
Modulation of Skin Cancer by the Stimulator of Interferon Genes

Abstract
:1. Introduction
2. Epidemiology and Brief Classification of Skin Cancers
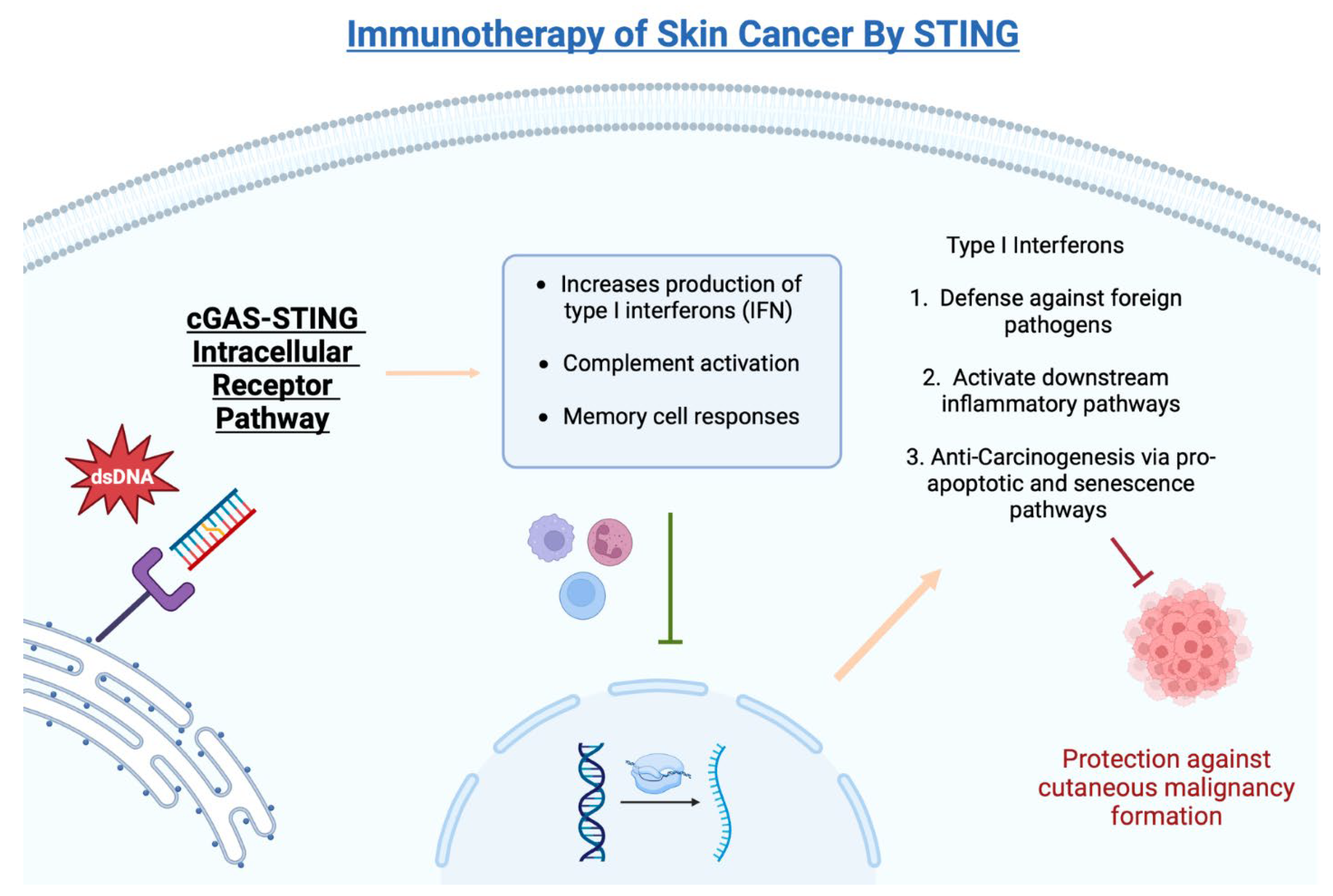
3. What Is STING (Stimulator of Interferon-Related Genes)?
4. Role of STING in DNA Damage
5. Role of STING in Immunotherapy of Skin Cancer
5.1. Melanoma
5.2. Squamous Cell Carcinoma
5.3. Basal Cell Carcinoma
5.4. STING Resistance in Tumor Cells
6. Further Potential Therapies
7. STING and Clinical Trials
8. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Balducci, L. Epidemiology of Cancer and Aging. J. Oncol. Manag. 2005, 14, 47–50. [Google Scholar] [PubMed]
- Stern, R.S. Prevalence of a history of skin cancer in 2007. Arch. Dermatol. 2010, 146, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Guy, G.P.; Thomas, C.C.; Thompson, T.; Watson, M.; Massetti, G.M.; Richardson, L.C. Melanoma Incidence and Mortality Trends and Projections—United States, 1982–2030, 21st ed.; Ser. Vital Signs; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2015; Volume 64, pp. 591–596.
- Guy, G.P.; Machlin, S.R.; Ekwueme, D.U.; Yabroff, K.R. Prevalence and costs of skin cancer treatment in the U.S., 2002–2006 and 2007–2011. Am. J. Prev. Med. 2015, 48, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Eide, M.J.; Krajenta, R.; Johnson, D.; Long, J.J.; Jacobsen, G.; Asgari, M.M.; Lim, H.W.; Johnson, C.C. Identification of patients with nonmelanoma skin cancer using health maintenance organization claims data. Am. J. Epidemiol. 2010, 171, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.H.; Mir, M.; Qian, L.; Baloch, M.; Khan, M.F.A.; Ngowi, E.E.; Wu, D.D.; Ji, X.Y. Skin cancer biology and barriers to treatment: Recent applications of polymeric micro/nanostructures. J. Adv. Res. 2022, 36, 223–247. [Google Scholar] [CrossRef]
- Sato, S.; Sawada, Y.; Nakamura, M. STING Signaling and Skin Cancers. Cancers 2021, 13, 5603. [Google Scholar] [CrossRef]
- Zhang, H.; You, Q.-D.; Xu, X.-L. Targeting stimulator of interferon genes (sting): A medicinal chemistry perspective. J. Med. Chem. 2019, 63, 3785–3816. [Google Scholar] [CrossRef]
