
Molecular Characterization of the Acyl-CoA-Binding Protein Genes Reveals Their Significant Roles in Oil Accumulation and Abiotic Stress Response in Cotton


Abstract
1. Introduction
2. Materials and Methods
2.1. Identification of Cotton ACBP Genes
2.2. Phylogenic, Gene Structure, and Conserved Motif Analyses
2.3. Chromosomal Mapping and Gene Duplication Analyses
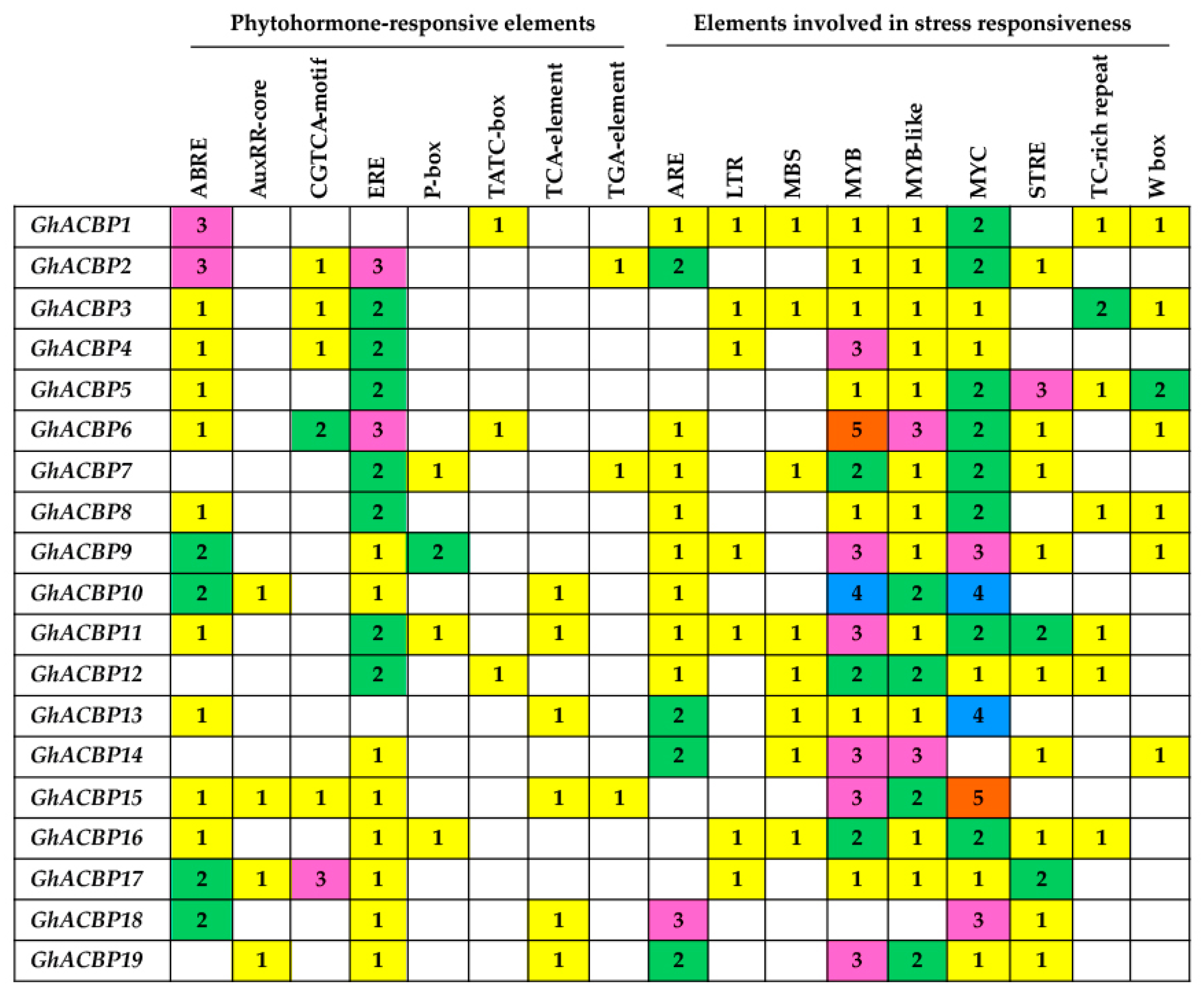
2.4. Promoter Analysis for Cis-Acting Regulatory Elements
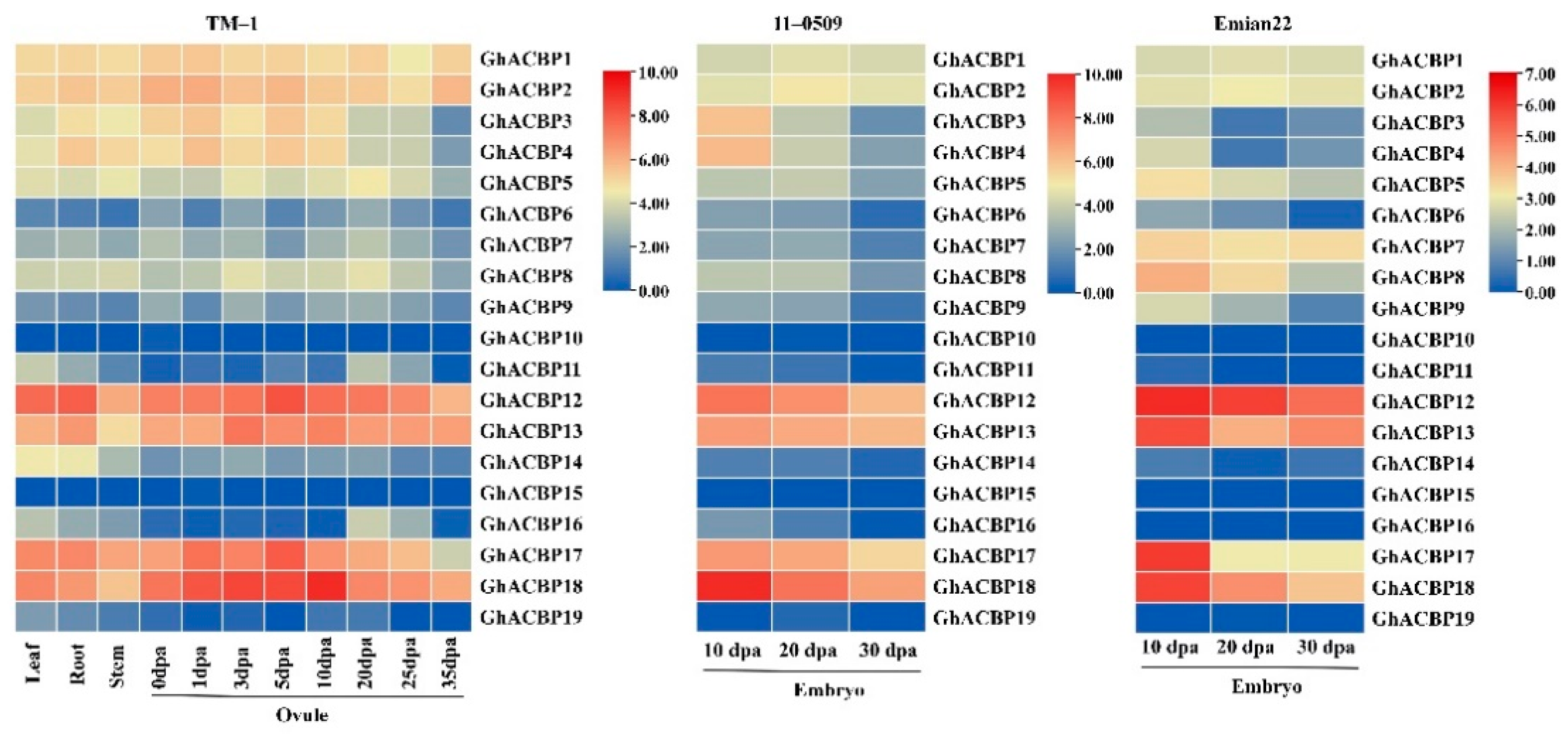
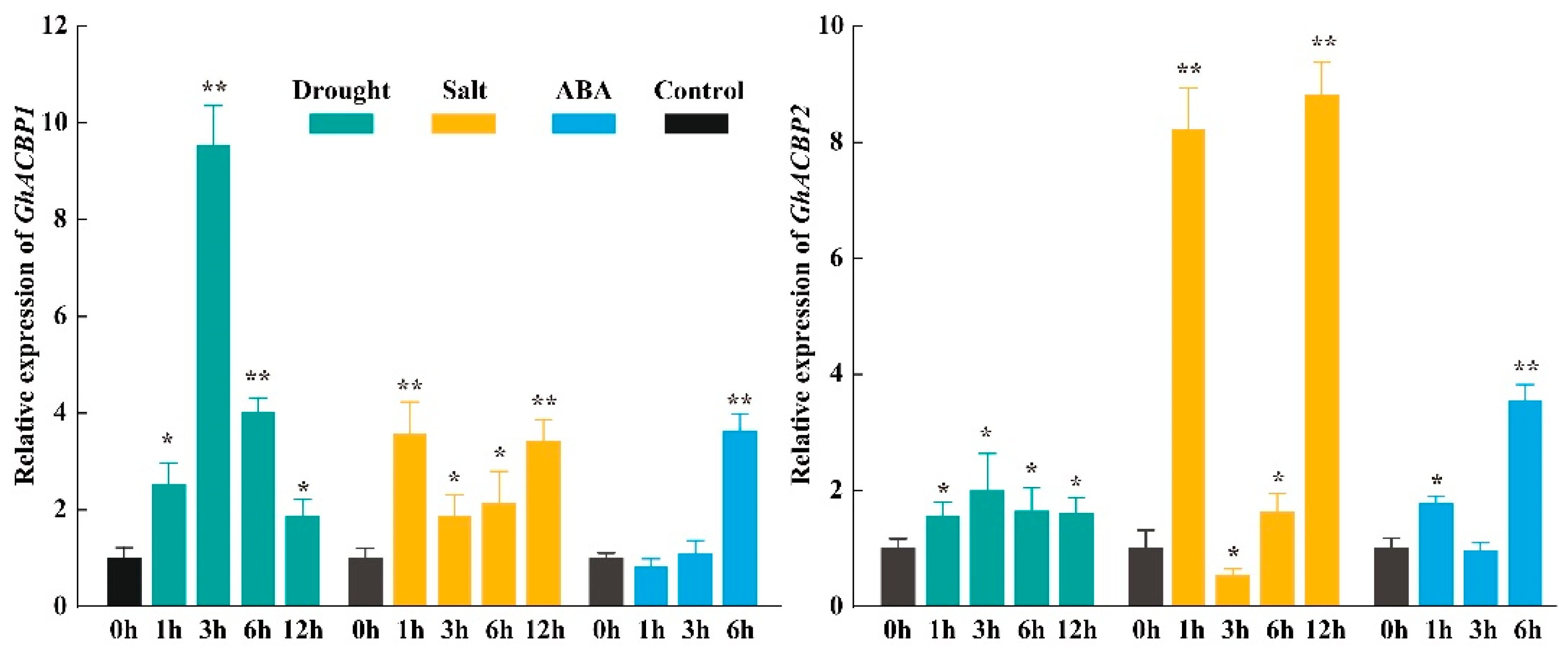
2.5. Expression Profile Analysis of GhACBP Genes
3. Results
3.1. Identification and Phylogenetic Analysis of Gossypium ACBP Genes
3.2. Gene Structure and Conserved Motif Analysis of Cotton ACBP Genes
3.3. Genomic Localization and Gene Duplication Analysis of Cotton ACBP Genes
3.4. Cis-Acting Regulatory Analysis of GhACBP Genes’ Promoters
3.5. Expression Pattern Analysis of GhACBP Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Raboanatahiry, N.; Wang, B.; Yu, L.; Li, M. Functional and structural diversity of acyl-CoA binding proteins in oil crops. Front. Genet. 2018, 9, 182. [Google Scholar] [CrossRef]
- Azlan, N.S.; Guo, Z.H.; Yung, W.S.; Wang, Z.; Lam, H.M.; Lung, S.C.; Chye, M.L. In silico analysis of acyl-CoA-binding protein expression in soybean. Front. Plant Sci. 2021, 12, 646938. [Google Scholar] [CrossRef] [PubMed]
- Ling, J.; Lin, L.; Xie, H.; Zheng, Y.; Wan, X. Genome-wide identification of acyl-CoA binding proteins and possible functional prediction in legumes. Front. Genet. 2023, 13, 1057160. [Google Scholar] [CrossRef]
- Hills, M.J.; Dann, R.; Lydiate, D.; Sharpe, A. Molecular cloning of a cDNA from Brassica napus L. for a homologue of acyl-CoA-binding protein. Plant Mol. Biol. 1994, 25, 917–920. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.P.; Johnson, P.; Rawsthorne, S.; Hills, M.J. Expression and properties of acyl-CoA binding protein from Brassica napus. Plant Physiol. Biochem. 1998, 36, 629–635. [Google Scholar] [CrossRef]
- Xiao, S.; Chye, M.L. An Arabidopsis family of six acyl-CoA-binding proteins has three cytosolic members. Plant Physiol. Bioch. 2009, 47, 479–484. [Google Scholar] [CrossRef]
