
Genome-Wide Analysis of the LBD Gene Family in Melon and Expression Analysis in Response to Wilt Disease Infection

Abstract
1. Introduction
2. Materials and Methods
2.1. Identification of the Melon LBD Gene Family
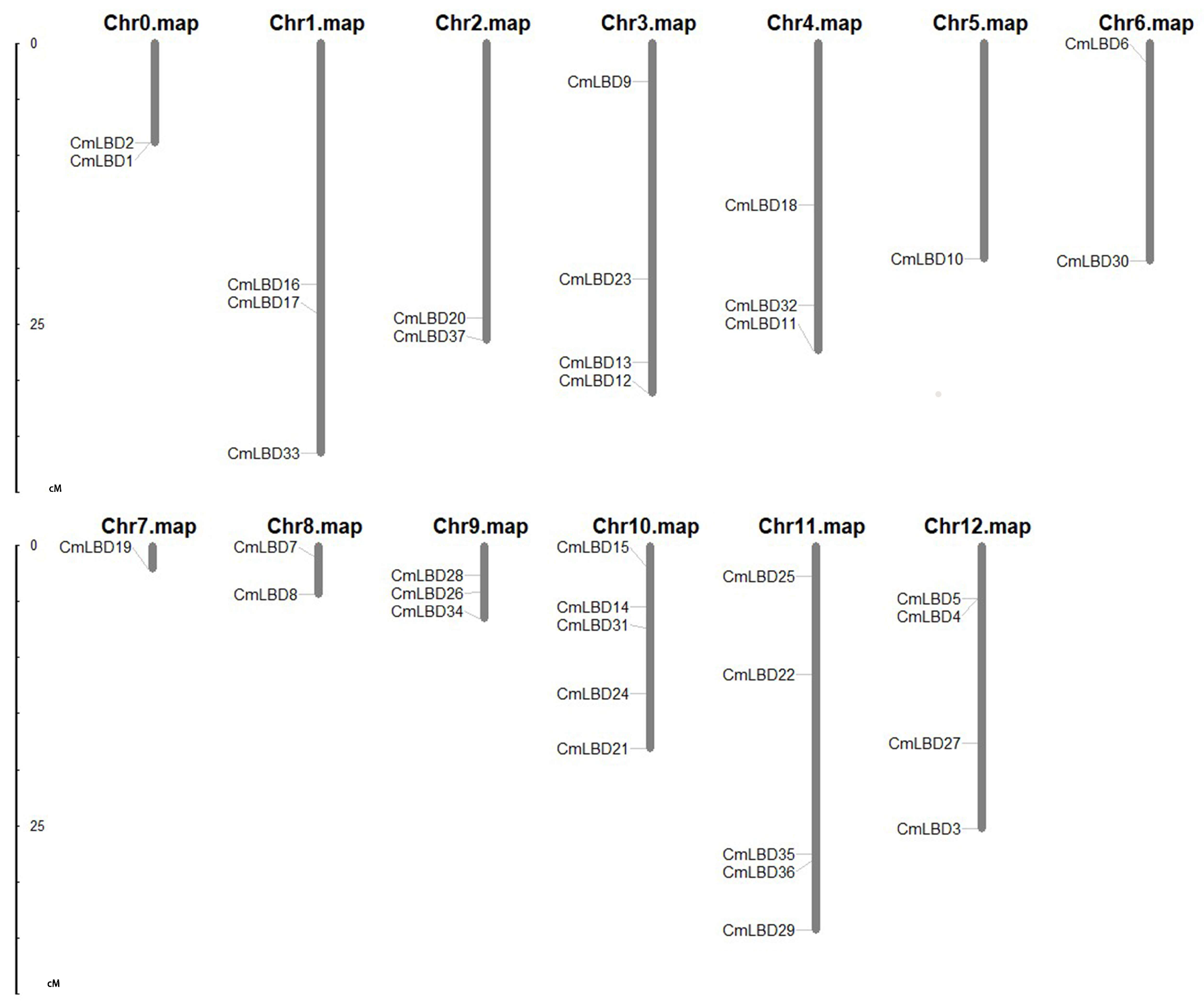
2.2. Chromosomal Position Analysis of Melon LBD Gene Family
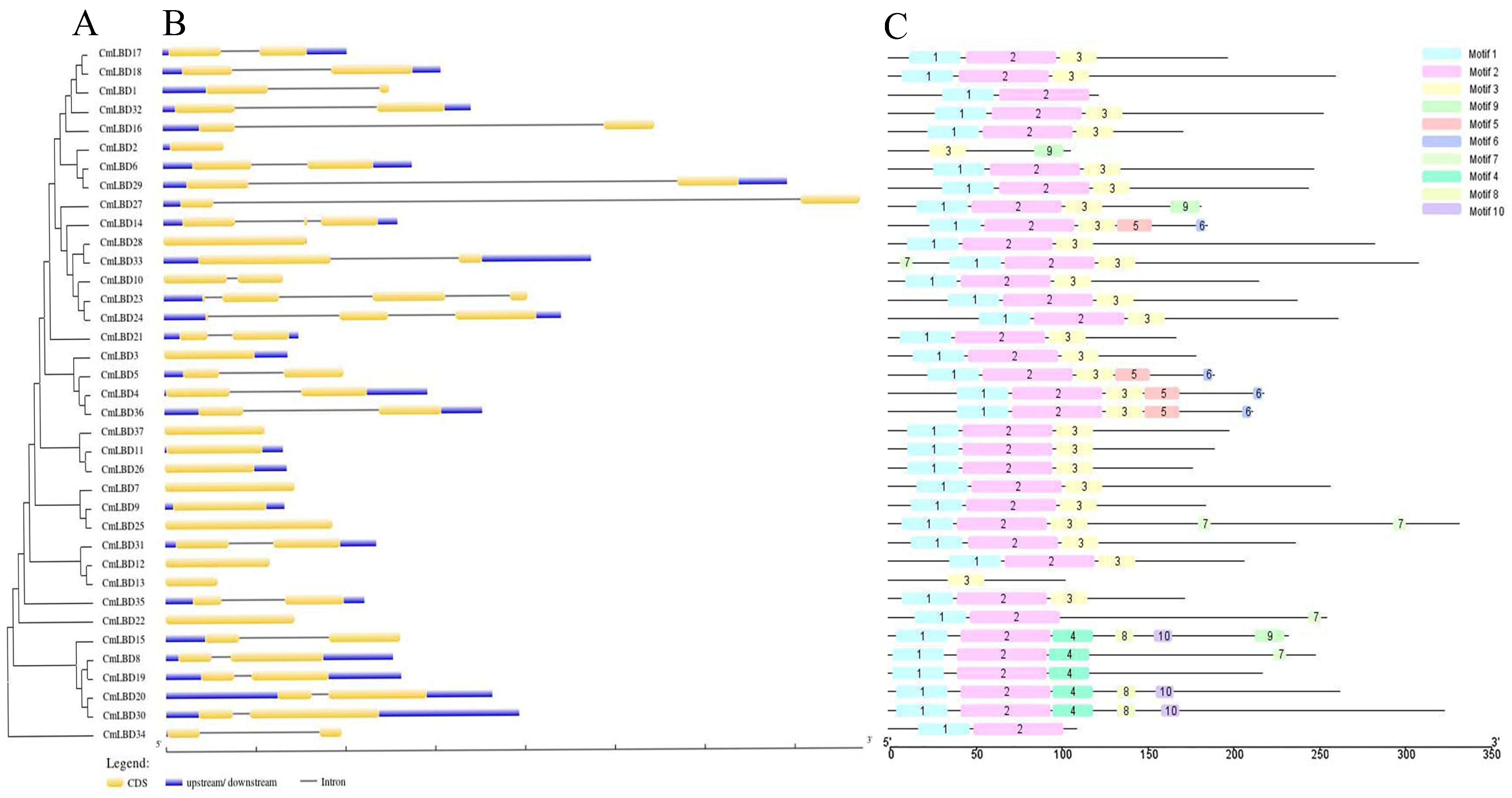
2.3. Construction of the Phylogenetic Tree for the Melon LBD Gene Family
2.4. Synteny Analysis of Melon LBD Gene Family
2.5. Gene Structure Analysis of the Melon LBD Family
2.6. Expression Pattern Analysis of Melon LBD Gene Family
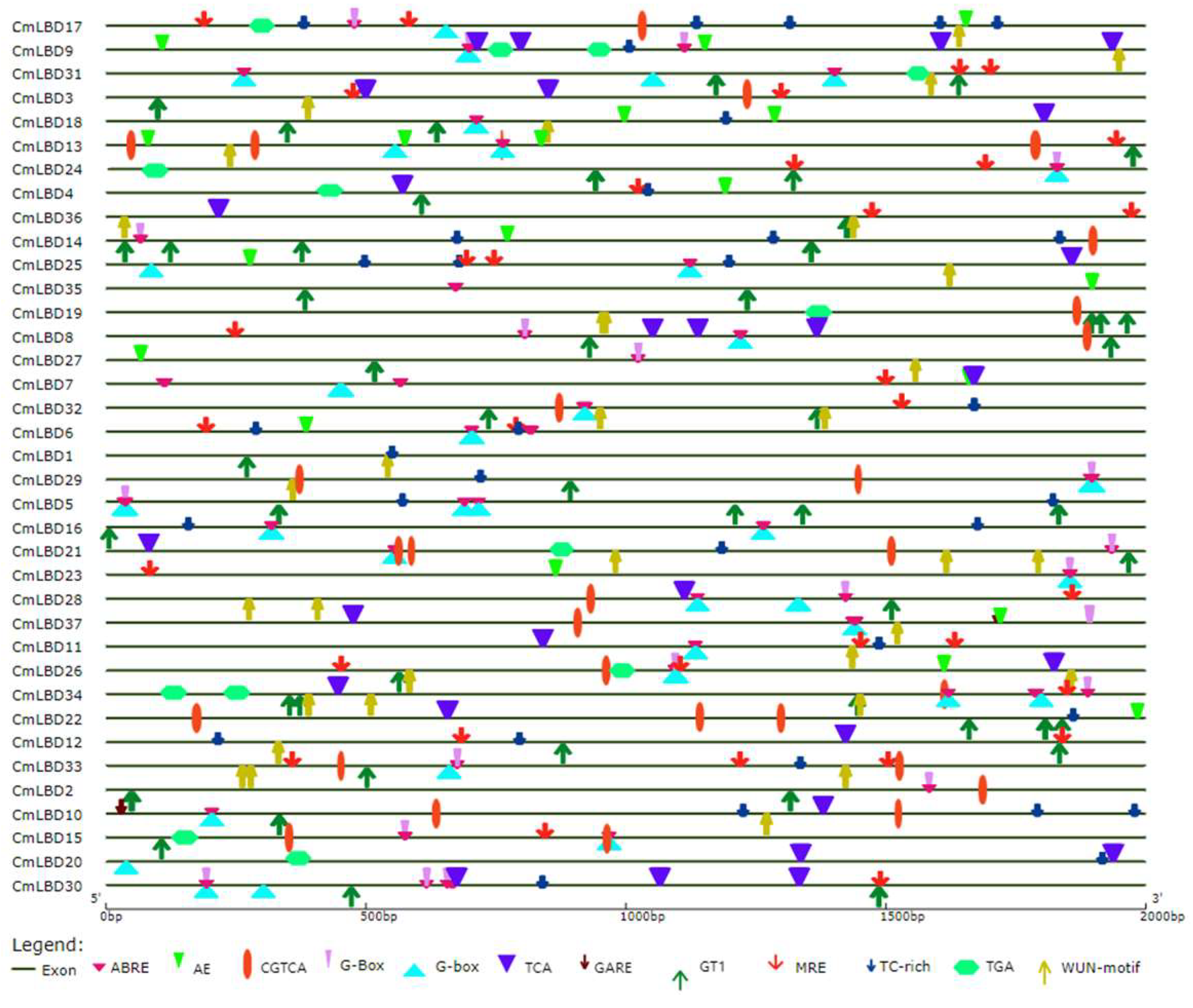
2.7. Promoter Analysis of the Melon LBD Gene Family
2.8. RNA Extraction, cDNA Synthesis, and qRT-PCR of the Melon LBD Gene Family
2.9. Protein-Protein Interaction Network of CmLBDs
3. Results
3.1. Identification and Analysis of Melon LBD Gene Family
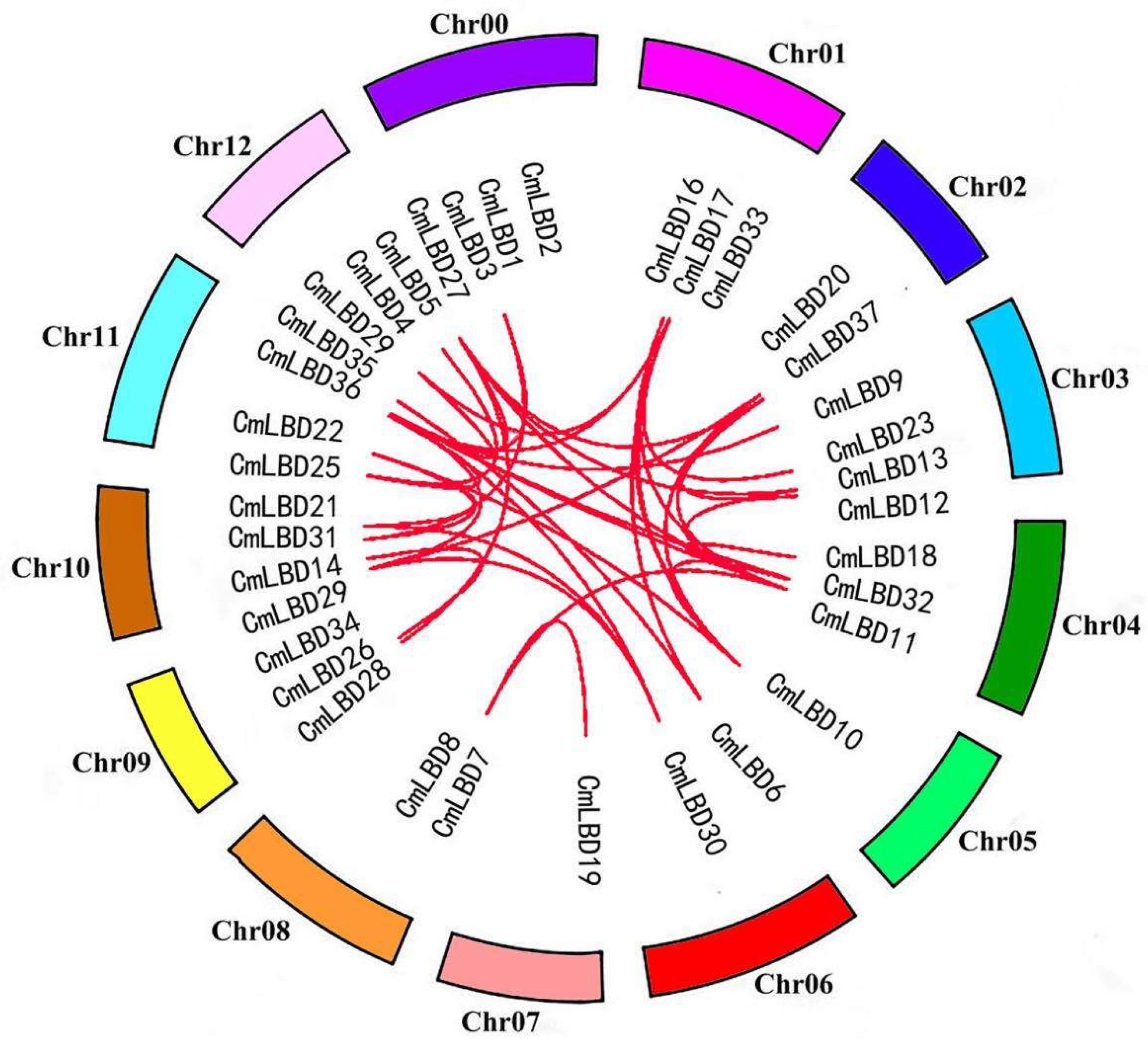
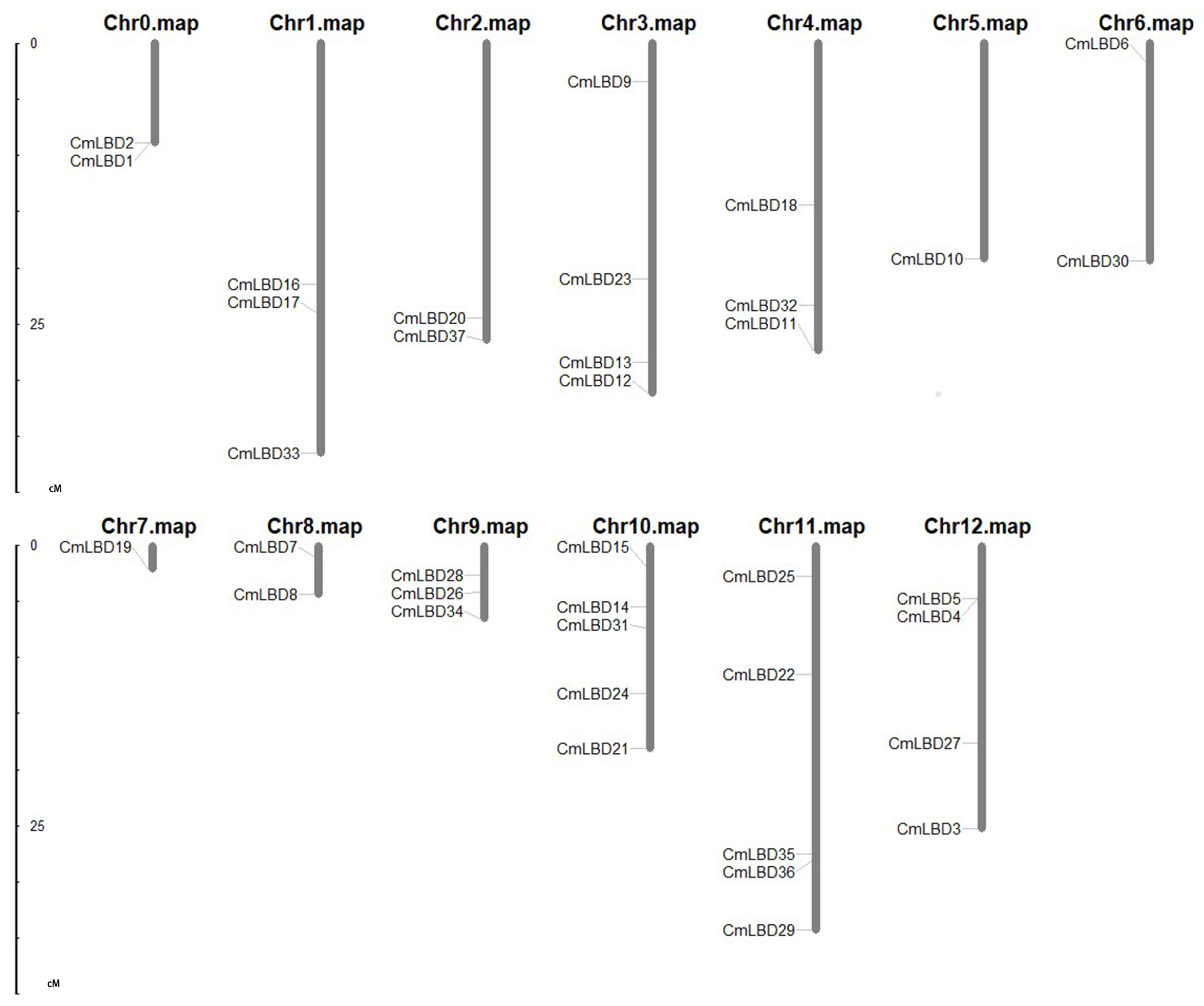
3.2. Chromosomal Distribution and Synteny Analysis of Melon LBD Gene Family
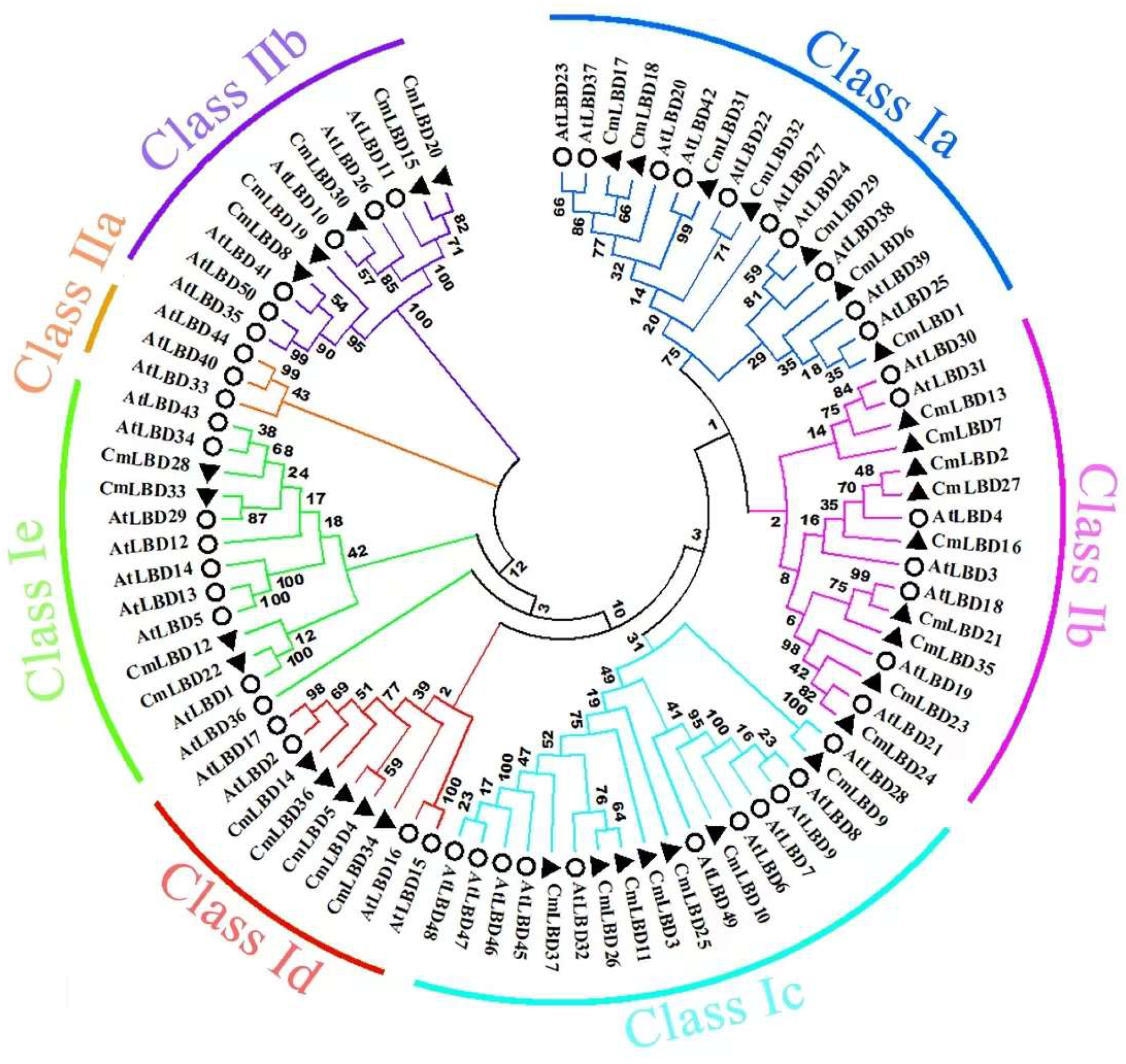
3.3. Construction of the Phylogenetic Tree of Melon and Arabidopsis LBD Gene Family Members
3.4. Conservation Domain Analysis of Melon LBD Family Genes
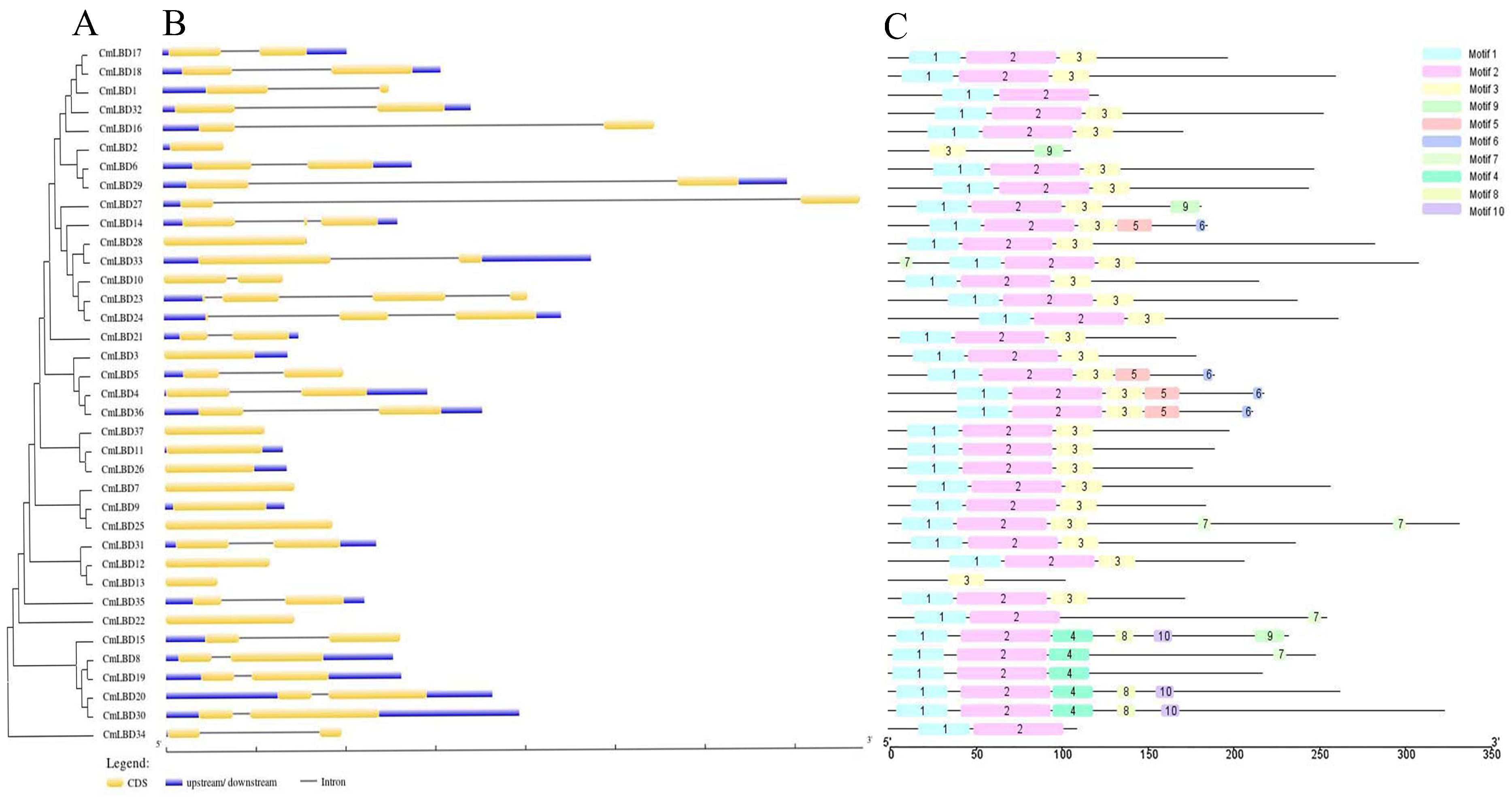
3.5. Gene Structure and Conserved Motif Analysis of Melon LBD Family Genes
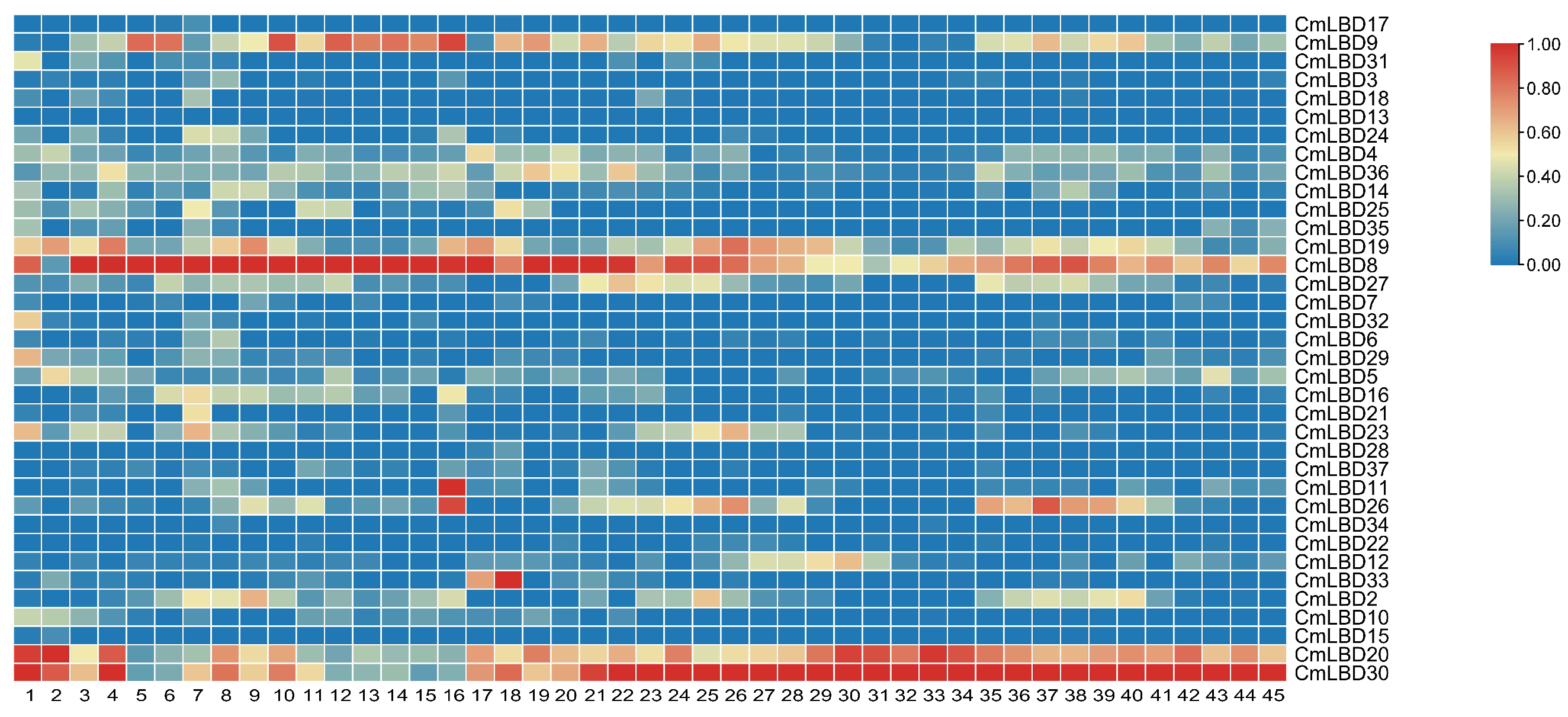
3.6. Expression Pattern Analysis of Melon LBD Family Genes
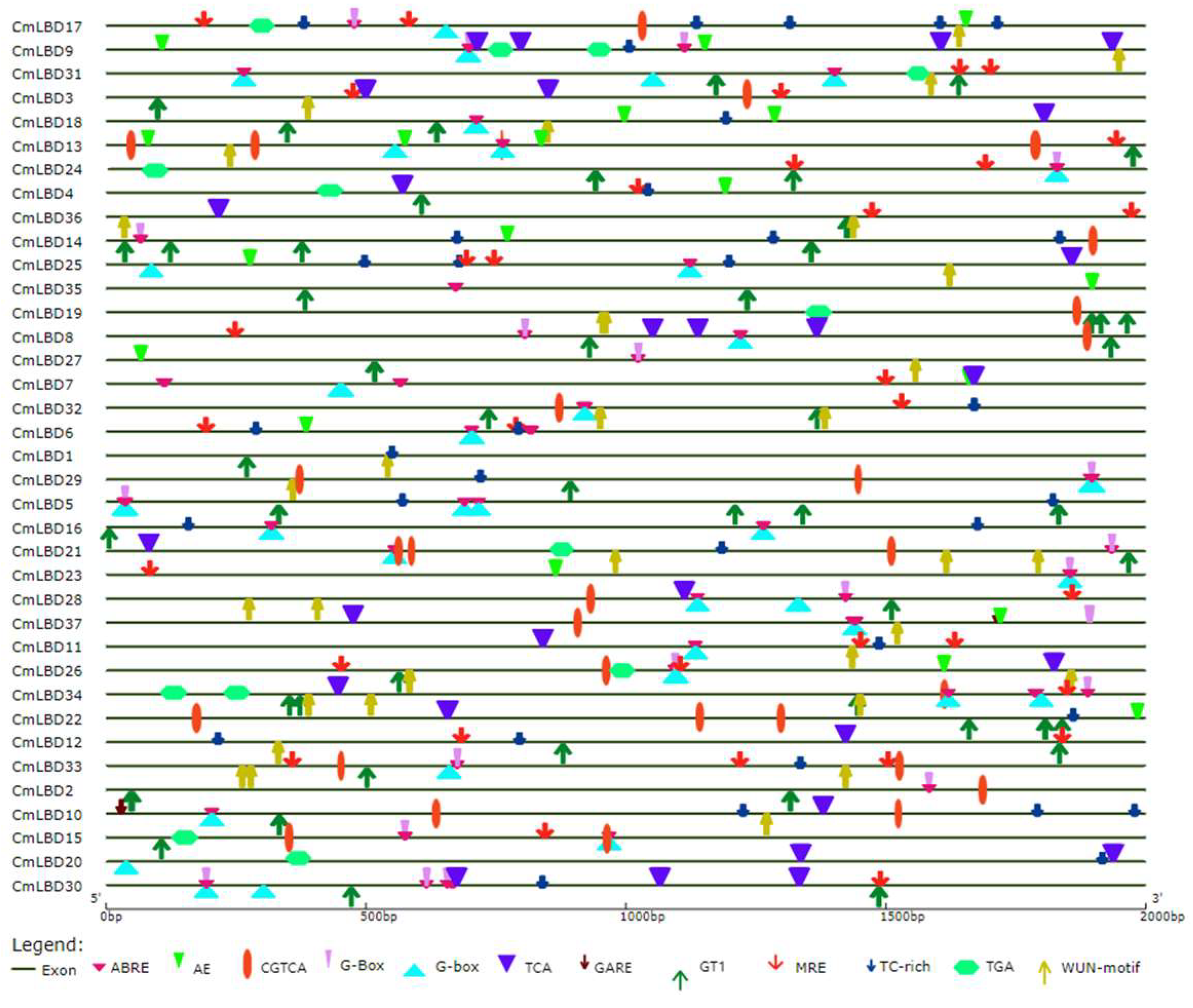
3.7. Cis-Acting Element Analysis of Melon LBD Family Genes
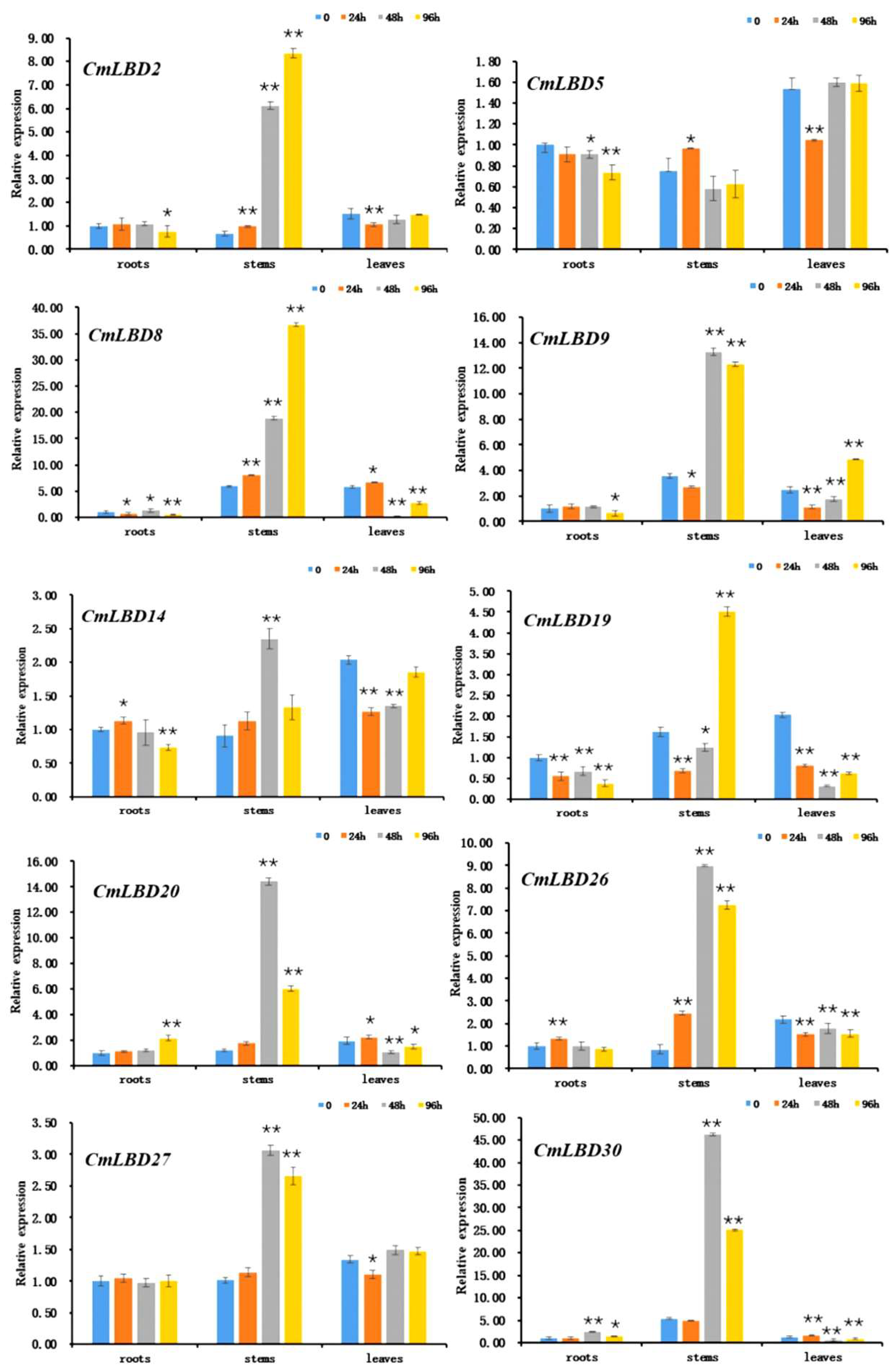
3.8. Expression of Melon LBD Family Genes in Response to Wilt Disease Pathogen
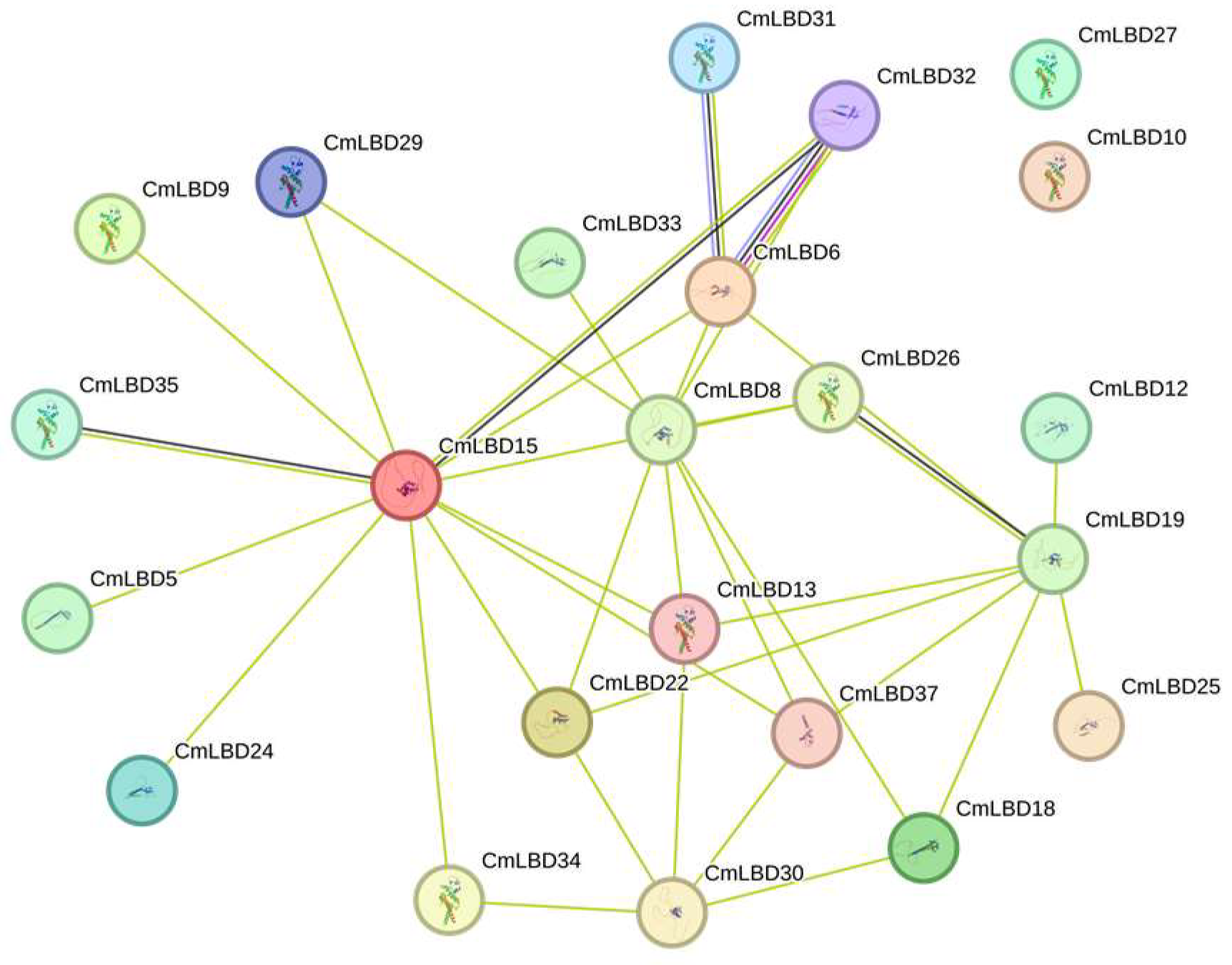
3.9. Protein Interactions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ma, W.; Wu, F.; Sheng, P.; Wang, X.; Zhang, Z.; Zhou, K.; Zhang, H.; Hu, J.; Lin, Q.; Cheng, Z.; et al. Rice LBD 12-1 reduces meristem size by inhibiting ago10-hd-zip III pathway. Plant Physiol. 2017, 173, 801–811. [Google Scholar] [CrossRef] [PubMed]