- D’Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV radiation and the skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef]
- McDaniel, B.; Badri, T.; Steele, R.B. Basal Cell Carcinoma; StatPearls: Treasure Island, FL, USA, 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK482439/ (accessed on 2 August 2023).
- Losquadro, W.D. Anatomy of the skin and the pathogenesis of nonmelanoma skin cancer. Facial Plast. Surg. Clin. N. Am. 2017, 25, 283–289. [Google Scholar] [CrossRef]
- Sample, A.; He, Y.Y. Mechanisms and prevention of UV-induced melanoma. Photodermatol. Photoimmunol. Photomed. 2018, 34, 13–24. [Google Scholar] [CrossRef]
- Apalla, Z.; Nashan, D.; Weller, R.B.; Castellsagué, X. Skin Cancer: Epidemiology, Disease Burden, Pathophysiology, Diagnosis, and Therapeutic Approaches. Dermatol. Ther. 2017, 7 (Suppl. 1), 5–19. [Google Scholar] [CrossRef] [PubMed]
- Naik, P.P.; Desai, M.B. Basal Cell Carcinoma: A Narrative Review on Contemporary Diagnosis and Management. Oncol. Ther. 2022, 10, 317–335. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, A.S.; Ogle, C.A.; Shim, E.K. Metastatic cutaneous squamous cell carcinoma: An update. Dermatol. Surg. 2007, 33, 885–899. [Google Scholar] [CrossRef] [PubMed]
- Wehner, M.R.; Cidre Serrano, W.; Nosrati, A.; Schoen, P.M.; Chren, M.M.; Boscardin, J.; Linos, E. All-cause mortality in patients with basal and squamous cell carcinoma: A systematic review and meta-analysis. J. Am. Acad. Dermatol. 2018, 78, 663–672.e3. [Google Scholar] [CrossRef] [PubMed]
- Tokez, S.; Wakkee, M.; Kan, W.; Venables, Z.C.; Mooyaart, A.L.; Louwman, M.; Nijsten, T.; Hollestein, L.M. Cumulative incidence and disease-specific survival of metastatic cutaneous squamous cell carcinoma: A nationwide cancer registry study. J. Am. Ac. Dermatol. 2022, 86, 331–338. [Google Scholar] [CrossRef] [PubMed]
- National Cancer Institute. Melanoma of the Skin—Cancer Stat Facts. SEER. Available online: https://seer.cancer.gov/statfacts/html/melan.html (accessed on 7 August 2023).