- Meng, W.; Su, Y.C.F.; Saunders, R.M.K.; Chye, M.L. The rice acyl-CoA-binding protein gene family: Phylogeny, expression and functional analysis. New Phytol. 2011, 189, 1170–1184. [Google Scholar] [CrossRef]
- Zhu, J.; Li, W.; Zhou, Y.; Pei, L.; Liu, J.; Xia, X.; Che, R.; Li, H. Molecular characterization, expression and functional analysis of acyl-CoA-binding protein gene family in maize (Zea mays). BMC Plant Biol. 2021, 21, 94. [Google Scholar] [CrossRef]
- Raboanatahiry, N.H.; Yin, Y.; Chen, L.; Li, M. Genome-wide identification and phylogenic analysis of kelch motif containing ACBP in Brassica napus. BMC Genom. 2015, 16, 512. [Google Scholar] [CrossRef] [PubMed]
- Yurchenko, O.P.; Nykiforuk, C.L.; Moloney, M.M.; Stahl, U.; Banas, A.; Stymne, S.; Weselake, R.J. A 10-kDa acyl-CoA-binding protein (ACBP) from Brassica napus enhances acyl exchange between acyl-CoA and phosphatidylcholine. Plant Biotechnol. J. 2009, 7, 602–610. [Google Scholar] [CrossRef]
- Chen, Q.F.; Xiao, S.; Qi, W.; Mishra, G.; Ma, J.; Wang, M.; Chye, M.L. The Arabidopsis acbp1acbp2 double mutant lacking acyl-CoA-binding proteins ACBP1 and ACBP2 is embryo lethal. New Phytol. 2010, 186, 843–855. [Google Scholar] [CrossRef]
- Du, Z.Y.; Chen, M.X.; Chen, Q.E.; Xiao, S.; Chye, M.L. Overexpression of Arabidopsis acyl-CoA-binding protein ACBP2 enhances drought tolerance. Plant Cell Environ. 2013, 36, 300–314. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Huang, G.; He, S.; Yang, Z.; Sun, G.; Ma, X.; Li, N.; Zhang, X.; Sun, J.; Liu, M.; et al. Resequencing of 243 diploid cotton accessions based on an updated A genome identifies the genetic basis of key agronomic traits. Nat. Genet. 2018, 50, 796–802. [Google Scholar] [CrossRef] [PubMed]
- Paterson, A.H.; Wendel, J.F.; Gundlach, H.; Guo, H.; Jenkins, J.; Jin, D.; Llewellyn, D.; Showmaker, K.C.; Shu, S.; Udall, J.; et al. Repeated polyploidization of Gossypium genomes and the evolution of spinnable cotton fibres. Nature 2012, 492, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Tu, L.; Yuan, D.; Zhu, D.; Shen, C.; Li, J.; Liu, F.; Pei, L.; Wang, P.; Zhao, G.; et al. Reference genome sequences of two cultivated allotetraploid cottons, Gossypium hirsutum and Gossypium barbadense. Nat. Genet. 2019, 51, 224–229. [Google Scholar] [CrossRef]
- Zhu, T.; Liang, C.; Meng, Z.; Sun, G.; Meng, Z.; Guo, S.; Zhang, R. CottonFGD: An integrated functional genomics database for cotton. BMC Plant Biol. 2017, 17, 101. [Google Scholar] [CrossRef] [PubMed]
