

D299T Mutation in CYP76F14 Led to a Decrease in Wine Bouquet Precursor Production in Wine Grape


Abstract
1. Introduction
2. Results
2.1. Difference in Wine Bouquet Precursors Among Different Wine Grape Varieties
2.2. D299T Variation Was Observed in VvCYP76F14s of Neutral Varieties or Offspring Lines
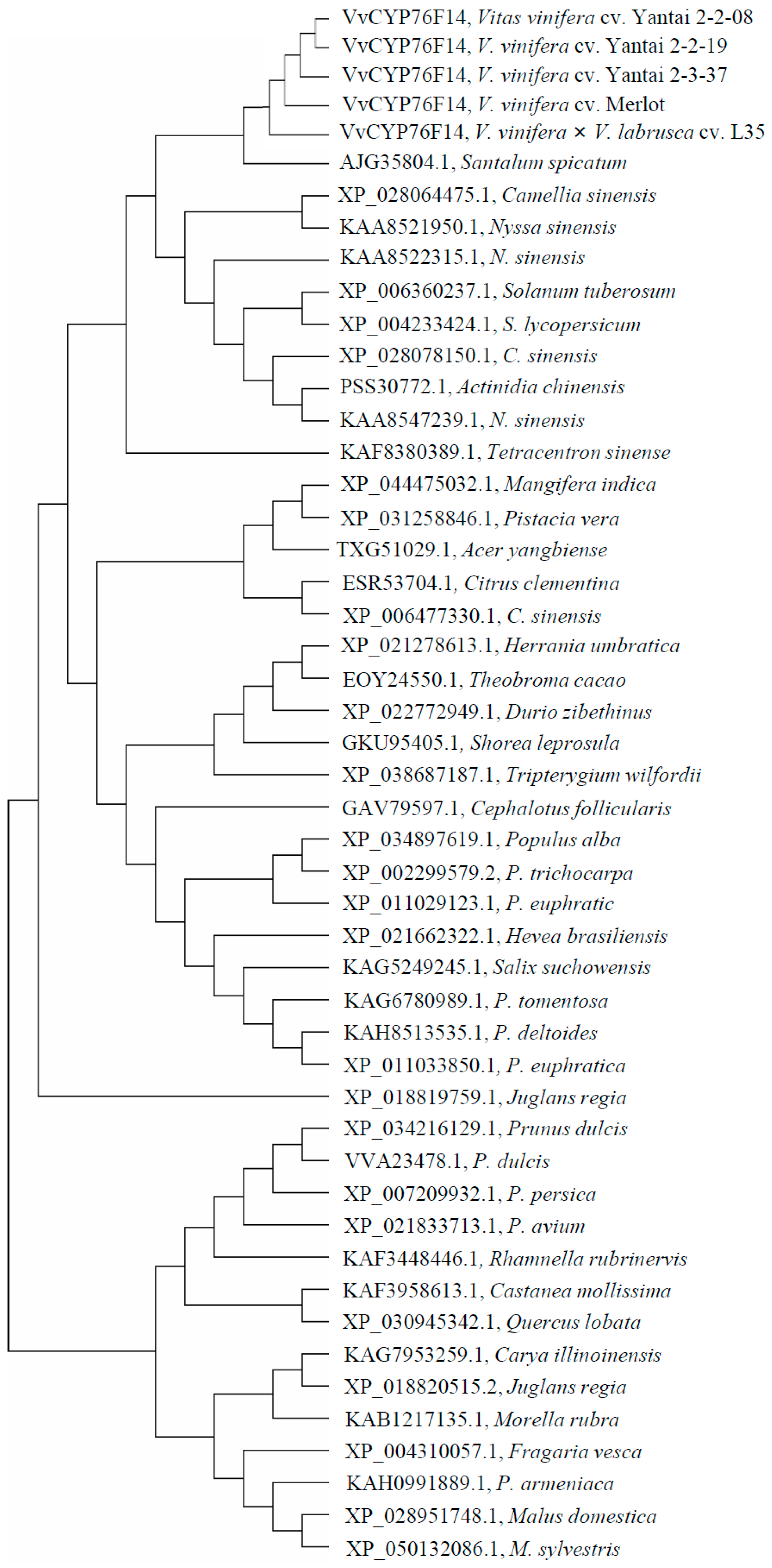
2.3. Phylogenetic Tree Analysis of Plant CYP76 Family Homologs
2.4. VvCYP76F14 Is Localized in the Endoplasmic Reticulum
2.5. Neutral-Type VvCYP76F14s Exhibited Lower Enzymatic Activity In Vitro
2.6. Transient Expression of the ‘L35’ or ‘Merlot’ VvCYP76F14 Enhanced the Wine Bouquet Precursor Contents in the D299T Substitution Mutant Lines
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Wine Grape
4.3. Analysis of Linalool and Linalool-Derived Compounds in Wine Grape Berries
4.4. Isolation and Sequence Analysis of VvCYP76F14s from Different Varieties or Offspring Lines
4.5. Phylogenetic Tree Analysis
4.6. Real-Time Quantitative PCR
4.7. Subcellular Localization of VvCYP76F14
4.8. Heterologous Expression of VvCYP76F14 in E. coli
4.9. In Vitro Enzymatic Activity Determination
4.10. Over-Expression of VvCYP76F14 in Berries of D299T Mutant Lines
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alegre, Y.; Saenz-Navajas, M.P.; Hernandez-Orte, P.; Ferreira, V. Sensory, olfactometric and chemical characterization of the aroma potential of Garnacha and Tempranillo winemaking grapes. Food Chem. 2020, 331, 127207. [Google Scholar] [CrossRef] [PubMed]
- Robinson, A.L.; Boss, P.K.; Solomon, P.S.; Trengove, R.D.; Heymann, H.; Ebeler, S.E. Origins of grape and wine aroma. Part 2. Chemical and sensory analysis. Am. J. Enol. Vitic. 2014, 65, 25–42. [Google Scholar] [CrossRef]
- Thomas-Danguin, T.; Ishii-Foret, A.; Atanasova, B.; Etievant, P. Wine bouquet: The perceptual integration of chemical complexity. In Wine Active Compounds; Conference Object; Oenoplurimédia: Beaune, France, 2011. [Google Scholar]
- Zhai, H.Y.; Li, S.Y.; Zhao, X.; Lan, Y.B.; Zhang, X.K.; Shi, Y.; Duan, C.Q. The compositional characteristics, influencing factors, effects on wine quality and relevant analytical methods of wine polysaccharides: A review. Food Chem. 2023, 403, 134467. [Google Scholar] [CrossRef] [PubMed]
- Noguerol-Pato, R.; Gonzalez-Barreiro, C.; Cancho-Grande, B.; Santiago, J.L.; Martinez, M.C.; Simal-Gandara, J. Aroma potential of Brancellao grapes from different cluster positions. Food Chem. 2012, 132, 112–124. [Google Scholar] [CrossRef]