- American Cancer Society. Melanoma Survival Rates: Melanoma Survival Statistics. Melanoma Survival Rates|Melanoma Survival Statistics|American Cancer Society. 2023. Available online: https://www.cancer.org/cancer/types/melanoma-skin-cancer/detection-diagnosis-staging/survival-rates-for-melanoma-skin-cancer-by-stage.html (accessed on 7 August 2023).
- Vashi, N.; Bakhoum, S.F. The Evolution of STING Signaling and Its Involvement in Cancer. Trends Biochem. Sci. 2021, 46, 446–460. [Google Scholar] [CrossRef]
- McNab, F.; Mayer-Barber, K.; Sher, A.; Wack, A.; O’garra, A. Type I interferons in infectious disease. Nat. Rev. Immunol. 2015, 15, 87–103. [Google Scholar] [CrossRef]
- Luo, K.; Li, N.; Ye, W.; Gao, H.; Luo, X.; Cheng, B. Activation of Stimulation of Interferon Genes (STING) Signal and Cancer Immunotherapy. Molecules 2022, 27, 4638. [Google Scholar] [CrossRef]
- Zhu, Y.; An, X.; Zhang, X.; Qiao, Y.; Zheng, T.; Li, X. STING: A master regulator in the cancer-immunity cycle. Mol. Cancer 2019, 18, 152. [Google Scholar] [CrossRef]
- Jiang, M.; Chen, P.; Wang, L.; Li, W.; Chen, B.; Liu, Y.; Wang, H.; Zhao, S.; Ye, L.; He, Y.; et al. cGAS-STING, an important pathway in cancer immunotherapy. J. Hematol. Oncol. 2020, 13, 81. [Google Scholar] [CrossRef]
- Hoong, B.Y.D.; Gan, Y.H.; Liu, H.; Chen, E.S. cGAS-STING pathway in oncogenesis and cancer therapeutics. Oncotarget 2020, 11, 2930–2955. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, K.J.; Carroll, P.; Martin, C.A.; Murina, O.; Fluteau, A.; Simpson, D.J.; Olova, N.; Sutcliffe, H.; Rainger, J.K.; Leitch, A.; et al. cGAS surveillance of micronuclei links genome instability to innate immunity. Nature 2017, 548, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Riley, J.S.; Quarato, G.; Cloix, C.; Lopez, J.; O’Prey, J.; Pearson, M.; Chapman, J.; Sesaki, H.; Carlin, L.M.; Passos, J.F.; et al. Mitochondrial inner membrane permeabilisation enables mtDNA release during apoptosis. EMBO J. 2018, 37, e99238. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Wang, H.; Ren, J.; Chen, Q.; Chen, Z.J. cGAS is essential for cellular senescence. Proc. Natl. Acad. Sci. USA 2017, 114, E4612–E4620. [Google Scholar] [CrossRef]
- Wu, M.Z.; Cheng, W.C.; Chen, S.F.; Nieh, S.; O’Connor, C.; Liu, C.L.; Tsai, W.W.; Wu, C.J.; Martin, L.; Lin, Y.S.; et al. Izpisua Belmonte JC. miR-25/93 mediates hypoxia-induced immunosuppression by repressing cGAS. Nat. Cell Biol. 2017, 19, 1286–1296. [Google Scholar] [CrossRef]
- Song, S.; Peng, P.; Tang, Z.; Zhao, J.; Wu, W.; Li, H.; Shao, M.; Li, L.; Yang, C.; Duan, F.; et al. Decreased expression of STING predicts poor prognosis in patients with gastric cancer. Sci. Rep. 2017, 7, 39858. [Google Scholar] [CrossRef]
- Wu, S.; Zhang, Q.; Zhang, F.; Meng, F.; Liu, S.; Zhou, R.; Wu, Q.; Li, X.; Shen, L.; Huang, J.; et al. HER2 recruits AKT1 to disrupt STING signalling and suppress antiviral defence and antitumour immunity. Nat. Cell Biol. 2019, 21, 1027–1040. [Google Scholar] [CrossRef]