- Blum, M.; Chang, H.Y.; Chuguransky, S.; Grego, T.; Kandasaamy, S.; Mitchell, A.; Nuka, G.; Paysan-Lafosse, T.; Qureshi, M.; Raj, S.; et al. The InterPro protein families and domains database: 20 years on. Nucleic Acids Res. 2021, 49, D344–D354. [Google Scholar] [CrossRef]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent updates, new developments and status in 2020. Nucleic Acids Res. 2020, D1, D458–D460. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef]
- Fu, M.; Chen, Y.; Li, H.; Wang, L.; Liu, R.; Liu, Z. Genome-wide identification and expression analyses of the cotton AGO genes and their potential roles in fiber development and stress response. Genes 2022, 13, 1492. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Zhang, T.; Hu, Y.; Jiang, W.; Fang, L.; Guan, X.; Chen, J.; Zhang, J.; Saski, C.A.; Scheffler, B.E.; Stelly, D.M.; et al. Sequencing of allotetraploid cotton (Gossypium hirsutum L. acc. TM-1) provides a resource for fiber improvement. Nat. Biotechnol. 2015, 33, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, Y.; Huang, Y.; Cui, Y.; Hua, J. Gene network of oil accumulation reveals expression profiles in developing embryos and fatty acid composition in upland cotton. J. Plant Physiol. 2018, 228, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Le, Y.; Zhang, R.; Li, X.; Lin, Z. A global survey of the gene network and key genes for oil accumulation in cultivated tetraploid cottons. Plant Biotechnol. J. 2021, 19, 1170–1182. [Google Scholar] [CrossRef]
- Chen, Y.; Fu, M.; Li, H.; Wang, L.; Liu, R.; Liu, Z. Genome-wide characterization of the UDP-glycosyltransferase gene family reveals their potential roles in leaf senescence in cotton. Int. J. Biol. Macromol. 2022, 222, 2648–2660. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chen, M.X.; Hu, T.H.; Xue, Y.; Zhu, F.Y.; Du, Z.Y.; Lo, C.; Chye, M.L. Arabidopsis acyl-coenzyme-A-binding protein ACBP1 interacts with AREB1 and mediates salt and osmotic signaling in seed germination and seedling growth. Environ. Exp. Bot. 2018, 156, 130–140. [Google Scholar] [CrossRef]
- Enikeev, A.G.; Mishutina, U.O. Physiological effects of rapeseed transformation with the acb gene as affected by the genetic vector structure. Russ. J. Plant Physiol. 2005, 52, 668–671. [Google Scholar] [CrossRef]
