, G.; Swanson, M.S.; Cooper, T.A. Muscleblind proteins regulate alternative splicing. EMBO J. 2004, 23, 3103–3112. [Google Scholar] [CrossRef]
- Davis, J.; Salomonis, N.; Ghearing, N.; Lin, S.C.; Kwong, J.Q.; Mohan, A.; Swanson, M.S.; Molkentin, J.D. MBNL1-mediated regulation of differentiation RNAs promotes myofibroblast transformation and the fibrotic response. Nat. Commun. 2015, 6, 10084. [Google Scholar] [CrossRef] [PubMed]
- Masuda, A.; Andersen, H.S.; Doktor, T.K.; Okamoto, T.; Ito, M.; Andresen, B.S.; Ohno, K. CUGBP1 and MBNL1 preferentially bind to 3′ UTRs and facilitate mRNA decay. Sci. Rep. 2012, 2, 209. [Google Scholar] [CrossRef]
- Rau, F.; Freyermuth, F.; Fugier, C.; Villemin, J.P.; Fischer, M.C.; Jost, B.; Dembele, D.; Gourdon, G.; Nicole, A.; Duboc, D.; et al. Misregulation of miR-1 processing is associated with heart defects in myotonic dystrophy. Nat. Struct. Mol. Biol. 2011, 18, 840–845. [Google Scholar] [CrossRef]
- Begemann, G.; Paricio, N.; Artero, R.; Kiss, I.; Pérez-Alonso, M.; Mlodzik, M. Muscleblind, a gene required for photoreceptor differentiation in Drosophila, encodes novel nuclear Cys3His-type zinc-finger-containing proteins. Development 1997, 124, 4321–4331. [Google Scholar] [CrossRef]
- Artero, R.; Prokop, A.; Paricio, N.; Begemann, G.; Pueyo, I.; Mlodzik, M.; Perez-Alonso, M.; Baylies, M.K. The muscleblind gene participates in the organization of Z-bands and epidermal attachments of Drosophila muscles and is regulated by Dmef2. Dev. Biol. 1998, 195, 131–143. [Google Scholar] [CrossRef]
- Fardaei, M.; Rogers, M.T.; Thorpe, H.M.; Larkin, K.; Hamshere, M.G.; Harper, P.S.; Brook, J.D. Three proteins, MBNL, MBLL and MBXL, co-localize in vivo with nuclear foci of expanded-repeat transcripts in DM1 and DM2 cells. Hum. Mol. Genet. 2002, 11, 805–814. [Google Scholar] [CrossRef]
- Sznajder, Ł.J.; Swanson, M.S. Short Tandem Repeat Expansions and RNA-Mediated Pathogenesis in Myotonic Dystrophy. Int. J. Mol. Sci. 2019, 20, 3365. [Google Scholar] [CrossRef]
- Liquori, C.L.; Ricker, K.; Moseley, M.L.; Jacobsen, J.F.; Kress, W.; Naylor, S.L.; Day, J.W.; Ranum, L.P. Myotonic dystrophy type 2 caused by a CCTG expansion in intron 1 of ZNF9. Science 2001, 293, 864–867. [Google Scholar] [CrossRef]
- Mankodi, A.; Urbinati, C.R.; Yuan, Q.P.; Moxley, R.T.; Sansone, V.; Krym, M.; Henderson, D.; Schalling, M.; Swanson, M.S.; Thornton, C.A. Muscleblind localizes to nuclear foci of aberrant RNA in myotonic dystrophy types 1 and 2. Hum. Mol. Genet. 2001, 10, 2165–2170. [Google Scholar] [CrossRef] [PubMed]
- Savkur, R.S.; Philips, A.V.; Cooper, T.A. Aberrant regulation of insulin receptor alternative splicing is associated with insulin resistance in myotonic dystrophy. Nat. Genet. 2001, 29, 40–47. [Google Scholar] [CrossRef]
- Kino, Y.; Washizu, C.; Oma, Y.; Onishi, H.; Nezu, Y.; Sasagawa, N.; Nukina, N.; Ishiura, S. MBNL and CELF proteins regulate alternative splicing of the skeletal muscle chloride channel CLCN1. Nucleic Acids Res. 2009, 37, 6477–6490. [Google Scholar] [CrossRef]
- Freyermuth, F.; Rau, F.; Kokunai, Y.; Linke, T.; Sellier, C.; Nakamori, M.; Kino, Y.; Arandel, L.; Jollet, A.; Thibault, C.; et al. Splicing misregulation of SCN5A contributes to cardiac-conduction delay and heart arrhythmia in myotonic dystrophy. Nat. Commun. 2016, 7, 11067. [Google Scholar] [CrossRef] [PubMed]
- Kanadia, R.N.; Johnstone, K.A.; Mankodi, A.; Lungu, C.; Thornton, C.A.; Esson, D.; Timmers, A.M.; Hauswirth, W.W.; Swanson, M.S. A muscleblind knockout model for myotonic dystrophy. Science 2003, 302, 1978–1980. [Google Scholar] [CrossRef]