- Majer, C.; Hochholdinger, F. Defining the boundaries: Structure and function of LOB domain proteins. Trends Plant Sci. 2010, 16, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Shuai, B.; Reynaga-Pena, C.G.; Springer, P.S. The lateral organ boundaries gene defines a novel, plant-specific gene family. Plant Physiol. 2002, 129, 747–761. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Kim, M.J.; Park, M.Y.; Han, K.H.; Kim, J. The conserved proline residue in the LOB domain of LBD18 is critical for DNA-binding and biological function. Mol. Plant 2013, 6, 1722–1725. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, S.; Yu, X.; Yu, J.; He, X.; Zhang, S.; Shou, H.; Wu, P. ARL1, a LOB-domain protein required for adventitious root formation in rice. Plant J. 2005, 43, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Xie, Q.; Li, C.; Dong, Y.; Zhu, S.; Chen, J. Comprehensive characterization and gene expression patterns of LBD gene family in Gossypium Planta. Planta 2020, 251, 81. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, S.; Su, L.; Liu, X.; Hao, Y. A Genome-Wide Analysis of the LBD Gene Family in Malus domestica with a Functional Characterization of MdLBD11. PLoS ONE 2013, 8, e57044. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, J.; Zhao, M.; Yuan, H. Identification and expression analysis of Lateral Organ Bound Aries Domain (LBD) transcription factor genes in Fragaria vesca. Plant Sci. 2017, 23, 4700. [Google Scholar] [CrossRef]
- Rubin, G.; Tohge, T.; Matsuda, F.; Saito, K.; Scheible, W.R. Members of the LBD family of transcription factors repress anthocyanin synthesis and affect additional nitrogen responses in Arabidopsis. Plant Cell 2009, 21, 3567–3584. [Google Scholar] [CrossRef]