- Engeseth, N.J.; Pacovsky, R.S.; Newman, T.; Ohlrogge, J.B. Characterization of an acyl-CoA binding protein from Arabidopsis thaliana. Arch. Biochem. Biophys. 1996, 331, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Lung, S.C.; Lai, S.H.; Wang, H.; Zhang, X.; Liu, A.; Guo, Z.H.; Lam, H.M.; Chye, M.L. Oxylipin signaling in salt-stressed soybean is modulated by ligand-dependent interaction of class II acyl-CoA-binding proteins with lipoxygenase. Plant Cell 2022, 34, 1117–1143. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Chr. | Gene Location (5′-3′) | CDS (bp) | Protein (aa) | MW (kD) | pI | Exon No. |
|---|---|---|---|---|---|---|---|---|
| GaACBP1 | Ga09G2073 | Chr09 | 78,553,212–78,556,181 | 1107 | 368 | 40.08 | 4.28 | 6 |
| GaACBP2 | Ga06G0242 | Chr06 | 2,161,888–2,164,066 | 1032 | 343 | 37.70 | 4.00 | 5 |
| GaACBP3 | Ga11G0450 | Chr11 | 6,033,134–6,039,718 | 1953 | 650 | 71.28 | 5.31 | 18 |
| GaACBP4 | Ga11G0874 | Chr11 | 14,739,861–14,750,764 | 2028 | 675 | 73.66 | 4.90 | 18 |
| GaACBP5 | Ga13G1376 | Chr13 | 85,900,097–85,912,287 | 2031 | 676 | 74.27 | 5.60 | 18 |
| GaACBP6 | Ga13G2190 | Chr13 | 116,634,315–116,641,164 | 2016 | 671 | 73.30 | 4.86 | 18 |
| GaACBP7 | Ga07G0156 | Chr07 | 1,719,913–1,721,334 | 267 | 88 | 9.89 | 8.86 | 4 |
| GaACBP8 | Ga11G0343 | Chr11 | 4,369,004–4,370,524 | 270 | 89 | 10.00 | 5.89 | 4 |
| GaACBP9 | Ga11G0344 | Chr11 | 4,375,942–4,377,237 | 273 | 90 | 10.23 | 5.21 | 4 |
| GaACBP10 | Ga11G1841 | Chr11 | 85,515,304–85,516,624 | 861 | 286 | 31.74 | 4.34 | 4 |
| GaACBP11 | Ga12G2217 | Chr12 | 73,224,127–73,225,994 | 846 | 281 | 31.31 | 4.04 | 4 |
| GrACBP1 | Gorai.006G202200 | Chr06 | 45,862,534–45,866,605 | 1104 | 367 | 39.95 | 4.25 | 6 |
| GrACBP2 | Gorai.010G024700 | Chr10 | 1,965,619–1,968,441 | 1029 | 342 | 37.80 | 3.98 | 5 |
| GrACBP3 | Gorai.013G078800 | Chr13 | 10,499,480–10,502,223 | 729 | 242 | 27.61 | 9.46 | 5 |
| GrACBP4 | Gorai.007G298800 | Chr07 | 50,996,820–51,005,403 | 2037 | 678 | 74.17 | 4.86 | 18 |
| GrACBP5 | Gorai.007G342000 | Chr07 | 56,727,854–56,734,910 | 1953 | 650 | 71.19 | 4.99 | 18 |
| GrACBP6 | Gorai.013G125100 | Chr13 | 32,513,007–32,521,783 | 2031 | 676 | 74.17 | 5.71 | 18 |
