Abstract
Breast cancer is considered the most frequent cause of mortality from malignancy among females. Fibroblast growth factor receptor 2 (FGFR2) gene polymorphisms are highly related to the risk of breast cancer. However, no investigation has been carried out to determine the association of FGFR2 gene polymorphisms in the Bangladeshi population. Based on polymerase chain reaction–restriction fragment length polymorphism (PCR-RFLP), this study was performed to evaluate the association of FGFR2 (rs1219648, rs2420946, and rs2981582) variants in 446 Bangladeshi women (226 cases and 220 controls). A significant association of the FGFR2 rs1219648 variant with breast malignancy was reported in additive model 1 (aOR = 2.87, p < 0.0001), additive model 2 (aOR = 5.62, p < 0.0001), the dominant model (aOR = 2.87, p < 0.0001), the recessive model (aOR = 4.04, p < 0.0001), and the allelic model (OR = 2.16, p < 0.0001). This investigation also explored the significant association of the rs2981582 variant with the risk of breast cancer in additive model 2 (aOR = 2. 60, p = 0.010), the recessive model (aOR = 2.47, p = 0.006), and the allelic model (OR = 1.39, p = 0.016). However, the FGFR2 rs2420946 polymorphism showed no association with breast cancer except in the overdominant model (aOR = 0.62, p = 0.048). Furthermore, GTT (p < 0.0001) haplotypes showed a correlation with breast cancer risk, and all variants showed strong linkage disequilibrium. Moreover, in silico gene expression analysis showed that the FGFR2 level was upregulated in BC tissues compared to healthy tissues. This study confirms the association of FGFR2 polymorphisms with breast cancer risk.
1. Introduction
Breast cancer is one of the most prevalent types of malignancy. The number of newly diagnosed patients identified was more than two million, approximately 11.7% of total cancer cases worldwide, in 2020. According to a report by GLOBOCAN, it is the fifth leading cause of morbidity (684,996 deaths, 6.9%) in the world [1]. It is also considered the most common cancer in 159 nations out of 185 and the most common cause of death in 110 countries. In addition, breast cancer is gradually becoming a major concern in South Asian countries, including Bangladesh, where the incidence rate is 22.5 per 100,000 women [2]. According to prior research, limited awareness of and learning about breast cancer among the people of Bangladesh has increased the risk of developing breast cancer [3]. Several factors, including maturity, circulating hormone concentrations, hereditary factors, and environmental exposure, contribute to the susceptibility to BC; however, the exact reason behind this is unclear. Moreover, hereditary factors are considered the most crucial attributes in the genetic variation in susceptible genes of approximately 5–10% of all breast cancer patients [4]. In addition, genome-wide association studies have found that numerous breast cancer-related genes and single-nucleotide polymorphisms (SNPs) are linked to the occurrence and progression of breast cancer [5].
Fibroblast growth factor receptor 2 (FGFR2) is a membrane-associated receptor tyrosine kinase responsible for transmitting a signal from fibroblast growth factors (FGFs). It is mapped on chromosome 10q26 and has a minimum of 22 members that serve numerous functions in cell proliferation and differentiation, growth, survival, and cell death. However, mutation of this gene leads to the proliferation and survival of tumor cells but also could suppress tumor growth [6,7,8,9,10]. In addition, FGFR2 mutations enhance DNA damage signaling and p53-induced senescence [11]. Oncogenic FGFR2 triggering is induced by FGFR2 missense mutations surrounding the third Ig-like domain. In contrast, those interiors in the tyrosine kinase domain generate oncogenic FGFR2 activation owing to the acquisition of ligand sovereignty [12].
FGFR2 may be a plausible mediator of breast cancer risk due to its role in signaling, the incidence of FGFR2 gene amplification in breast carcinoma, and the presence of risk SNPs within its intron 13. Intron 2 of the FGFR2 gene, highly conserved in mammals, has numerous possible transcription factor binding sites, some of which are positioned adjacent to the variants and speculated to be most strongly related to breast cancer [13,14,15]. There is also evidence that the risk allele contributes to an upregulation in the expression of FGFR2 [16]. Therefore, the FGFR2 gene has been identified as a potential risk factor for breast cancer development due to the genetic variations in this gene. The FGFR2 gene polymorphisms (rs1219648 A > G, rs2420946 C > T, and rs2981582 C > T) are located on intron 2 of chromosome 10 at different positions. SNP rs1219648 A > G is located in Chr10:121586676, whereas rs2420946 C > T is positioned in Chr10:121591810 and rs2981582 C > T is mapped in Chr10:121592803. These variants are predicted to enhance histone marks and motif changing. In addition to these functions, rs1219648 A > G and rs2981582 C > T are also speculated to be involved in GRASP QTL hitting. Previous studies also frequently showed that rs1219648, rs2420946, and rs2981582 are linked with the development of breast cancer in different ethnicities [12,14,15,17,18]. However, their association with breast cancer is still ambiguous due to the differences in ethnicity, geographic characteristics, and other determinants.