- Albinsky, D.; Kusano, M.; Higuchi, M.; Hayashi, N.; Kobayashi, M.; Fukushima, A.; Mori, M.; Ichikawa, T.; Matsui, K.; Kuroda, H.; et al. Metabolomic screening applied to rice FOX Arabidopsis lines leads to the identification of a gene-changing nitrogen metabolism. Mol. Plant 2010, 3, 125–142. [Google Scholar] [CrossRef]
- Lavenus, J.; Goh, T.; Roberts, I.; Guyomarc’hv, S.; Lucas, M.; Smet, I.D.; Fukaki, H.; Beeckman, T.; Bennett, M.; Laplaze, L. Lateral root development in Arabidopsis: Fifty shades of auxin. Trends Plant Sci. 2013, 18, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Gil, J.F.; Liebe, S.; Thiel, H.; Lennefors, B.; Kraft, T.; Gilmer, D.; Maiss, E.; Varrelmann, M.; Savenkov, E.I. Massive up-regulation of LBD transcription factors and EXPANSINs highlights the regulatory programs of rhizomania disease. Mol. Plant Pathol. 2018, 19, 2333–2348. [Google Scholar] [CrossRef]
- Cao, H.; Liu, C.Y.; Liu, C.X.; Zhao, Y.L.; Xu, R.R. Genomewide analysis of the lateral organ boundaries domain gene family in Vitis vinifera. J. Genet. Genom. 2016, 95, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.M. Response Mechanism of Black Seed Pumpkin to Fusarium Wilt Infection and Screening of NBS Resistance Genes; Southwest University: Chongqing, China, 2019. (In Chinese) [Google Scholar]
- Huang, S.X.; Gao, Y.F.; Liu, J.K.; Peng, X.L.; Niu, X.L.; Fei, Z.J.; Cao, S.Q.; Liu, Y.S. Genome-wide analysis of Wrky transcription factors in Solanum lycopersicum. Mol. Genet. Genom. 2012, 287, 495–511. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yu, X.; Wu, P. Comparison and evolution analysis of two rice subspecies LATERAL ORGAN BOUNDARIES domain gene family and their evolutionary characterization from Arabidopsis. Mol. Phylogenet. Evol. 2006, 9, 248–262. [Google Scholar] [CrossRef] [PubMed]
