Abstract
Mitochondrial DNA and nonrecombinant parts of Y-chromosome DNA are a great tool for looking at a species’ past. They are inherited for generations almost unaffected because they do not participate in recombination; thus, the time of occurrence of each mutation can be estimated based on the average mutation rate. Thanks to this, male and female haplogroups guide confirming events in the distant past (potential centers of domestication, settlement of areas, trade connections) as well as in modern breeding (crossbreeding, confirmation of paternity). This research focuses mainly on the development of domestic sheep and its post-domestication expansion, which has occurred through human trade from one continent to another. So far, five mitochondrial and five Y-chromosome haplogroups and dozens of their haplotypes have been detected in domestic sheep through studies worldwide. Mitochondrial DNA variability is more or less correlated with distance from the domestication center, but variability on the recombinant region of the Y chromosome is not. According to available data, central China shows the highest variability of male haplogroups and haplotypes.
1. Introduction
Domestic sheep (Ovis orientalis Linnaeus, 1758), together with domestic goat (Capra aegagrus hircus Linnaeus, 1758), were among the first livestock to be domesticated through several domestication events between the eleventh and eighth millennia BP [1,2,3]. Throughout almost ten millennia, domestic sheep have spread with the help of man to almost all continents, different climatic zones, and altitudes. This adaptability and production variability are naturally rooted in its genome. Indeed, most of this functional genetic diversity comes from wild ancestors in which they have already been segregated [4,5]. Scientists are increasingly seeking functional genes that cause this natural adaptability due to their potential use in marker-assisted selection [6,7,8].
The urial (Ovis vignei Blyth, 1841) was first considered the main ancestor of domestic sheep, and only after the number of chromosomes in individual related species of the genus Ovis was revealed, the scientific public leaned towards the theory of a single ancestor, the European mouflon (Ovis orientalis musimon Pallas, 1811) [9]. According to the latest findings supported by several mtDNA haplotype studies [10,11,12,13], the direct ancestor of the domestic sheep appears to be the Asian mouflon (Ovis gmelinii Gmelin, 1774), while a close relative of the sheep, the European mouflon, appears only to be a feralized remnant of the originally domesticated sheep. This statement was confirmed when the same retrotypes were found in the Corsican, Cypriot, and Sardinian mouflons as in primitive Nordic sheep breeds [14]. Modern breeds are characterized by a high frequency and fixation of the retrotype called enJSRV-18. In contrast, primitive populations, including the European mouflon, do not carry this retrotype. Instead, they either have a high frequency of enJSRV-7 or are generally deficient in insertional polymorphic enJSRVs, including enJSRV-7 [14].
Molecular genetic methods provide information today on the dispersion and genetic diversity of domestic sheep. Over the last few decades, significant progress has been made in the genomic sequencing of animals, including sheep [15]. However, many methods targeting specific sections of the genome are also used to study the genetic diversity of sheep: the study of haplotypes (mitochondrial and non-recombinant parts of the Y chromosome), autosomal microsatellite markers, and, most recently, single nucleotide polymorphisms, SNPs [16]. With the increasing amount of genetic information available, our information on the origin, development, adaptation mechanisms, and variability of livestock is becoming more accurate [15]. For the study of development and origin, information from Y or mt haplotypes of primitive national breeds, or even better directly from archaeological finds, is particularly valuable [17]. Indigenous breeds are not expected to have a larger proportion of newly introduced genes, as their development is closely linked to the development of ethnic groups, which usually still breed them in the traditional pastoral way in certain areas for many centuries and millennia [18,19,20].
Some studies even point to the possibility of using recombinant sections of gonosomes. Diversity on the X chromosome and autosomal chromosomes in wild and domestic sheep across continents revealed a decrease in the diversity of single nucleotide polymorphisms (SNPs) on the X chromosome compared to autosomes [21]. On the other hand, a smaller number of selective SNPs are found on the X chromosome, probably because most target loci and genes that are long-term are affected by human selection and are located on autosomes. Chessa et al. [22] focused on these loci and demonstrated that even in the functional regions of the sheep genome, there is considerable genetic variability, reflecting years of adaptation, natural or artificial selection, migration, and crossing. They can, therefore, also be used to study current biodiversity.