- Charizanis, K.; Lee, K.Y.; Batra, R.; Goodwin, M.; Zhang, C.; Yuan, Y.; Shiue, L.; Cline, M.; Scotti, M.M.; Xia, G.; et al. Muscleblind-like 2-mediated alternative splicing in the developing brain and dysregulation in myotonic dystrophy. Neuron 2012, 75, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.Y.; Li, M.; Manchanda, M.; Batra, R.; Charizanis, K.; Mohan, A.; Warren, S.A.; Chamberlain, C.M.; Finn, D.; Hong, H.; et al. Compound loss of muscleblind-like function in myotonic dystrophy. EMBO Mol. Med. 2013, 5, 1887–1900. [Google Scholar] [CrossRef]
- Rudnicki, D.D.; Holmes, S.E.; Lin, M.W.; Thornton, C.A.; Ross, C.A.; Margolis, R.L. Huntington’s disease--like 2 is associated with CUG repeat-containing RNA foci. Ann. Neurol. 2007, 61, 272–282. [Google Scholar] [CrossRef]
- Mootha, V.V.; Hussain, I.; Cunnusamy, K.; Graham, E.; Gong, X.; Neelam, S.; Xing, C.; Kittler, R.; Petroll, W.M. TCF4 Triplet Repeat Expansion and Nuclear RNA Foci in Fuchs’ Endothelial Corneal Dystrophy. Investig. Ophthalmol. Vis. Sci. 2015, 56, 2003–2011. [Google Scholar] [CrossRef]
- Iwahashi, C.K.; Yasui, D.H.; An, H.J.; Greco, C.M.; Tassone, F.; Nannen, K.; Babineau, B.; Lebrilla, C.B.; Hagerman, R.J.; Hagerman, P.J. Protein composition of the intranuclear inclusions of FXTAS. Brain 2006, 129, 256–271. [Google Scholar] [CrossRef]
- Kume, K.; Kurashige, T.; Muguruma, K.; Morino, H.; Tada, Y.; Kikumoto, M.; Miyamoto, T.; Akutsu, S.N.; Matsuda, Y.; Matsuura, S.; et al. CGG repeat expansion in LRP12 in amyotrophic lateral sclerosis. Am. J. Hum. Genet. 2023, 110, 1086–1097. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Shan, J.; Yu, M.; Di, L.; Xie, Z.; Zhang, W.; Lv, H.; Meng, L.; Zheng, Y.; Zhao, Y.; et al. The CGG repeat expansion in RILPL1 is associated with oculopharyngodistal myopathy type 4. Am. J. Hum. Genet. 2022, 109, 533–541. [Google Scholar] [CrossRef]
- Yu, J.; Deng, J.; Guo, X.; Shan, J.; Luan, X.; Cao, L.; Zhao, J.; Yu, M.; Zhang, W.; Lv, H.; et al. The GGC repeat expansion in NOTCH2NLC is associated with oculopharyngodistal myopathy type 3. Brain 2021, 144, 1819–1832. [Google Scholar] [CrossRef]
- Zhou, H.; Xu, J.; Pan, L. Functions of the Muscleblind-like protein family and their role in disease. Cell Commun. Signal. 2025, 23, 97. [Google Scholar] [CrossRef] [PubMed]
- Goers, E.S.; Purcell, J.; Voelker, R.B.; Gates, D.P.; Berglund, J.A. MBNL1 binds GC motifs embedded in pyrimidines to regulate alternative splicing. Nucleic Acids Res. 2010, 38, 2467–2484. [Google Scholar] [CrossRef] [PubMed]
- Konieczny, P.; Stepniak-Konieczna, E.; Taylor, K.; Sznajder, L.J.; Sobczak, K. Autoregulation of MBNL1 function by exon 1 exclusion from MBNL1 transcript. Nucleic Acids Res. 2017, 45, 1760–1775. [Google Scholar] [CrossRef]
- Tran, H.; Gourrier, N.; Lemercier-Neuillet, C.; Dhaenens, C.M.; Vautrin, A.; Fernandez-Gomez, F.J.; Arandel, L.; Carpentier, C.; Obriot, H.; Eddarkaoui, S.; et al. Analysis of exonic regions involved in nuclear localization, splicing activity, and dimerization of Muscleblind-like-1 isoforms. J. Biol. Chem. 2011, 286, 16435–16446. [Google Scholar] [CrossRef]
- Kino, Y.; Washizu, C.; Kurosawa, M.; Oma, Y.; Hattori, N.; Ishiura, S.; Nukina, N. Nuclear localization of MBNL1: Splicing-mediated autoregulation and repression of repeat-derived aberrant proteins. Hum. Mol. Genet. 2015, 24, 740–756. [Google Scholar] [CrossRef]
- Gates, D.P.; Coonrod, L.A.; Berglund, J.A. Autoregulated splicing of muscleblind-like 1 (MBNL1) Pre-mRNA. J. Biol. Chem. 2011, 286, 34224–34233. [Google Scholar] [CrossRef]