- Lukic, I.; Lotti, C.; Vrhovsek, U. Evolution of free and bound volatile aroma compounds and phenols during fermentation of Muscat blanc grape juice with and without skins. Food Chem. 2017, 232, 25–35. [Google Scholar] [CrossRef]
- Wang, X.J.; Tao, Y.S.; Wu, Y.; An, R.Y.; Yue, Z.Y. Aroma compounds and characteristics of noble-rot wines of Chardonnay grapes artificially botrytized in the vineyard. Food Chem. 2017, 226, 41–50. [Google Scholar] [CrossRef]
- Peng, B.; Ran, J.G.; Li, Y.Y.; Tang, M.L.; Xiao, H.L.; Shi, S.P.; Ning, Y.Z.; Dark, A.; Guan, X.Q.; Song, Z.Z. Site-directed mutagenesis of VvCYP76F14 (cytochrome P450) unveils its potential for selection in wine grape varieties linked to the development of wine bouquet. J. Agric. Food Chem. 2024, 72, 3683–3694. [Google Scholar] [CrossRef]
- Parker, M.; Capone, D.L.; Francis, I.L.; Herderich, M.J. Aroma Precursors in Grapes and Wine: Flavor Release during Wine Production and Consumption. J. Agric. Food Chem. 2017, 66, 2281–2286. [Google Scholar] [CrossRef]
- Ghaste, M.; Narduzzi, L.; Carlin, S.; Vrhovsek, U.; Shulaev, V.; Mattivi, F. Chemical composition of volatile aroma metabolites and their glycosylated precursors that can uniquely differentiate individual grape cultivars. Food Chem. 2015, 188, 309–319. [Google Scholar] [CrossRef]
- Lin, J.; Massonnet, M.; Cantu, D. The genetic basis of grape and wine aroma. Hortic. Res. 2019, 6, 81. [Google Scholar] [CrossRef]
- Giaccio, J.; Capone, D.L.; Håkansson, A.E.; Smyth, H.E.; Elsey, G.M.; Sefton, M.A.; Taylor, D.K. The formation of wine lactone from grape-derived secondary metabolites. J. Agric. Food Chem. 2011, 59, 660–664. [Google Scholar] [CrossRef] [PubMed]
- Ilc, T.; Halter, D.; Miesch, L.; Lauvoisard, F.; Kriegshauser, L.; Ilg, A.; Baltenweck, R.; Hugueney, P.; Werck-Reichhart, D.; Duchêne, E.; et al. A grapevine cytochrome P450 generates the precursor of wine lactone, a key odorant in wine. N. Phytol. 2017, 213, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Kunert, M.; Langley, C.; Lucier, R.; Ploss, K.; López, C.Z.R.; Guerrero, A.D.S.; Rothe, E.; O’Connor, S.E. Promiscuous CYP87A enzyme activity initiates cardenolide biosynthesis in plants. Nat. Plants 2023, 9, 1607–1617. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Lewis, K.M.; Abril, A.; Davydov, D.R.; Vermerris, W.; Sattler, S.E.; Kang, C. Structure and function of the cytochrome P450 monooxygenase cinnamate 4-hydroxylase from Sorghum Bicolor. Plant Physiol. 2020, 183, 957–973. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Jin, G.J.; Wang, X.J.; Kong, C.L.; Liu, J.; Tao, Y.S. Chemical profiles and aroma contribution of terpene compounds in Meili (Vitis vinifera L.) grape and wine. Food Chem. 2019, 284, 155–161. [Google Scholar] [CrossRef]