The associations of these polymorphisms were found to be inconsistent in different ethnic groups; however, there is no such investigation on the Bangladeshi population. Considering this fact, the current case–control study was performed to clarify the potential relationship between breast cancer and three common FGFR2 polymorphisms (rs1219648, rs2420946, and rs2981582).
2. Materials and Methods
2.1. Study Population
This case–control study was conducted at the Pharmacogenomics and Molecular Biology Laboratory at Noakhali Science and Technology University, Bangladesh, and approved by the ethical committee of the National Institute of Cancer Research and Hospital (NICRH), Bangladesh (NICRH/Ethics/2019/446). Around 446 women participated in this study, including 226 breast cancer patients and 220 healthy individuals. A comprehensive questionnaire was used to collect patients’ demographic and clinicopathological data with informed written consent. Selection guidelines for healthy controls mentioned no indication of cancer and other diseases such as kidney, liver, and lung diseases. For the clinicopathological features of breast cancer patients, we mostly focused on age, marital status, BMI, histological type of cancer, progesterone receptor status, estrogen receptor status, human epidermal growth factor receptor 2 (HER2/neu) status, grade, tumor measurement, lymph node condition, nodal stage, and distant metastasis. All information was retrieved from the patient’s medical record with the assistance of a physician. The Helsinki Declaration and its further correction were followed during the study [19].
2.2. Sample Preparation
Approximately 3 mL of blood was collected from all participants and stored in an EDTA-Na2-containing sterile tube at −80 °C until DNA extraction. DNA extraction was conducted via an established method routinely used in our lab [20]. Genotyping of all extracted DNA was performed by PCR using the target DNA fragment. A micro-volume spectrophotometer measured the purity and concentration of all DNA at 260 nm and 280 nm (Genova Nano, Jenway, IL, USA). Primer Blast online-based software was utilized to design the primer sequences.
2.3. Genotyping
SNPs rs1219648, rs2420946, and rs2981582 were genotyped by using the PCR-based restriction fragment length polymorphism (RFLP) technique. A working mix was prepared for each sample by adding EmeraldAmp GT PCR Master Mix (2x) and two designed complementary primers (forward and reverse) at a suitable concentration. For PCR, 20 µL of this working mix and 1 µL of DNA sample were mixed in a PCR tube and then amplified at certain conditions. The gel electrophoresis (1% agarose) method was followed to analyze the desired PCR fragments. Digestion of PCR products of all three SNPs was performed with HinP1I (Thermo Fisher Scientific, MA, USA) by incubation at 37 °C overnight. Digestion conditions and expected fragment lengths of all SNPs are listed in Table 1. Finally, the digested fragments were stained with Ethidium Bromide (EtBr) and visualized on 1.3% agarose gel electrophoresis, where a 100 bp DNA ladder was used to measure the fragments more accurately. The reverse and forward primer sequences and their required PCR conditions with fragment lengths are given in Table 1 (Supplementary Figures S1–S3)

Table 1.
Primers, PCR, and digestion conditions with respective fragment sizes.
2.4. Statistical Analysis
The chi-square (χ2) test was used to evaluate the Hardy–Weinberg Equilibrium (Table S1), and an odds ratio (OR) with 95% confidence interval (CI) was used to examine the risk of breast cancer. A significant association was considered at p < 0.05. ORs were adjusted (aOR) for BMI, age, marital status, family history, smoking status, OCP history, age at menarche, age of menopause, and drinking status for all genetic models except the allele model. In addition, targeted variants were compared with several clinicopathological characteristics. Meanwhile, the p-value was corrected by using Bonferroni correction, where the significance level was measured at p < 0.017 [21,22]. Additionally, linkage disequilibrium (LD) and haplotypes of rs1219648, rs2981582, and rs2420946 for breast cancer risk were also determined by applying the SHEsis online application.
3. Results
3.1. Demographic and Clinicopathological Characteristics
Table 2 shows the detailed demographic and clinicopathological variables. As per the observation, breast cancer was more prevalent among married patients (97.35%) and those aged more than 35 years (69.47%). A slightly significant difference (p = 0.049) was observed between cases and controls in terms of family history. Most subjects were overweight (28.76) or obese (30.51%). In addition, invasive ductal carcinoma (57.08%) and grade 2 (57.96%) types of cancer were the most common among the participants. Moreover, a higher percentage of them were recognized with a positive nodal status (64.16%) and Mx metastasis condition (75.22%).
Table 2.
Demographic and clinicopathological features of breast cancer patients.