| GrACBP7 | Gorai.013G205300 | Chr13 | 51,615,237–51,622,742 | 2016 | 671 | 73.30 | 4.90 | 18 |
| GrACBP8 | Gorai.001G014900 | Chr01 | 1,397,605–1,399,298 | 267 | 88 | 9.86 | 9.40 | 4 |
| GrACBP9 | Gorai.007G221500 | Chr07 | 25,732,138–25,734,387 | 861 | 286 | 31.71 | 4.22 | 4 |
| GrACBP10 | Gorai.007G349800 | Chr07 | 57,989,173–57,990,985 | 273 | 90 | 10.21 | 5.21 | 4 |
| GrACBP11 | Gorai.007G349900 | Chr07 | 57,996,246–5,799,7051 | 258 | 85 | 9.59 | 5.16 | 3 |
| GrACBP12 | Gorai.008G080300 | Chr08 | 15,569,425–15,572,020 | 906 | 301 | 33.51 | 4.07 | 4 |
| GbACBP1 | Gbar_A09G019530 | A09 | 71,712,892–71,716,687 | 1008 | 335 | 36.53 | 4.36 | 6 |
| GbACBP2 | Gbar_D09G019330 | D09 | 45,806,450–45,810,624 | 1104 | 367 | 39.95 | 4.28 | 6 |
| GbACBP3 | Gbar_A06G002170 | A06 | 2,339,193–2,342,347 | 1311 | 436 | 48.83 | 4.25 | 5 |
| GbACBP4 | Gbar_D06G002250 | D06 | 2,169,955–2,172,708 | 1029 | 342 | 37.85 | 4.02 | 5 |
| GbACBP5 | Gbar_A11G028440 | A11 | 100,646,756–100,653,463 | 1971 | 656 | 71.80 | 4.94 | 18 |
| GbACBP6 | Gbar_A11G031740 | A11 | 107,991,123–107,998,178 | 2010 | 669 | 73.45 | 5.08 | 18 |
| GbACBP7 | Gbar_A13G012150 | A13 | 75,687,001–75,714,191 | 2013 | 670 | 73.62 | 5.70 | 19 |
| GbACBP8 | Gbar_D11G032520 | D11 | 64,081,464–64,088,613 | 1959 | 652 | 71.56 | 5.07 | 18 |
| GbACBP9 | Gbar_D13G011800 | D13 | 33,826,642–33,835,279 | 2040 | 679 | 74.61 | 5.82 | 19 |
| GbACBP10 | Gbar_D13G019150 | D13 | 53,310,527–53,318,013 | 2040 | 679 | 74.106 | 4.712 | 19 |
| GbACBP11 | Gbar_A07G001530 | A07 | 1,702,904–1,704,442 | 267 | 88 | 9.89 | 8.86 | 4 |
| GbACBP12 | Gbar_A11G022440 | A11 | 59,361,384–59,363,924 | 999 | 332 | 37.41 | 4.41 | 5 |
| GbACBP13 | Gbar_A11G032690 | A11 | 109,782,889–109,784,801 | 273 | 90 | 10.21 | 5.18 | 4 |
| GbACBP14 | Gbar_A11G032700 | A11 | 109,789,213–109,791,374 | 270 | 89 | 10.02 | 5.90 | 4 |
| GbACBP15 | Gbar_A12G007880 | A12 | 27,624,133–27,626,733 | 852 | 283 | 31.53 | 4.10 | 4 |
| GbACBP16 | Gbar_D07G001530 | D07 | 1,536,362–1,539,288 | 267 | 88 | 9.80 | 9.48 | 4 |
| GbACBP17 | Gbar_D11G021460 | D11 | 26,688,623–26,691,070 | 861 | 286 | 31.75 | 4.25 | 4 |
| GbACBP18 | Gbar_D11G033470 | D11 | 65,738,849–65,740,669 | 273 | 90 | 10.21 | 5.21 | 4 |
| GbACBP19 | Gbar_D11G033480 | D11 | 65,745,816–65,747,314 | 270 | 89 | 10.02 | 5.90 | 4 |