- Cheng, J.; Li, J.; Zhou, P.H.; Jian, M.D.; Dong, Y.W.; Wang, T.Q.; Wu, F.; Qiu, H. Study on the difference of phenolic substances in different wine grapes from Jiaodong peninsula Shandong province. Food Eng. 2020, 1, 71–75. (In Chinese) [Google Scholar]
- Hofer, R.; Dong, L.; Andre, F.; Ginglinger, J.F.; Lugan, R.; Gavira, C.; Grec, S.; Lang, G.; Memelink, J.; Van der Krol, S.; et al. Geraniol hydroxylase and hydroxygeraniol oxidase activities of the CYP76 family of cytochrome P450 enzymes and potential for engineering the early steps of the (seco) iridoid pathway. Metab. Eng. 2013, 20, 221–232. [Google Scholar] [CrossRef]
- Hofer, R.; Boachon, B.; Renault, H.; Gavira, C.; Miesch, L.; Iglesias, J.; Ginglinger, J.F.; Allouche, L.; Miesch, M.; Grec, S.; et al. Dual function of the cytochrome P450 CYP76 family from Arabidopsis thaliana in the metabolism of monoterpenols and phenylurea herbicides. Plant Physiol. 2014, 166, 1149–1161. [Google Scholar] [CrossRef]
- Renault, H.; Bassard, J.E.; Hamberger, B.; Werck-Reichhart, D. Cytochrome P450-mediated metabolic engineering: Current progress and future challenges. Curr. Opin. Plant Biol. 2014, 19, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Bathe, U.; Frolov, A.; Porzel, A.; Tissier, A. CYP76 oxidation network of abietane diterpenes in lamiaceae reconstituted in yeast. J. Agric. Food Chem. 2019, 67, 3437–13450. [Google Scholar] [CrossRef]
- Walsh, C.T.; Wencewicz, T.A. Flavoenzymes: Versatile catalysts in biosynthetic pathways. Nat. Prod. Rep. 2013, 30, 175–200. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.Z.; Peng, B.; Gu, Z.X.; Tang, M.L.; Li, B.; Liang, M.X.; Wang, L.M.; Guo, X.T.; Wang, J.P.; Sha, Y.F.; et al. Site-directed mutagenesis identified the key active site residues of alcohol acyltransferase PpAAT1 responsible for aroma biosynthesis in peach fruits. Hortic. Res. 2021, 8, 32. [Google Scholar] [CrossRef] [PubMed]
- Peng, B.; Yu, M.L.; Zhang, B.B.; Xu, J.L.; Ma, R.J. Differences in PpAAT1 activity in high- and low-aroma peach varieties affect γ-decalactone production. Plant Physiol. 2020, 182, 2065–2080. [Google Scholar] [CrossRef] [PubMed]
- D’Onofrio, C.; Matarese, F.; Cuzzola, A. Effect of methyl jasmonate on the aroma of Sangiovese grapes and wines. Food Chem. 2018, 242, 352–361. [Google Scholar] [CrossRef]