- Taramino, G.; Sauer, M.; Stauffer, J.L.; Multani, D.; Niu, X.; Sakai, H.; Hochholdinger, F. The maize (Zea mays L.) RTCS gene encodes a LOB domain protein that is a key regulator of embryonic seminal and post-embryonic shoot-borne root initiation. Plant J. 2007, 50, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Evans, M. The indeterminate gametophyte1 gene of maize encodes a LOB domain protein required for embryo sac and leaf development. Plant Cell 2007, 19, 46–62. [Google Scholar] [CrossRef] [PubMed]
- Bortiri, E.; Chuck, G.; Vollbrecht, E.; Rocheford, T.; Martienssen, R.; Hake, S. ramosa2 encodes a LATERAL ORGAN BOUNDARY domain protein that determines the fate of stem cells in branch meristems of maize. Plant Cell 2006, 18, 574–585. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.L.; Yan, H.Q.; Liu, Y.J.; Yi, Y. Genome-wide analysis of LATERAL ORGAN BOUNDARIES DOMAIN-in Physcomitrella patens and stress responses. Genes Genom. 2020, 42, 651–662. [Google Scholar] [CrossRef]
- Liang, J.; Hou, Z.; Liao, J.; Qin, Y.; Wang, L.; Wang, X.; Su, W.; Cai, Z.; Fang, Y.; Aslam, M.; et al. Genome-Wide Identification and Expression Analysis of LBD Transcription Factor Genes in Passion Fruit (Passiflora edulis). Int. J. Mol. Sci. 2022, 23, 4700. [Google Scholar] [CrossRef]
- Sylvia, C.; Sun, J.; Zhang, Y.; Ntini, C.; Ogutu, C.; Zhao, Y.; Han, Y. Genome-wide analysis of ATP binding cassette (ABC) transporters in peach (Prunus persica) and identification of a Gene PpABCC1 involved in anthocyanin accumulation. Int. J. Mol. Sci. 2023, 24, 1931. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, W.; Cheng, Y.; Feng, L. Genome-Wide Identification of LATERAL ORGAN BOUNDARIES DOMAIN (LBD) Transcription Factors and Screening of Salt Stress Candidates of Rosa rugosa Thunb. Biology 2021, 10, 992. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, H.; Zheng, Y.F.; Huang, R.D.; Liu, C.S. Identification of LBD gene family in Amaranthus tricolor L. based on transcriptome and expression analysis under blue light. J. China Agric. Univ. 2021, 26, 66–80. [Google Scholar] [CrossRef]