- Sznajder, Ł.J.; Michalak, M.; Taylor, K.; Cywoniuk, P.; Kabza, M.; Wojtkowiak-Szlachcic, A.; Matłoka, M.; Konieczny, P.; Sobczak, K. Mechanistic determinants of MBNL activity. Nucleic Acids Res. 2016, 44, 10326–10342. [Google Scholar] [CrossRef]
- Aoki, Y.; Yanaizu, M.; Ohki, A.; Nishimiya, K.; Kino, Y. CUG repeat RNA-dependent proteasomal degradation of MBNL1 in a cellular model of myotonic dystrophy type 1. Biochem. Biophys. Res. Commun. 2024, 733, 150729. [Google Scholar] [CrossRef] [PubMed]
- Kawada, R.; Jonouchi, T.; Kagita, A.; Sato, M.; Hotta, A.; Sakurai, H. Establishment of quantitative and consistent in vitro skeletal muscle pathological models of myotonic dystrophy type 1 using patient-derived iPSCs. Sci. Rep. 2023, 13, 94. [Google Scholar] [CrossRef] [PubMed]
- Holt, I.; Jacquemin, V.; Fardaei, M.; Sewry, C.A.; Butler-Browne, G.S.; Furling, D.; Brook, J.D.; Morris, G.E. Muscleblind-like proteins: Similarities and differences in normal and myotonic dystrophy muscle. Am. J. Pathol. 2009, 174, 216–227. [Google Scholar] [CrossRef] [PubMed]
- Yanaizu, M.; Adachi, H.; Araki, M.; Kontani, K.; Kino, Y. Translational regulation and protein-coding capacity of the 5′ untranslated region of human TREM2. Commun. Biol. 2023, 6, 616. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MBNL1 Antibody | Company | Catalog Number | Host Species | Immunogen |
|---|---|---|---|---|
| 3E7 | Abnova (Taipei, Taiwan) | H00004154-M02 | Mouse monoclonal | Full-length recombinant human MBNL1 |
| 4A8 | Santa Cruz Biotechnology (Dallas, TX, USA) | sc-136165 | Mouse monoclonal | Full-length recombinant human MBNL1 |
| MBNL1/2/3 | Santa Cruz Biotechnology | sc-58790 | Mouse monoclonal | A 371 amino acid fragment of human MBNL1 |
| 66837-1-Ig | Proteintech (Rosemont, IL, USA) | 66837-1-Ig | Mouse monoclonal | Recombinant protein (amino acids 59–400 of human MBNL143) |
| ab45899 | Abcam (Cambridge, UK) | ab45899 | Rabbit polyclonal | Synthetic peptide (amino acids 250–350 of human MBNL1) |
| ARP41227 | Aviva Systems Biology (San Diego, CA, USA) | ARP41227_P050 | Rabbit polyclonal | Synthetic peptide (amino acids 268–317) |
| MBNL1 Antibody | Recognition Site | RNA Foci Detection | Immunoprecipitation |
|---|---|---|---|
| 3E7 | 139–161 (exon 3) | Applicable | Applicable |
| 4A8 | 139–161 (exon 3) | Applicable | Applicable |
| sc-58790 | 184–238 (exon 4) | Not applicable | Not applicable |
| 66837-1-Ig | 139–161 (exon 3) | Not applicable | Applicable |
| ab45899 | 299–308 (exon 6) | Applicable | Applicable (recommended) |
| ARP41227 | 270–275 (exon 5) | Applicable | Applicable (not efficient) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aoki, Y.; Ohki, A.; Yanaizu, M.; Kino, Y. Comparative Analysis of MBNL1 Antibodies: Characterization of Recognition Sites and Detection of RNA Foci Colocalization. Genes 2025, 16, 658. https://doi.org/10.3390/genes16060658
Aoki Y, Ohki A, Yanaizu M, Kino Y. Comparative Analysis of MBNL1 Antibodies: Characterization of Recognition Sites and Detection of RNA Foci Colocalization. Genes. 2025; 16(6):658. https://doi.org/10.3390/genes16060658
Chicago/Turabian StyleAoki, Yoshitaka, Ai Ohki, Motoaki Yanaizu, and Yoshihiro Kino. 2025. "Comparative Analysis of MBNL1 Antibodies: Characterization of Recognition Sites and Detection of RNA Foci Colocalization" Genes 16, no. 6: 658. https://doi.org/10.3390/genes16060658
APA StyleAoki, Y., Ohki, A., Yanaizu, M., & Kino, Y. (2025). Comparative Analysis of MBNL1 Antibodies: Characterization of Recognition Sites and Detection of RNA Foci Colocalization. Genes, 16(6), 658. https://doi.org/10.3390/genes16060658