- Riffle, V.L.; Arredondo, J.A.; LoMonaco, I.; Appel, C.; Catania, A.A.; Peterson, J.C.D.; Casassa, L.F. Vine age affects vine performance, grape and wine chemical and sensory composition of cv. Zinfandel from California. Am. J. Enol. Vitic. 2022, 73, 277–293. [Google Scholar] [CrossRef]
- Picard, M.; Tempere, S.; de Revel, G.; Marchand, S. A sensory study of the ageing bouquet of red Bordeaux wines: A three-step approach for exploring a complex olfactory concept. Food Qual. Prefer. 2015, 42, 110–122. [Google Scholar] [CrossRef]
- Peng, B.; Gu, Z.X.; Zhou, Y.F.; Ning, Y.Z.; Xu, H.Y.; Li, G.; Ni, Y.; Sun, P.P.; Xie, Z.Q.; Shi, S.P.; et al. Potential role of fatty acid desaturase 2 in regulating peach aroma formation. Postharvest Biol. Technol. 2023, 204, 112473. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wine Grape | Linalool (μg·g−1 FW) | (E)-8-Hydroxylinalool (μg·g−1 FW) | (E)-8-Oxolinalool (μg·g−1 FW) | (E)-8-Carboxylinalool (μg·g−1 FW) | Wine Bouquet Type |
|---|---|---|---|---|---|
| V. vinifera cv. Yantai 2-2-08 | 5.59 ± 0.61 a | 0.15 ± 0.021 c | N.D. | N.D. | Neutral |
| V. vinifera cv. Yantai 2-2-19 | 5.48 ± 0.53 a | 0.13 ± 0.018 c | N.D. | N.D. | Neutral |
| V. vinifera cv. Yantai 2-3-37 | 5.51 ± 0.54 a | 0.16 ± 0.022 c | N.D. | N.D. | Neutral |
| V. vinifera cv. Merlot | 5.74 ± 0.63 a | 1.38 ± 0.15 b | 0.89 ± 0.092 b | 0.34 ± 0.041 b | Aromatic |
| V. vinifera × V. labrusca cv. L35 | 6.01 ± 0.64 a | 3.74 ± 0.41 a | 2.23 ± 0.24 a | 0.92 ± 0.11 a | Full-Bodied |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; Xiao, H.; Shi, M.; Tang, M.; Song, Z. D299T Mutation in CYP76F14 Led to a Decrease in Wine Bouquet Precursor Production in Wine Grape. Genes 2024, 15, 1478. https://doi.org/10.3390/genes15111478
Liu W, Xiao H, Shi M, Tang M, Song Z. D299T Mutation in CYP76F14 Led to a Decrease in Wine Bouquet Precursor Production in Wine Grape. Genes. 2024; 15(11):1478. https://doi.org/10.3390/genes15111478
Chicago/Turabian StyleLiu, Wanhao, Huilin Xiao, Matthew Shi, Meiling Tang, and Zhizhong Song. 2024. "D299T Mutation in CYP76F14 Led to a Decrease in Wine Bouquet Precursor Production in Wine Grape" Genes 15, no. 11: 1478. https://doi.org/10.3390/genes15111478
APA StyleLiu, W., Xiao, H., Shi, M., Tang, M., & Song, Z. (2024). D299T Mutation in CYP76F14 Led to a Decrease in Wine Bouquet Precursor Production in Wine Grape. Genes, 15(11), 1478. https://doi.org/10.3390/genes15111478