- Semiarti, E.; Ueno, Y.; Iwakawa, H.; Tsukaya, H.; Machida, C.; Machida, Y. The Asymmetric Leaves2 (AS2) gene of Arabidopsis thaliana regulates Lamina formation and is required for patterning of leaf venation. Prog. Biotechnol. 2001, 18, 63–68. [Google Scholar] [CrossRef]
- Nakazawa, M.; Ichikawa, T.; Ishikawa, A.; Kobayashi, H.; Tsuhara, Y.; Kawashima, M.; Suzuki, K.; Muto, S.; Matsui, M. Activation tagging, a novel tool to dissect the functions of a gene family. Plant J. 2003, 34, 741–750. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.C.; Guo, W.L.; Wang, Z.J. Bioinformatics analysis of the whole genome LBD gene family in Carya illinoensis. J. Zhejiang A F Univ. 2021, 38, 464–475. [Google Scholar] [CrossRef]
- Zheng, Z.F.; Zhang, Y.L.; Hu, C.; Dai, X.Z.; Liu, F.; Yuan, Z.H. Genome-wide Identification and Expressing Analysis of LBD Transcription Factors in Pepper. Acta Hortic. Sin. 2016, 43, 683–694. [Google Scholar] [CrossRef]
- Shi, Y.; Shen, S.Y.; Zhang, Q.Y.; Shun, Z.M.; Wu, R.L.; Guo, Y.Q. Research progress of LBD gene family. Chin. J. Cell Biol. 2019, 41, 738–745. [Google Scholar] [CrossRef]
- Xu, L. De novo root regeneration from leaf explants: Wounding, auxin, and cell fate transition. Curr. Opin. Plant Biol. 2018, 41, 39–45. [Google Scholar] [CrossRef]
- Thatcher, L.F.; Powell, J.J.; Aitken, E.A.B.; Kazan, K.; Manners, J.M. The lateral organ boundaries domain transcription factor LBD20 functions in Fusarium wilt susceptibility and Jasmonate signaling in Arabidopsis. Plant Physiol. 2012, 160, 407–418. [Google Scholar] [CrossRef]
- Lang, D.; Yu, X.; Jia, X.; Li, Z.X.; Zhang, X.H. Methyl jasmonate improves metabolism and growth of NaCl-stressed Glycyrrhiza uralensis seedlings. Sci. Hortic. 2020, 266, 109287. [Google Scholar] [CrossRef]