| GbACBP20 | Gbar_D12G007830 | D12 | 15,705,344–15,707,870 | 903 | 300 | 33.51 | 4.10 | 4 |
| GhACBP1 | Ghir_A09G019320 | A09 | 75,596,866–75,600,857 | 1104 | 367 | 40.03 | 4.30 | 6 |
| GhACBP2 | Ghir_D09G018830 | D09 | 47,230,179–47,234,288 | 1104 | 367 | 39.92 | 4.28 | 6 |
| GhACBP3 | Ghir_A06G002230 | A06 | 2,488,919–2,492,044 | 1314 | 437 | 48.89 | 4.27 | 5 |
| GhACBP4 | Ghir_D06G002060 | D06 | 2,200,409–2,203,279 | 1029 | 342 | 37.85 | 4.00 | 5 |
| GhACBP5 | Ghir_A11G028840 | A11 | 111,279,451–111,286,281 | 2028 | 675 | 73.87 | 5.00 | 18 |
| GhACBP6 | Ghir_A11G032190 | A11 | 118,023,421–118,030,387 | 1953 | 650 | 71.26 | 5.00 | 18 |
| GhACBP7 | Ghir_A13G011410 | A13 | 73,709,513–73,722,200 | 2031 | 676 | 74.23 | 5.59 | 18 |
| GhACBP8 | Ghir_D11G029020 | D11 | 61,586,023–61,592,814 | 1824 | 607 | 66.57 | 4.70 | 17 |
| GhACBP9 | Ghir_D11G032680 | D11 | 67,415,479–67,422,462 | 1953 | 650 | 71.26 | 4.94 | 18 |
| GhACBP10 | Ghir_A07G001520 | A07 | 1,613,013–1,614,450 | 267 | 88 | 9.89 | 8.86 | 4 |
| GhACBP11 | Ghir_A11G023070 | A11 | 64,631,311–64,633,846 | 999 | 332 | 37.41 | 4.41 | 5 |
| GhACBP12 | Ghir_A11G033220 | A11 | 119,683,936–119,685,963 | 273 | 90 | 10.21 | 5.18 | 4 |
| GhACBP13 | Ghir_A11G033230 | A11 | 119,690,233–119,692,421 | 270 | 89 | 10.02 | 5.90 | 4 |
| GhACBP14 | Ghir_A12G007910 | A12 | 28,971,057–28,973,672 | 852 | 283 | 31.58 | 4.10 | 4 |
| GhACBP15 | Ghir_D07G001540 | D07 | 1,575,302–1,576,882 | 267 | 88 | 9.86 | 9.40 | 4 |
| GhACBP16 | Ghir_D11G021280 | D11 | 27,187,461–27,190,098 | 861 | 286 | 31.78 | 4.25 | 4 |
| GhACBP17 | Ghir_D11G033830 | D11 | 69,635,385–69,637,499 | 273 | 90 | 10.22 | 5.21 | 4 |
| GhACBP18 | Ghir_D11G033840 | D11 | 69,642,750–69,644,268 | 279 | 92 | 10.54 | 5.86 | 4 |
| GhACBP19 | Ghir_D12G009120 | D12 | 29,686,393–29,688,931 | 885 | 294 | 32.69 | 4.08 | 4 |
| AtACBP1 | AT5G53470 | Chr5 | 21,710,170–21,712,614 | 1017 | 338 | 37.53 | 4.25 | 6 |
| AtACBP2 | AT4G27780 | Chr4 | 13,847,549–13,849,934 | 1065 | 354 | 38.48 | 4.16 | 6 |
| AtACBP3 | AT4G24230 | Chr4 | 12,566,631–12,568,866 | 1095 | 364 | 39.53 | 3.88 | 4 |
| AtACBP4 | AT3G05420 | Chr3 | 1,561,671–1,567,336 | 2007 | 668 | 73.07 | 4.95 | 18 |
| AtACBP5 | AT5G27630 | Chr5 | 9,775,854–9,781,002 | 1947 | 648 | 71.01 | 6.27 | 18 |