3.2. Association of FGFR2 Polymorphisms with Breast Cancer
Table 3 provides the genotype and haplotype-based association of the FGFR2 rs1219648, rs2420946, and rs2981582 polymorphisms with the risk of breast cancer in the Bangladeshi population. As presented, this study identified a greater percentage of homozygote genotypes of the FGFR2 rs1219648 variant in controls. In contrast, a higher percentage of heterozygotes and mutant homozygotes of this SNP was observed in patients. As a result, a significant association of these genotypes with breast carcinoma was identified in various association models, including additive model 1 (AG vs. AA: aOR = 2.87, 95% CI = 1.76–3.69, p < 0.0001), additive model 2 (GG vs. AA: aOR = 5.62, 95% CI = 2.52–12.54, p < 0.0001), the dominant model (AG + GG vs. AA: aOR = 2.87, 95% CI =1.76–4.69, p < 0.0001), the recessive model (GG vs. AA + AG: aOR = 4.04, 95% CI = 1.90–8.59, p = 0.0001), and the allelic model (A vs. G: OR = 2.16, 95% CI = 1.62–2.89, p < 0.0001). Unlike the first variant discussed above, a greater percentage of heterozygotes of the FGFR2 rs2420946 variant was observed in controls. However, no substantial relationship was observed in any genetic inheritance models for this polymorphism except the overdominant model (CT vs. CC + TT: aOR = 0.62, 95% CI= 0.39–1.0, p = 0.048). Again, mutant homozygotes of the rs2981582 variant were more prevalent in cases compared to healthy individuals. As is presented, this study illustrated the significant association of this SNP with an elevated risk of breast cancer in additive model 2 (TT vs. CC: aOR = 2.60, 95% CI = 1.25–5.37, p = 0.010), the recessive model (TT vs. CC + CT: aOR = 2.47, 95% CI = 1.13–4.69, p = 0.006), and the allelic model (C vs. T: OR = 1.39, 95% CI = 1.06–1.82, p = 0.016).
Table 3.
Genotype frequencies and association analysis of FGFR2 rs1219648, rs2420946, and rs2981582 polymorphisms with breast cancer.
Furthermore, the haplotype-based analysis revealed a significant association of the GTT, ATT, and ATC haplotypes with breast cancer. The GTT haplotype (OR = 2.51, 95% CI = 1.86–3.55, p < 0.0001) depicted an enhanced risk of breast cancer, whereas the ATT (OR = 0.18, 95% CI = 0.10–0.33, p < 0.0001) and ATC (OR = 0.13, 95% CI = 0.04–0.43, p = 0.0009) haplotypes showed a protective association. Moreover, this study also illustrated the linkage disequilibrium (LD) of all SNPs in the breast cancer cases and controls (Figure 1) that represented the LD for rs1219648 and rs2420946 (D’ = 0.931, r2 = 0.624); rs1219648 and rs2981582 (D’ = 0.777, r2 = 0.389), and rs2420946 and rs2981582 (D’ = 0.823, r2 = 0.606).
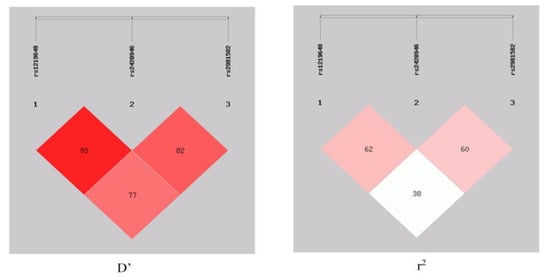
Figure 1.
LD block between rs1219648, rs2420946, and rs2981582 in the case–control respondents.
3.3. Association of FGFR2 Polymorphisms with Clinicopathological Variables
We investigated the linkage of breast cancer risk with FGFR2 gene polymorphisms considering various clinicopathological variables of patients, as illustrated in Table 4. A significant association of the FGFR2 rs1219648 variant with BC risk was found for the HER2 (+) status, grade 2 cancer, and positive lymph node status of patients. The genotype distribution of this variant increased the breast cancer risk in patients with HER2-positive status (OR = 2.13, 95% CI = 1.10–4.13, p = 0.025); however, it showed a protective association among those who had grade 2 cancer (OR = 0.38, 95% CI = 0.19–0.75, p = 0.006) and a positive lymph node status (OR = 0.55, 95% CI = 0.30–0.98, p = 0.043). In addition, patients identified with N1 nodal status showed a link with breast cancer (OR = 2.64, 95% CI = 1.42–4.93, p = 0.002) compared to the N0 nodal status of patients in terms of the rs2420946 polymorphism. Moreover, a protective correlation was also observed in patients with a positive lymph node status (OR = 0.40, 95% CI = 0.22–0.75, p = 0.004) in the case of the rs2981582 variant.
Table 4.
Associations of FGFR2 rs1219648, rs2420946, and rs2981582 polymorphisms with clinicopathological variables.
3.4. Analysis of In Silico Gene Expression
We used the GEPIA (http://gepia.cancer-pku.cn/, accessed on 20 November, 2022) and UALCAN (http://ualcan.path.uab.edu/, accessed on 20 November, 2022) databases to examine the FGFR2 mRNA expression levels in breast cancer. Breast cancer tissues had considerably more significant levels of FGFR2 mRNA expression than normal tissues (p < 0.01) (Figure 2). We also found significantly low expression of FGFR2 in Asians (Normal vs. Asian: p = 2.83 × 10−4) compared to Caucasians (Normal vs. Caucasian: p = 8.79 × 10−1). On the other hand, moderate expression was observed in African Americans (Normal vs. African American: p = 2.09 × 10−12) (Figure 3). All three races were compared with healthy individuals from the UALCAN database.