- Liu, L.; Feng, D.; Chen, G.; Chen, M.; Zheng, Q.; Song, P.; Ma, Q.; Zhu, C.; Wang, R.; Qi, W.; et al. Mitochondrial outer-membrane protein FUNDC1 mediates hypoxia-induced mitophagy in mammalian cells. Nat. Cell Biol. 2012, 14, 177–185. [Google Scholar] [CrossRef]
- Chen, R.; Liu, M.; Jiang, Q.; Meng, X.; Wei, J. The cyclic guanosine monophosphate synthase-stimulator of interferon genes pathway as a potential target for tumor immunotherapy. Front. Immunol. 2023, 14, 1121603. [Google Scholar] [CrossRef]
- Mender, I.; Zhang, A.; Ren, Z.; Han, C.; Deng, Y.; Siteni, S.; Li, H.; Zhu, J.; Vemula, A.; Shay, J.W.; et al. Telomere stress potentiates STING-dependent anti-tumor immunity. Cancer Cell 2020, 38, 400–411.e6. [Google Scholar] [CrossRef]
- Storozynsky, Q.; Hitt, M.M. The Impact of Radiation-Induced DNA Damage on cGAS-STING-Mediated Immune Responses to Cancer. Int. J. Mol. Sci. 2020, 21, 8877. [Google Scholar] [CrossRef]
- Hou, Y.; Liang, H.; Rao, E.; Zheng, W.; Huang, X.; Deng, L.; Zhang, Y.; Yu, X.; Xu, M.; Mauceri, H.; et al. Non-canonical NF-κB Antagonizes STING Sensor-Mediated DNA Sensing in Radiotherapy. Immunity 2018, 49, 490–503.e4. [Google Scholar] [CrossRef]
- Tian, J.; Zhang, D.; Kurbatov, V.; Wang, Q.; Wang, Y.; Fang, D.; Wu, L.; Bosenberg, M.; Muzumdar, M.D.; Khan, S.; et al. 5-Fluorouracil efficacy requires anti-tumor immunity triggered by cancer-cell-intrinsic STING. EMBO J. 2021, 40, e106065. [Google Scholar] [CrossRef] [PubMed]
- Kitai, Y.; Kawasaki, T.; Sueyoshi, T.; Kobiyama, K.; Ishii, K.J.; Zou, J.; Akira, S.; Matsuda, T.; Kawai, T.D. DNA-Containing Exosomes Derived from Cancer Cells Treated with Topotecan Activate a STING-Dependent Pathway and Reinforce Antitumor Immunity. J. Immunol. 2017, 198, 1649–1659. [Google Scholar] [CrossRef] [PubMed]
- Skopelja-Gardner, S.; An, J.; Elkon, K.B. Role of the cGAS-STING pathway in systemic and organ-specific diseases. Nat. Rev. Nephrol. 2022, 18, 558–572. [Google Scholar] [CrossRef] [PubMed]
- Haggerty, T.J.; Dunn, I.S.; Rose, L.B.; Newton, E.E.; Martin, S.; Riley, J.L.; Kurnick, J.T. Topoisomerase inhibitors modulate expression of melanocytic antigens and enhance T cell recognition of tumor cells. Cancer Immunol. Immunother. 2011, 60, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Chelvanambi, M.; Fecek, R.J.; Taylor, J.L.; Storkus, W.J. STING agonist-based treatment promotes vascular normalization and tertiary lymphoid structure formation in the therapeutic melanoma microenvironment. J. Immunother. Cancer 2021, 9, e001906. [Google Scholar] [CrossRef]
- Cooper, B.T.; Chmura, S.J.; Luke, J.J.; Shiao, S.L.; Basho, R.K.; Iams, W.T.; Page, D.B.; Li, C.; Gregory, R.C.; Shaw, M.; et al. TAK-676 in combination with pembrolizumab after radiation therapy in patients (pts) with advanced non–small cell lung cancer (NSCLC), triple-negative breast cancer (TNBC), or squamous-cell carcinoma of the head and neck (SCCHN): Phase 1 study design. J. Clin. Oncol. 2022, 40 (Suppl. S16), TPS2698. [Google Scholar] [CrossRef]