| AtACBP6 | AT1G31812 | Chr1 | 11,410,766–11,412,233 | 279 | 92 | 10.39 | 4.91 | 4 |
| OsACBP1 | LOC_Os08g06550 | Chr8 | 3,698,312–3,700,553 | 276 | 91 | 10.14 | 4.87 | 4 |
| OsACBP2 | LOC_Os06g02490 | Chr6 | 860,905–862,569 | 276 | 91 | 10.25 | 4.69 | 4 |
| OsACBP3 | LOC_Os03g37960 | Chr3 | 21,082,861–21,084,238 | 468 | 155 | 17.67 | 9.06 | 1 |
| OsACBP4 | LOC_Os04g58550 | Chr4 | 34,810,479–34,813,527 | 1011 | 336 | 35.90 | 4.23 | 6 |
| OsACBP5 | LOC_Os03g14000 | Chr3 | 7,591,868–7,597,447 | 1710 | 569 | 61.22 | 3.99 | 5 |
| OsACBP6 | LOC_Os03g61930 | Chr3 | 35,105,143–35,112,533 | 1971 | 656 | 71.54 | 5.05 | 18 |
| Duplicated Pair | Duplicated Type | Ka | Ks | Ka/Ks |
|---|---|---|---|---|
| GhACBP1/GhACBP2 | Segmental | 0.011 | 0.038 | 0.281 |
| GhACBP3/GhACBP4 | Segmental | 0.021 | 0.019 | 1.101 |
| GhACBP5/GhACBP6 | Segmental | 0.109 | 0.444 | 0.244 |
| GhACBP5/GhACBP7 | Segmental | 0.080 | 0.394 | 0.204 |
| GhACBP5/GhACBP8 | Segmental | 0.011 | 0.036 | 0.302 |
| GhACBP5/GhACBP9 | Segmental | 0.110 | 0.445 | 0.247 |
| GhACBP6/GhACBP8 | Segmental | 0.110 | 0.475 | 0.231 |
| GhACBP6/GhACBP9 | Segmental | 0.017 | 0.038 | 0.452 |
| GhACBP7/GhACBP8 | Segmental | 0.080 | 0.412 | 0.194 |
| GhACBP8/GhACBP9 | Segmental | 0.112 | 0.475 | 0.236 |
| GhACBP10/GhACBP15 | Segmental | 0.025 | 0.073 | 0.335 |
| GhACBP11/GhACBP14 | Segmental | 0.238 | 0.499 | 0.478 |
| GhACBP11/GhACBP16 | Segmental | 0.050 | 0.085 | 0.590 |
| GhACBP12/GhACBP17 | Segmental | 0.019 | 0.018 | 1.071 |
| GhACBP14/GhACBP16 | Segmental | 0.202 | 0.440 | 0.459 |
| GhACBP12/GhACBP13 | Tandem | 0.102 | 0.299 | 0.341 |
| GhACBP17/GhACBP18 | Tandem | 0.219 | 0.369 | 0.592 |
| GbACBP1/GbACBP2 | Segmental | 0.046 | 0.072 | 0.638 |
| GbACBP3/GbACBP4 | Segmental | 0.019 | 0.029 | 0.642 |
| GbACBP5/GbACBP6 | Segmental | 0.105 | 0.440 | 0.238 |
| GbACBP5/GbACBP7 | Segmental | 0.077 | 0.393 | 0.196 |
| GbACBP5/GbACBP8 | Segmental | 0.108 | 0.445 | 0.243 |
| GbACBP5/GbACBP9 | Segmental | 0.077 | 0.380 | 0.203 |
| GbACBP5/GbACBP10 | Segmental | 0.079 | 0.368 | 0.216 |
| GbACBP6/GbACBP8 | Segmental | 0.016 | 0.040 | 0.391 |
| GbACBP7/GbACBP9 | Segmental | 0.013 | 0.054 | 0.235 |
| GbACBP7/GbACBP10 | Segmental | 0.085 | 0.390 | 0.219 |
| GbACBP9/GbACBP10 | Segmental | 0.087 | 0.375 | 0.233 |
| GbACBP11/GbACBP16 | Segmental | 0.025 | 0.036 | 0.683 |