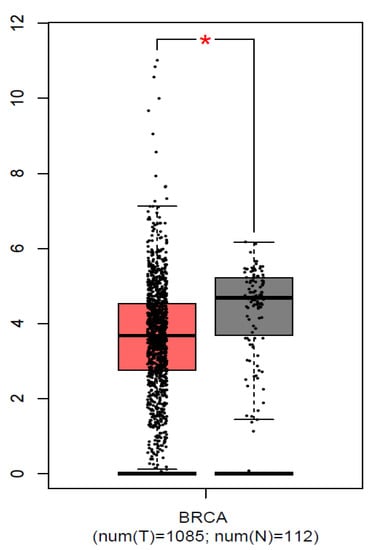
Figure 2.
Expression of FGFR2 in breast cancer tissues and normal tissues. Each bar indicates the average level of expression of FGFR2. Error bars are the standard deviation of the mean value. Data from the GEPIA database were extracted (http://gepia.cancer-pku.cn/, accessed on 20 November, 2022). * statistical significance (p < 0.01).
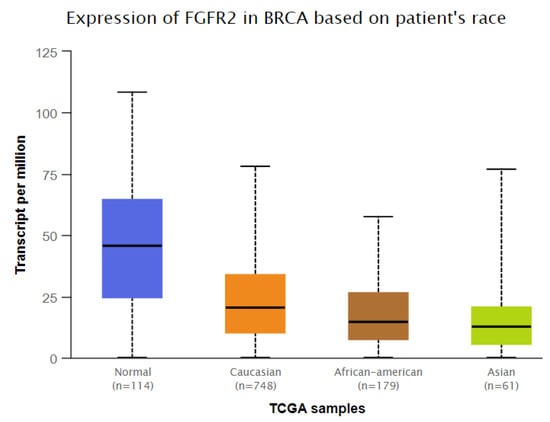
Figure 3.
Association of FGFR2 expression in different ethnic populations (Normal vs. Caucasian: p = 8.79 × 10−1, Normal vs. African American: p = 2.09 × 10−12, Normal vs. Asian: p = 2.83 × 10−4). Data from the UALCAN database were extracted (http://ualcan.path.uab.edu/, accessed on 20 November, 2022).
3.5. Genotype-Based FGFR2 mRNA Expression
The analysis of genotype-based FGFR2 mRNA expression for the studied SNPs (rs1219648, rs2420946, and rs2981582) from the Genotype-Tissue Expression (GTEx) database (http://www.gtexportal.org/, accessed on 20 November, 2022) is shown in Figure 4. The eQTL plots show that there was a statistically non-significant difference in the expression level of the FGFR2 rs1219648 polymorphism in breast tissues (p = 0.20) depending on the genotype. In the case of the FGFR2 rs2420946 and FGFR2 rs2981582 polymorphisms, the expression levels in breast tissues were also reported to be non-significant between the genotypes (p = 0.18 and p = 0.26, respectively).
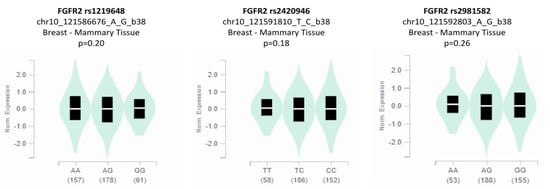
Figure 4.
Genotype-based mRNA expression of FGFR2 rs1219648, rs2420946, and rs2981582 polymorphisms.
4. Discussion
This case–control study was performed to clarify the association between breast cancer and three common FGFR2 polymorphisms (rs1219648, rs2420946, and rs2981582) in the Bangladeshi population. This is the first study in Bangladesh that has looked at the association of FGFR2 polymorphisms with breast cancer and observed a significant outcome. The increased incidence of cancer in recent years has had a devastating impact on the physical, mental, and social lives of human beings, making it one of the major problems of the century. The most common type of cancer in women worldwide is breast cancer, and this alarming condition has been associated with the most cancer-related deaths among women [23]. This is increasing day by day, and patients are being diagnosed at an earlier age. Though several causes have been identified behind this cancer, lifestyle and genetic variations are addressed as the main causes among different races. Previous investigations showed that genetic variations in many genes play a crucial role in breast cancer advancement [24]. However, the association between genetic risk factors in the etiology of breast carcinoma is still obscure [25,26].