- Muñoz, N.M.; Williams, M.; Dixon, K.; Dupuis, C.; McWatters, A.; Avritscher, R.; Manrique, S.Z.; McHugh, K.; Murthy, R.; Tam, A.; et al. Influence of injection technique, drug formulation and tumor microenvironment on intratumoral immunotherapy delivery and efficacy. J. Immunother. Cancer 2021, 9, e001800. [Google Scholar] [CrossRef]
- Garland, K.M.; Sheehy, T.L.; Wilson, J.T. Chemical and Biomolecular Strategies for STING Pathway Activation in Cancer Immunotherapy. Chem. Rev. 2022, 122, 5977–6039. [Google Scholar] [CrossRef]
- Emran, A.A.; Fisher, D.E. Dual Targeting with EZH2 Inhibitor and STING Agonist to Treat Melanoma. J. Investig. Dermatol. 2022, 142, 1004–1006. [Google Scholar] [CrossRef] [PubMed]
- Muto, P.; Pastore, F. Radiotherapy in the Adjuvant and Advanced Setting of CSCC. Dermatol. Pract. Concept. 2021, 11 (Suppl. 2), e2021168S. [Google Scholar] [CrossRef]
- Hayman, T.J.; Baro, M.; MacNeil, T.; Phoomak, C.; Aung, T.N.; Cui, W.; Leach, K.; Iyer, R.; Challa, S.; Sandoval-Schaefer, T.; et al. STING enhances cell death through regulation of reactive oxygen species and DNA damage. Nat. Commun. 2021, 12, 2327. [Google Scholar] [CrossRef] [PubMed]
- Shalhout, S.Z.; Emerick, K.S.; Kaufman, H.L.; Miller, D.M. Immunotherapy for Non-melanoma Skin Cancer. Curr. Oncol. Rep. 2021, 23, 125. [Google Scholar] [CrossRef] [PubMed]
- Buechner, S.A.; Wernli, M.; Harr, T.; Hahn, S.; Itin, P.; Erb, P. Regression of basal cell carcinoma by intralesional interferon-α treatment is mediated by CD95 (Apo-1/Fas)-CD95 ligand-induced suicide. J. Clin. Investig. 1997, 100, 2691–2696. [Google Scholar] [CrossRef] [PubMed]
- Zou, S.S.; Qiao, Y.; Zhu, S.; Gao, B.; Yang, N.; Liu, Y.J.; Chen, J. Intrinsic strategies for the evasion of cGAS-STING signaling-mediated immune surveillance in human cancer: How therapy can overcome them. Pharmacol. Res. 2021, 166, 105514. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, Y.; Chen, X.; Zhang, W.; Zhao, L.; Weng, L.; Tian, H.; Wu, Z.; Tan, X.; Ge, X.; et al. Deubiquitinase USP35 restrains STING-mediated interferon signaling in ovarian cancer. Cell Death Differ. 2021, 28, 139–155. [Google Scholar] [CrossRef]
- Liu, W.; Alameh, M.G.; Yang, J.F.; Xu, J.R.; Lin, P.J.; Tam, Y.K.; Weissman, D.; You, J. Lipid Nanoparticles Delivering Constitutively Active STING mRNA to Stimulate Antitumor Immunity. Int. J. Mol. Sci. 2022, 23, 14504. [Google Scholar] [CrossRef]
- Fukuda, K. Immune Regulation by Cytosolic DNA Sensors in the Tumor Microenvironment. Cancers 2023, 15, 2114. [Google Scholar] [CrossRef]
- Rossi, M.; Carboni, S.; Berardino-Besson, D.; Riva, E.; Santiago-Raber, M.L.; Belnoue, E.; Derouazi, M. STING Agonist Combined to a Protein-Based Cancer Vaccine Potentiates Peripheral and Intra-Tumoral T Cell Immunity. Front. Immunol. 2021, 12, 695056. [Google Scholar] [CrossRef]
- Harding, S.M.; Benci, J.L.; Irianto, J.; Discher, D.E.; Minn, A.J.; Greenberg, R.A. Mitotic progression following DNA damage enables pattern recognition within micronuclei. Nature 2017, 548, 466–470. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Liang, H.; Xu, M.; Yang, X.; Burnette, B.; Arina, A.; Li, X.D.; Mauceri, H.; Beckett, M.; Darga, T.; et al. STING-dependent cytosolic DNA sensing promotes radiation-induced type I interferon-dependent antitumor immunity in immunogenic tumors. Immunity 2014, 41, 543–852. [Google Scholar] [CrossRef] [PubMed]