| GbACBP12/GbACBP15 | Segmental | 0.240 | 0.500 | 0.480 |
| GbACBP12/GbACBP17 | Segmental | 0.050 | 0.080 | 0.624 |
| GbACBP13/GbACBP18 | Segmental | 0.014 | 0.018 | 0.801 |
| GbACBP15/GbACBP17 | Segmental | 0.204 | 0.441 | 0.462 |
| GbACBP13/GbACBP14 | Tandem | 0.102 | 0.299 | 0.341 |
| GbACBP18/GbACBP19 | Tandem | 0.107 | 0.380 | 0.282 |
| GaACBP3/GaACBP4 | Segmental | 0.107 | 0.436 | 0.245 |
| GaACBP3/GaACBP5 | Segmental | 0.119 | 0.428 | 0.279 |
| GaACBP4/GaACBP5 | Segmental | 0.081 | 0.386 | 0.210 |
| GaACBP10/GaACBP11 | Segmental | 0.217 | 0.494 | 0.439 |
| GaACBP5/GaACBP6 | Segmental | 0.083 | 0.384 | 0.217 |
| GaACBP8/GaACBP9 | Tandem | 0.096 | 0.353 | 0.273 |
| GrACBP9/GrACBP2 | Segmental | 0.457 | 1.588 | 0.287 |
| GrACBP4/GrACBP5 | Segmental | 0.110 | 0.467 | 0.235 |
| GrACBP4/GrACBP6 | Segmental | 0.075 | 0.387 | 0.194 |
| GrACBP4/GrACBP7 | Segmental | 0.078 | 0.379 | 0.206 |
| GrACBP5/GrACBP6 | Segmental | 0.116 | 0.448 | 0.260 |
| GrACBP5/GrACBP7 | Segmental | 0.116 | 0.428 | 0.270 |
| GrACBP6/GrACBP7 | Segmental | 0.083 | 0.368 | 0.225 |
| GrACBP10/GrACBP11 | Tandem | 0.113 | 0.311 | 0.364 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Fu, M.; Li, H.; Wang, L.; Liu, R.; Liu, Z. Molecular Characterization of the Acyl-CoA-Binding Protein Genes Reveals Their Significant Roles in Oil Accumulation and Abiotic Stress Response in Cotton. Genes 2023, 14, 859. https://doi.org/10.3390/genes14040859
Chen Y, Fu M, Li H, Wang L, Liu R, Liu Z. Molecular Characterization of the Acyl-CoA-Binding Protein Genes Reveals Their Significant Roles in Oil Accumulation and Abiotic Stress Response in Cotton. Genes. 2023; 14(4):859. https://doi.org/10.3390/genes14040859
Chicago/Turabian StyleChen, Yizhen, Mingchuan Fu, Hao Li, Liguo Wang, Renzhong Liu, and Zhanji Liu. 2023. "Molecular Characterization of the Acyl-CoA-Binding Protein Genes Reveals Their Significant Roles in Oil Accumulation and Abiotic Stress Response in Cotton" Genes 14, no. 4: 859. https://doi.org/10.3390/genes14040859
APA StyleChen, Y., Fu, M., Li, H., Wang, L., Liu, R., & Liu, Z. (2023). Molecular Characterization of the Acyl-CoA-Binding Protein Genes Reveals Their Significant Roles in Oil Accumulation and Abiotic Stress Response in Cotton. Genes, 14(4), 859. https://doi.org/10.3390/genes14040859