The FGFR2 gene belongs to the tyrosine kinase receptor family, which is involved in multiple signaling pathways in tumorigenesis through apoptosis, differentiation, and cell growth [27]. Various studies revealed their relevance to breast cancer risk, where a polymorphism occurred in the FGFR2 intron 2 location and modulated the interaction of two transcription factors named Oct-1/Runx2 and C/EBPb. Consequently, increased FGFR2 gene expression in both cell lines and breast tissues occurred [16]. As is observed, the present study also found a significant association of the FGFR2 rs1219648 polymorphism with breast cancer risk in all determined genetic models, including additive model 1 (aOR = 2.87, p < 0.0001), additive model 2 (aOR = 5.62, p < 0.0001), the dominant model (aOR = 2.87, p < 0.0001), the recessive model (OR = 4.04, p = 0.0001), and alleles (OR = 2.16, p < 0.0001). Studies conducted in different ethnic and regional populations, such as North Indian, American, Turkish, Arab, and Kazakhstani populations, also revealed the association of rs1219648 with BC risk [28,29,30].
In addition, our study demonstrated the significant relationship between the rs2981582 variant and breast cancer in terms of some genetic models, including additive model 2 (aOR = 2.60, p = 0.010), the recessive model (aOR = 2.47, p = 0.006), and allele models (OR = 1.39, p = 0.016). Siddiqui et al. found an association of the TT allele of the FGFR2 rs2981582 polymorphism with an increased BC risk in North Indian women [28]. Their study also reported the linkage of this variant with breast cancer risk in the premenopausal patient, where the T allele showed a stronger association with ER (+) and PR (+) clinicopathology history. The FGFR2 rs2420946 polymorphism showed no significant association with breast cancer except in the overdominant model (aOR = 0.62, p = 0.048). A previous study conducted among Jewish and Arab Israeli BC populations also described the genetic variations of the FGFR2 gene (including rs2981582 and rs2420946) as a risk factor for breast cancer [17].
Furthermore, several case–control studies were conducted in Mexican, West Siberian, South American, Chinese, Iranian, Australian Caucasian, and Jordanian Arab women to determine the correlation between FGFR2 gene polymorphisms and breast cancer vulnerability [18,31,32,33,34,35,36]. They found that genetic variants in FGFR2 (rs2981582 C > T, rs1219648 A > G, and rs2420946 C > T) were associated with breast cancer risk. Additionally, Liang Jie et al. demonstrated that breast cancer in Chinese women mainly occurred in the premenopausal stage [18]. However, another study with Chinese Han women showed the opposite result. Their findings showed that polymorphisms such as rs2420946, rs2981582, and rs1219648 were significantly linked with breast cancer risk in postmenopausal patients but not in premenopausal subjects [37]. They observed no correlation between hormone receptor status (estrogen and progesterone) and breast cancer risk.
Liang et al. conducted a case–control study in Southern Han Chinese women and confirmed the association of genetic variation of the FGFR2 gene with the risk of breast cancer development [38]. Moreover, they stated that the association could differ with the variation of intrinsic subtypes. However, they found no significant correlations between FGFR2 polymorphisms and ER/PR/HER2 subtypes of breast cancer [38]. Our study also found no significant association of ER/PR/HER2 hormone receptors with breast cancer susceptibility in the case of rs2981582 and rs2420946. Notably, a significant association of HER2 (+) receptor (p = 0.025) with BC risk was observed in the case of the rs1219648 variant. In addition, this variant also showed an increased risk of breast cancer with grade 2 cancer (p = 0.006) and positive lymph nodes (p = 0.043) in patients. Meanwhile, rs2420946 and rs2981582 only showed an association with N1 nodal status (p = 0.002) and positive lymph nodes (p = 0.004) accordingly.
Furthermore, the haplotype-based analysis demonstrated a significant association of GTT, ATT, and ATC haplotypes with breast cancer. In addition, this study illustrated the LD of all SNPs in the breast cancer cases and controls, with the LD between rs1219648 and rs2420946 (D’ = 0.931, r2 = 0.624); rs1219648 and rs2981582 (D’ = 0.777, r2 = 0.389), and rs2420946 and rs2981582 (D’ = 0.823, r2 = 0.606). The GEPIA and UALCAN databases revealed that breast cancer tissues possess greater levels of FGFR2 mRNA expression than normal tissues (p < 0.01). Significantly lower expression of FGFR2 in Asians (p = 2.83 × 10−4) was observed compared to Caucasians (p = 8.79 × 10−1). On the other hand, moderate expression was observed in African Americans (p = 2.09 × 10−12).
Notably, there were some limitations in our study that must be acknowledged and overcome in future research. Firstly, we only investigated three available SNPs of the FGFR2 gene other than novel SNPs. Secondly, the comparatively small sample size of participants does not represent the whole scenario of all breast cancer patients in Bangladesh. Thirdly, there may be other factors, such as gene–environment interactions, which were not evaluated. Despite these limitations, this study has provided a clear indication of the relationship between FGFR2 gene polymorphisms and the risk of breast cancer development in the Bangladeshi population.