- Diamond, J.M.; Vanpouille-Box, C.; Spada, S.; Rudqvist, N.P.; Chapman, J.R.; Ueberheide, B.M.; Pilones, K.A.; Sarfraz, Y.; Formenti, S.C.; Demaria, S. Exosomes Shuttle TREX1-Sensitive IFN-Stimulatory dsDNA from Irradiated Cancer Cells to DCs. Cancer Immunol. Res. 2018, 6, 910–920. [Google Scholar] [CrossRef] [PubMed]
- Ludgate, C.M. Optimizing Cancer Treatments to Induce an Acute Immune Response: Radiation Abscopal Effects, PAMPs, and DAMPs. Clin. Cancer Res. 2012, 18, 4522–4525. [Google Scholar] [CrossRef]
- Baird, J.R.; Friedman, D.; Cottam, B.; Dubensky, T.W., Jr.; Kanne, D.B.; Bambina, S.; Bahjat, K.; Crittenden, M.R.; Gough, M.J. Radiotherapy Combined with Novel STING-Targeting Oligonucleotides Results in Regression of Established Tumors. Cancer Res. 2016, 76, 50–61. [Google Scholar] [CrossRef]
- Fu, J.; Kanne, D.B.; Leong, M.; Glickman, L.H.; McWhirter, S.M.; Lemmens, E.; Mechette, K.; Leong, J.J.; Lauer, P.; Liu, W.; et al. STING agonist formulated cancer vaccines can cure established tumors resistant to PD-1 blockade. Sci. Transl. Med. 2015, 7, 283ra52. [Google Scholar] [CrossRef]
- Xu, M.M.; Pu, Y.; Han, D.; Shi, Y.; Cao, X.; Liang, H.; Chen, X.; Li, X.D.; Deng, L.; Chen, Z.J.; et al. Dendritic Cells but Not Macrophages Sense Tumor Mitochondrial DNA for Cross-priming through Signal Regulatory Protein α Signaling. Immunity 2017, 47, 363–373.e5. [Google Scholar] [CrossRef]
- Woo, S.R.; Fuertes, M.B.; Corrales, L.; Spranger, S.; Furdyna, M.J.; Leung, M.Y.; Duggan, R.; Wang, Y.; Barber, G.N.; Fitzgerald, K.A.; et al. STING-dependent cytosolic DNA sensing mediates innate immune recognition of immunogenic tumors. Immunity 2014, 41, 830–842. [Google Scholar] [CrossRef]
- Wang, H.; Hu, S.; Chen, X.; Shi, H.; Chen, C.; Sun, L.; Chen, Z.J. cGAS is essential for the antitumor effect of immune checkpoint blockade. Proc. Natl. Acad. Sci. USA 2017, 114, 1637–1642. [Google Scholar] [CrossRef]
- Liu, X.; Pu, Y.; Cron, K.; Deng, L.; Kline, J.; Frazier, W.A.; Xu, H.; Peng, H.; Fu, Y.X.; Xu, M.M. CD47 blockade triggers T cell-mediated destruction of immunogenic tumors. Nat. Med. 2015, 21, 1209–1215. [Google Scholar] [CrossRef]
- Foote, J.B.; Kok, M.; Leatherman, J.M.; Armstrong, T.D.; Marcinkowski, B.C.; Ojalvo, L.S.; Kanne, D.B.; Jaffee, E.M.; Dubensky, T.W., Jr.; Emens, L.A. A STING Agonist Given with OX40 Receptor and PD-L1 Modulators Primes Immunity and Reduces Tumor Growth in Tolerized Mice. Cancer Immunol. Res. 2017, 5, 468–479. [Google Scholar] [CrossRef] [PubMed]
- National Library of Medicine (U.S.). (2019, August–2021, June). A Phase 2 Efficacy and Safety Trial of ADU-S100 and Pembrolizumab in Adults with Head and Neck Cancer. Identifier NCT03937141. Available online: https://classic.clinicaltrials.gov/ct2/show/NCT03937141?recrs=abdefghim&cond=ADU-S100&draw=2&rank=1 (accessed on 9 September 2023).