- Gómez, S.; Ferrieri, R.A.; Schueller, M.; Orians, C.M. Methyl jasmonate elicits rapid changes in carbon and nitrogen dynamics in tomato. New Phytol. 2010, 188, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Abdoulaye, A.H.; Song, J.; Yao, X.; Zhang, Y.; Gao, Y.; Ye, Y.; Luo, K.; Xia, W.; Chen, Y. Genome-wide analyses of LATERAL ORGAN BOUNDARIES in cassava reveal the role of LBD47 in defence against bacterial blight. PLoS ONE 2023, 18, e0282100. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.X.; Zhang, M.F. Research Progress on Watermelon Fusarium Wilt. China Watermelon Melon 2004, 1, 17–19. (In Chinese) [Google Scholar]
- Lü, G.; Guo, S.; Zhang, H.; Geng, L.; Song, F.; Fei, Z.; Xu, Y. Transcriptional profiling of watermelon during its incompatible interaction with Fusarium oxysporum f. sp. niveum. Eur. J. Plant Pathol. 2011, 131, 585–601. [Google Scholar] [CrossRef]
- Zhou, X.; Wu, F. Differentially expressed transcripts from cucumber (Cucumis sativus L.) root upon inoculation with Fusarium oxysporum f. sp. cucumerinum Owen. Physiol. Mol. Plant Pathol. 2009, 74, 142–150. [Google Scholar] [CrossRef]
- Sun, J.; Wei, D.; Qin, L.; Wei, Y.; Guo, W.; Yu, W.; Zhang, Z.; Lu, J. Application of High-Throughput Sequencing Technology in the Research of Banana Resistance to Wilt Disease. J. South. Agric. 2014, 45, 1921–1925. (In Chinese) [Google Scholar]
- Thatcher, L.F.; Kazan, K.; Manners, J.M. Lateral organ boundaries domain transcription factors: New roles in plant defense. Plant Signal. Behavior. 2012, 7, 1702–1704. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Symbol | Protein Length (Amino acids) | Molecular Weight (Da) | Isoelectric Point | Instability Index | Fat Solubility Index | Mean Hydrophilic Value | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|
| CmLBD1 | MELO3C000068P1 | 115 | 11,743.48 | 9.06 | 55.11 | 76.43 | 0.163 | Nucleus |
| CmLBD2 | MELO3C000076P1 | 99 | 10,950.40 | 5.67 | 61.24 | 69.90 | −0.268 | Nucleus |
| CmLBD3 | MELO3C002290P1 | 167 | 18,814.35 | 8.52 | 47.79 | 73.05 | −0.359 | Nucleus |
| CmLBD4 | MELO3C005009P1 | 204 | 22,340.62 | 6.41 | 72.35 | 91.86 | −0.131 | Nucleus |