5. Conclusions
Our study found that FGFR2 genetic polymorphisms are significantly associated with breast cancer in Bangladeshi women. Different clinicopathological variables, such as HER2 status, cancer grade, and lymph node status, may have an impact on genetic polymorphisms that may be collectively associated with breast cancer risk. More research with a large sample size is suggested to confirm the findings of this study.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/genes14040819/s1, Figure S1: Restriction Endonuclease (HinP1I) digestion fragments of FGFR2 rs2981582 (1.3% agarose gel). Lane 1 indicates 100bp ladder; lanes 2-4,6,8,9 indicate TT genotype; lanes 10,11 indicate CT genotypes; lanes 5,7,12-16 indicate CC genotype; Figure S2: Restriction Endonuclease (HinP1I) digestion fragments of FGFR2 rs2420946 (1.3% agarose gel). Lane 1 indicates 100bp ladder; lanes 2,7,8,10,16 indicate CT genotype; lanes 3,4 indicate TT genotypes; lanes 5,6,9,11-15 indicate CC genotype; Figure S3: Restriction Endonuclease (HinP1I) digestion fragments of FGFR2 rs1219648 (1.3% agarose gel). Lane 1 indicates 100bp ladder; lanes 2-4,8-10,12,13 indicate GG genotype; lanes 5,11,14 indicate AG genotypes; lanes 7 indicate AA genotype.
Author Contributions
Conceptualization, M.S.I.; methodology, N.J., M.B., M.A.B., T.A. and M.S.H.; software, M.S.I., M.R.I. and T.A.; validation, N.J., M.B., M.A.A., T.A., K.K.B., M.S.H. and M.S.H.; formal analysis, N.J., M.B., M.A.A. and M.A.B.; investigation, N.J., M.B., T.A. and M.S.I.; resources, M.S.I. and H.S.A.; data curation, M.S.I.; writing—original draft preparation, N.J., M.A.A., M.A.B., M.S.H., M.R.I. and H.S.A.; writing—review and editing, M.S.I., M.A.B., H.S.A. and K.K.B.; visualization, M.S.I., M.A.B., M.S.H. and M.R.I.; supervision, M.S.I.; project administration, M.S.I. and M.B.; funding acquisition, M.S.I. All authors have read and agreed to the published version of the manuscript.
Funding
Research Cell: Noakhall Science and Technology University and National Science and Technology University (ID: NSTU/RC/19/C-84) and National Science and Technology (NST) Fellowship 2019, Ministry of Science and Technology, Bangladesh, have partially funded this research work.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Ethics Committee of the National Institute of Cancer Research and Hospital (NICRH) (protocol code NICRH/Ethics/20 l9/446 and date of approval 11 June 2019).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
We thank the authority of the National Institute of Cancer Research and Hospital, Dhaka, Bangladesh, for their cooperation and help during the collection of blood samples.
Conflicts of Interest
The authors declare no competing interests.
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Begum, S.; Mahmud, T.; Rahman, T.; Zannat, J.; Khatun, F.; Nahar, K.; Towhida, M.; Joarder, M.; Harun, A.; Sharmin, F. Knowledge, attitude and practice of Bangladeshi women towards breast cancer: A cross sectional study. Mymensingh Med. J. 2019, 28, 96–104. [Google Scholar] [PubMed]
- Alam, N.E.; Islam, M.S.; Ullah, H.; Molla, M.T.; Shifat, S.K.; Akter, S.; Aktar, S.; Khatun, M.M.; Ali, M.R.; Sen, T.C. Evaluation of knowledge, awareness and attitudes towards breast cancer risk factors and early detection among females in Bangladesh: A hospital based cross-sectional study. PLoS ONE 2021, 16, e0257271. [Google Scholar] [CrossRef] [PubMed]
- Barek, M.A.; Begum, M.; Noor, F.; Aziz, M.A.; Islam, M.S. The link between IL-6 rs2069840 SNP and cancer risk: Evidence from a systematic review and meta-analysis. Meta Gene 2021, 30, 100972. [Google Scholar] [CrossRef]
- Aziz, M.A.; Akter, T.; Sarwar, M.S.; Islam, M.S. The first combined meta-analytic approach for elucidating the relationship of circulating resistin levels and RETN gene polymorphisms with colorectal and breast cancer. Egypt. J. Med. Hum. Genet. 2022, 23, 27. [Google Scholar] [CrossRef]