- National Library of Medicine (U.S.). (2019, September–Present). A Phase 1a/1b Dose-escalation Study of Intravenously Administered SB 11285 Alone and in Combination with Atezolizumab in Patients with Advanced Solid Tumors. Identifier NCT04096638. Available online: https://classic.clinicaltrials.gov/ct2/show/NCT04096638 (accessed on 9 September 2023).
- National Library of Medicine (U.S.). (2020, March–2022, September). A Phase 2 Study in First Line Metastatic or Unresectable, Recurrent Head and Neck Squamous Cell Carcinoma to Evaluate Intratumoral MK-1454 in Combination with IV Pembrolizumab vs IV Pembrolizumab Monotherapy Identifier. NCT04220866. Available online: https://classic.clinicaltrials.gov/ct2/show/study/NCT04220866 (accessed on 9 September 2023).
- Zandberg, D.P.; Ferris, R.; Laux, D.; Mehra, R.; Nabell, L.; Kaczmar, J.; Gibson, M.K.; Kim, Y.J.; Neupane, P.; Bauman, J.; et al. 71P A phase II study of Adu-S100 in combination with pembrolizumab in adult patients with PD-L1+ recurrent or METASTATIC HNSCC: Preliminary safety, efficacy and PK/PD Results. Ann. Oncol. 2020, 31, 1446–1447. [Google Scholar] [CrossRef]

{kind=link}
| Agonist | Indications | Recruitment Status | Phase | Route | Co-Therapy | Results | NCT Number |
|---|---|---|---|---|---|---|---|
| ADU-S100 | PD-L1 positive recurrent or metastatic HNSCC | Terminated | II | I.T. | I.V. infusions of pembrolizumab | No significant anti-tumor response was observed | NCT03937141 |
| SB 11285 | Melanoma, HNSCC, and advanced solid tumors | Active | I | I.V. | Dose-escalation study: administered as a monotherapy, then combination with atezolizumab | N/A | NCT04096638 |
| MK-1454 | PD-L1 positive recurrent or metastatic HNSCC | Completed | II | I.T. | I.V. infusions of pembrolizumab | No study results posted | NCT04220866 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oscherwitz, M.; Jiminez, V.; Terhaar, H.; Yusuf, N. Modulation of Skin Cancer by the Stimulator of Interferon Genes. Genes 2023, 14, 1794. https://doi.org/10.3390/genes14091794
Oscherwitz M, Jiminez V, Terhaar H, Yusuf N. Modulation of Skin Cancer by the Stimulator of Interferon Genes. Genes. 2023; 14(9):1794. https://doi.org/10.3390/genes14091794
Chicago/Turabian StyleOscherwitz, Max, Victoria Jiminez, Hanna Terhaar, and Nabiha Yusuf. 2023. "Modulation of Skin Cancer by the Stimulator of Interferon Genes" Genes 14, no. 9: 1794. https://doi.org/10.3390/genes14091794
APA StyleOscherwitz, M., Jiminez, V., Terhaar, H., & Yusuf, N. (2023). Modulation of Skin Cancer by the Stimulator of Interferon Genes. Genes, 14(9), 1794. https://doi.org/10.3390/genes14091794