| CmLBD5 | MELO3C005013P1 | 177 | 20,037.16 | 7.60 | 62.04 | 82.20 | −0.265 | Nucleus |
| CmLBD6 | MELO3C006183P1 | 231 | 24,953.60 | 8.93 | 80.51 | 80.69 | −0.216 | Nucleus |
| CmLBD7 | MELO3C007098P1 | 240 | 27,293.82 | 7.62 | 74.98 | 64.62 | −0.634 | Nucleus |
| CmLBD8 | MELO3C007620P1 | 232 | 25,560.77 | 7.58 | 66.53 | 69.40 | −0.390 | Nucleus |
| CmLBD9 | MELO3C008309P1 | 172 | 19,111.61 | 8.10 | 62.87 | 68.72 | −0.322 | Nucleus |
| CmLBD10 | MELO3C008779P1 | 201 | 22,171.17 | 8.96 | 54.69 | 70.90 | −0.392 | Nucleus |
| CmLBD11 | MELO3C009989P1 | 177 | 19,508.68 | 7.58 | 68.27 | 54.63 | −0.640 | Nucleus |
| CmLBD12 | MELO3C010752P1 | 193 | 21,546.77 | 7.99 | 51.03 | 88.13 | −0.186 | Nucleus |
| CmLBD13 | MELO3C011135P1 | 96 | 10,508.77 | 4.75 | 31.24 | 87.60 | −0.089 | Nucleus |
| CmLBD14 | MELO3C011730P1 | 173 | 19,126.14 | 6.56 | 79.98 | 82.37 | −0.148 | Nucleus |
| CmLBD15 | MELO3C012250P1 | 217 | 23,478.48 | 8.45 | 36.69 | 72.40 | −0.328 | Nucleus |
| CmLBD16 | MELO3C012624P1 | 160 | 17,234.61 | 8.14 | 72.61 | 70.19 | −0.159 | Nucleus |
| CmLBD17 | MELO3C012741P1 | 184 | 20,243.07 | 6.28 | 68.12 | 80.22 | 0.014 | Nucleus |
| CmLBD18 | MELO3C012908P1 | 243 | 27,013.91 | 6.04 | 46.60 | 63.46 | −0.500 | Nucleus |
| CmLBD19 | MELO3C016808P1 | 203 | 22,236.13 | 6.50 | 56.75 | 80.69 | −0.202 | Nucleus |
| CmLBD20 | MELO3C017305P1 | 245 | 26,763.80 | 8.72 | 54.14 | 77.63 | −0.240 | Nucleus |
| CmLBD21 | MELO3C018380P1 | 156 | 17,169.95 | 7.52 | 61.42 | 86.86 | 0.023 | Nucleus |
| CmLBD22 | MELO3C019314P1 | 238 | 26,823.12 | 8.88 | 46.86 | 67.94 | −0.772 | Nucleus |
| CmLBD23 | MELO3C019995P1 | 222 | 23,851.07 | 9.10 | 79.33 | 75.63 | −0.223 | Nucleus |
| CmLBD24 | MELO3C020121P1 | 244 | 26,813.14 | 8.61 | 76.41 | 70.00 | −0.491 | Nucleus |
| CmLBD25 | MELO3C020944P1 | 310 | 34,948.92 | 7.12 | 67.35 | 64.87 | −0.792 | Nucleus |
| CmLBD26 | MELO3C021578P1 | 165 | 18,202.35 | 7.66 | 50.92 | 66.30 | −0.524 | Nucleus |