- Touat, M.; Ileana, E.; Postel-Vinay, S.; André, F.; Soria, J.-C. Targeting FGFR signaling in cancer. Clin. Cancer Res. 2015, 21, 2684–2694. [Google Scholar] [CrossRef]
- Turner, N.; Grose, R. Fibroblast growth factor signalling: From development to cancer. Nat. Rev. Cancer 2010, 10, 116–129. [Google Scholar] [CrossRef]
- Zhao, Y.; Adjei, A.A. Targeting angiogenesis in cancer therapy: Moving beyond vascular endothelial growth factor. Oncologist 2015, 20, 660–673. [Google Scholar] [CrossRef]
- Eswarakumar, V.; Lax, I.; Schlessinger, J. Cellular signaling by fibroblast growth factor receptors. Cytokine Growth Factor Rev. 2005, 16, 139–149. [Google Scholar] [CrossRef]
- Bai, A.; Meetze, K.; Vo, N.Y.; Kollipara, S.; Mazsa, E.K.; Winston, W.M.; Weiler, S.; Poling, L.L.; Chen, T.; Ismail, N.S. GP369, an FGFR2-IIIb–Specific Antibody, Exhibits Potent Antitumor Activity against Human Cancers Driven by Activated FGFR2 Signaling In vivo Efficacy of GP369 in FGFR2-Amplified Tumors. Cancer Res. 2010, 70, 7630–7639. [Google Scholar] [CrossRef]
- Ota, S.; Zhou, Z.-Q.; Link, J.M.; Hurlin, P.J. The role of senescence and prosurvival signaling in controlling the oncogenic activity of FGFR2 mutants associated with cancer and birth defects. Hum. Mol. Genet. 2009, 18, 2609–2621. [Google Scholar] [CrossRef] [PubMed]
- Katoh, M. Cancer genomics and genetics of FGFR2. Int. J. Oncol. 2008, 33, 233–237. [Google Scholar] [PubMed]
- Fletcher, M.N.C.; Castro, M.A.A.; Wang, X.; de Santiago, I.; O’Reilly, M.; Chin, S.-F.; Rueda, O.M.; Caldas, C.; Ponder, B.A.J.; Markowetz, F.; et al. Master regulators of FGFR2 signalling and breast cancer risk. Nat. Commun. 2013, 4, 2464. [Google Scholar] [CrossRef] [PubMed]
- Hunter, D.J.; Kraft, P.; Jacobs, K.B.; Cox, D.G.; Yeager, M.; Hankinson, S.E.; Wacholder, S.; Wang, Z.; Welch, R.; Hutchinson, A. A genome-wide association study identifies alleles in FGFR2 associated with risk of sporadic postmenopausal breast cancer. Nat. Genet. 2007, 39, 870–874. [Google Scholar] [CrossRef] [PubMed]
- Easton, D.F.; Pooley, K.A.; Dunning, A.M.; Pharoah, P.D.; Thompson, D.; Ballinger, D.G.; Struewing, J.P.; Morrison, J.; Field, H.; Luben, R. Genome-wide association study identifies novel breast cancer susceptibility loci. Nature 2007, 447, 1087–1093. [Google Scholar] [CrossRef]
- Meyer, K.B.; Maia, A.-T.; O’Reilly, M.; Teschendorff, A.E.; Chin, S.-F.; Caldas, C.; Ponder, B.A.J. Allele-specific up-regulation of FGFR2 increases susceptibility to breast cancer. PLoS Biol. 2008, 6, e108. [Google Scholar] [CrossRef]
- Raskin, L.; Pinchev, M.; Arad, C.; Lejbkowicz, F.; Tamir, A.; Rennert, H.S.; Rennert, G.; Gruber, S.B. FGFR2 is a breast cancer susceptibility gene in Jewish and Arab Israeli populations. Cancer Epidemiol. Biomark. Prev. 2008, 17, 1060–1065. [Google Scholar] [CrossRef]
- Liang, J.; Chen, P.; Hu, Z.; Zhou, X.; Chen, L.; Li, M.; Wang, Y.; Tang, J.; Wang, H.; Shen, H. Genetic variants in fibroblast growth factor receptor 2 (FGFR2) contribute to susceptibility of breast cancer in Chinese women. Carcinogenesis 2008, 29, 2341–2346. [Google Scholar] [CrossRef]
- General Assembly of the World Medical, A. World Medical Association Declaration of Helsinki: Ethical principles for medical research involving human subjects. J. Am. Coll. Dent. 2014, 81, 14–18. [Google Scholar]
- Islam, M.S.; Ahmed, M.U.; Sayeed, M.S.B.; Al Maruf, A.; Mostofa, A.; Hussain, S.M.A.; Kabir, Y.; Daly, A.K.; Hasnat, A. Lung cancer risk in relation to nicotinic acetylcholine receptor, CYP2A6 and CYP1A1 genotypes in the Bangladeshi population. Clin. Chim. Acta 2013, 416, 11–19. [Google Scholar] [CrossRef]
- Goeman, J.J.; Solari, A. Multiple hypothesis testing in genomics. Stat. Med. 2014, 33, 1946–1978. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.F. p Value fetishism and use of the Bonferroni adjustment. Evid.-Based Ment. Health 2007, 10, 34–35. [Google Scholar] [CrossRef] [PubMed]
- Lichtenstein, P.; Holm, N.V.; Verkasalo, P.K.; Iliadou, A.; Kaprio, J.; Koskenvuo, M.; Pukkala, E.; Skytthe, A.; Hemminki, K. Environmental and heritable factors in the causation of cancer—Analyses of cohorts of twins from Sweden, Denmark, and Finland. N. Engl. J. Med. 2000, 343, 78–85. [Google Scholar] [CrossRef]
- Yager, J.D.; Davidson, N.E. Estrogen carcinogenesis in breast cancer. N. Engl. J. Med. 2006, 354, 270–282. [Google Scholar] [CrossRef] [PubMed]
- Rebbeck, T.R.; Mitra, N.; Wan, F.; Sinilnikova, O.M.; Healey, S.; McGuffog, L.; Mazoyer, S.; Chenevix-Trench, G.; Easton, D.F.; Antoniou, A.C. Association of type and location of BRCA1 and BRCA2 mutations with risk of breast and ovarian cancer. JAMA 2015, 313, 1347–1361. [Google Scholar] [CrossRef] [PubMed]
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef]
- Wesche, J.; Haglund, K.; Haugsten, E.M. Fibroblast growth factors and their receptors in cancer. Biochem. J. 2011, 437, 199–213. [Google Scholar] [CrossRef]
- Siddiqui, S.; Chattopadhyay, S.; Akhtar, M.S.; Najm, M.Z.; Deo, S.; Shukla, N.; Husain, S.A. A study on genetic variants of Fibroblast growth factor receptor 2 (FGFR2) and the risk of breast cancer from North India. PLoS ONE 2014, 9, e110426. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, H.; Lin, M.; Wang, Y. Association of FGFR2 and PI3KCA genetic variants with the risk of breast cancer in a Chinese population. Cancer Manag. Res. 2018, 10, 1305. [Google Scholar] [CrossRef]
- Andersen, S.W.; Trentham-Dietz, A.; Figueroa, J.D.; Titus, L.J.; Cai, Q.; Long, J.; Hampton, J.M.; Egan, K.M.; Newcomb, P.A. Breast cancer susceptibility associated with rs1219648 (fibroblast growth factor receptor 2) and postmenopausal hormone therapy use in a population-based United States study. Menopause 2013, 20, 354–358. [Google Scholar] [CrossRef]
- Michailidou, K.; Hall, P.; Gonzalez-Neira, A.; Ghoussaini, M.; Dennis, J.; Milne, R.L.; Schmidt, M.K.; Chang-Claude, J.; Bojesen, S.E.; Bolla, M.K. Large-scale genotyping identifies 41 new loci associated with breast cancer risk. Nat. Genet. 2013, 45, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Boyarskikh, U.A.; Zarubina, N.A.; Biltueva, J.A.; Sinkina, T.V.; Voronina, E.N.; Lazarev, A.F.; Petrova, V.D.; Aulchenko, Y.S.; Filipenko, M.L. Association of FGFR2 gene polymorphisms with the risk of breast cancer in population of West Siberia. Eur. J. Hum. Genet. 2009, 17, 1688–1691. [Google Scholar] [CrossRef] [PubMed]
- Jara, L.; Gonzalez-Hormazabal, P.; Cerceno, K.; Di Capua, G.A.; Reyes, J.M.; Blanco, R.; Bravo, T.; Peralta, O.; Gomez, F.; Waugh, E. Genetic variants in FGFR2 and MAP3K1 are associated with the risk of familial and early-onset breast cancer in a South-American population. Breast Cancer Res. Treat. 2013, 137, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, M.; Houshmand, M.; Froozan, S. Association of FGFR2 and TOX3 Genetic Variants with the Risk of Breast Cancer in Iranian Women. Arch. Breast Cancer 2018, 10, 118–121. [Google Scholar]
- Arif, K.M.T.; Bradshaw, G.; Nguyen, T.T.N.; Smith, R.A.; Okolicsanyi, R.K.; Youl, P.H.; Haupt, L.M.; Griffiths, L.R. Genetic Association Analysis Implicates Six MicroRNA-Related SNPs With Increased Risk of Breast Cancer in Australian Caucasian Women. Clin. Breast Cancer 2021, 21, e694–e703. [Google Scholar] [CrossRef] [PubMed]
- Al-Eitan, L.N.; Rababa’H, D.M.; Aman, H.A. The Associations of Common Genetic Susceptibility Variants with Breast Cancer in Jordanian Arabs: A Case-Control Study. Asian Pac. J. Cancer Prev. 2020, 21, 3045–3054. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-L.; Hu, X.-P.; Guo, W.-D.; Yang, L.; Dang, J.; Jiao, H.-Y. Case-control study on the fibroblast growth factor receptor 2 gene polymorphisms associated with breast cancer in Chinese Han women. J. Breast Cancer 2013, 16, 366–371. [Google Scholar] [CrossRef]
- Liang, H.; Yang, X.; Chen, L.; Li, H.; Zhu, A.; Sun, M.; Wang, H.; Li, M. Heterogeneity of breast cancer associations with common genetic variants in FGFR2 according to the intrinsic subtypes in Southern Han Chinese women. BioMed Res. Int. 2015, 2015, 626948. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).