| CmLBD27 | MELO3C021789P1 | 170 | 18,490.96 | 8.32 | 62.51 | 73.47 | −0.281 | Nucleus |
| CmLBD28 | MELO3C021964P1 | 265 | 29,996.60 | 5.00 | 55.56 | 75.81 | −0.570 | Nucleus |
| CmLBD29 | MELO3C022505P1 | 228 | 24,793.47 | 6.97 | 47.74 | 80.44 | −0.145 | Nucleus |
| CmLBD30 | MELO3C023802P1 | 302 | 32,712.42 | 8.86 | 51.46 | 82.35 | −0.272 | Nucleus |
| CmLBD31 | MELO3C023880P1 | 221 | 24,271.06 | 4.45 | 48.49 | 78.10 | −0.051 | Nucleus |
| CmLBD32 | MELO3C024038P1 | 236 | 25,568.78 | 7.06 | 45.07 | 75.34 | −0.250 | Nucleus |
| CmLBD33 | MELO3C024387P1 | 288 | 32,397.20 | 5.42 | 51.16 | 76.60 | −0.544 | Nucleus |
| CmLBD34 | MELO3C025504P1 | 102 | 11,653.77 | 9.43 | 52.87 | 76.47 | −0.141 | Nucleus |
| CmLBD35 | MELO3C025701P1 | 161 | 18,044.50 | 4.94 | 56.38 | 73.35 | −0.207 | Nucleus |
| CmLBD36 | MELO3C025742P1 | 198 | 21,505.30 | 5.59 | 75.26 | 81.36 | −0.182 | Nucleus |
| CmLBD37 | MELO3C026269P1 | 185 | 20,363.90 | 7.63 | 53.81 | 71.30 | −0.405 | Nucleus |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, L.; Chao, Y.; Wang, Y.; Xu, Y.; Li, S. Genome-Wide Analysis of the LBD Gene Family in Melon and Expression Analysis in Response to Wilt Disease Infection. Genes 2024, 15, 442. https://doi.org/10.3390/genes15040442
Zheng L, Chao Y, Wang Y, Xu Y, Li S. Genome-Wide Analysis of the LBD Gene Family in Melon and Expression Analysis in Response to Wilt Disease Infection. Genes. 2024; 15(4):442. https://doi.org/10.3390/genes15040442
Chicago/Turabian StyleZheng, Ling, Yanrong Chao, Yian Wang, Yizhuo Xu, and Shipeng Li. 2024. "Genome-Wide Analysis of the LBD Gene Family in Melon and Expression Analysis in Response to Wilt Disease Infection" Genes 15, no. 4: 442. https://doi.org/10.3390/genes15040442
APA StyleZheng, L., Chao, Y., Wang, Y., Xu, Y., & Li, S. (2024). Genome-Wide Analysis of the LBD Gene Family in Melon and Expression Analysis in Response to Wilt Disease Infection. Genes, 15(4), 442. https://doi.org/10.3390/genes15040442