This review aims to summarize the current knowledge about the colonization dispersion of domestic sheep based on the two most used approaches to this issue, the study of the variability of mitochondrial and nonrecombinant Y DNA. The meta-analysis in the form of graphical outputs focused on recent local and transboundary breeds. Commercial or improved breeds (e.g., Texel) were not included in the dataset.
2. Mitochondrial Haplogroups and Haplotypes
MtDNA is inherited through the maternal lineage and, thus, lacks recombination. At the same time, it mutates five to ten times faster than nuclear DNA [23]. This may be due to a lack of repair mechanisms or the formation of free radicals during the phosphorylation process [24]. The hypervariable region of the mtDNA regulatory region is one of the most available and effective markers for population genetic studies, which allows monitoring of the maternal lineage of the gene pool and the related phylogenetic relationships, structure, and diversity of the population [25].
Assuming that humans take only part of the animal population from the original domestication center when colonizing new areas, mtDNA haplotype studies should logically reflect the geographical progression of the domestication of sheep. Thus, the greatest diversity of mitochondrial haplotypes in sheep can be expected in the Eastern Mediterranean [26]. For autosomal diversity or diversity on the X chromosome, no higher values in the area of domestication were confirmed. Certainly, the recent breeds kept in these areas no longer represent the genotype of the original thin-tailed sheep, which were one of the first to spread further around the world [21].
Of the specific regions monitored in sheep mtDNA, researchers most often focus on the D-loop region and the cytochrome-b-coding region [27]. More than 900 haplotypes have been found for cytochrome b [28]. It is currently assumed that there are up to six different haplogroups into which they can be divided, called A, B, C, D, E, and X. However, haplogroup X has so far only been described by a single study [29], and no subsequent studies have confirmed its existence. Based on genetic material from archaeological finds, it is assumed that there were originally more haplogroups and that some of them became extinct [30]. A and B are the most common groups in sheep from Europe (B) and Asia (A) and were also the first to be identified [10,31]. C is more genetically variable than the previous two groups but has nothing in common with any wild-type sheep. To a small extent, this type occurs in native Portuguese sheep, as well as in the Caucasus, the Middle East, and Asia. The D and E haplogroups are two of the rarest and were found in the North Caucasus region [26]. D also appears to be the haplogroup closest to the common ancestor of sheep and mouflon [32]. The last haplogroup was found at the Anatolian mouflon (Ovis gmelini anatolica Valenciennes, 1856), and it is very close to groups E and C [29]. The period of formation of these haplogroups is assumed to be sometime in the period of 5–35 thousand years ago, which is more than 150 thousand years later than the expected separation of the Cypriot mouflon (Ovis gmelini ophion Blyth, 1841) [13,32].
In addition to the study of population dispersion, the mitochondrial genome is also used for phylogenetic analyses of the genus Ovis. For example, according to a study that analyzed mitochondrial cytochrome b sequences [13], argali (Ovis ammon (Linnaeus, 1758)) was the first of the genus Ovis to diverge in Europe. The Meadows collective came to a somewhat different dating based on a study of complete mitogenomes in domestic and wild sheep [33]. They determined a calibration point based on the sequence of cytochrome b of an already extinct relative of the genus Myotragus, which separated 5.35 mya (million years ago) [33]. Based on this, they estimated the cleavage of the two major mitochondrial haplogroups, A and B, from the unfrequented C and E to 0.92 mya, and the separation of C from E to 0.26 mya. Sanna’s team reached a different estimate of the diversification of mitochondrial haplogroups on samples of whole mtDNA sequences (see Figure 1) [32]. The first separation of the two major branches of haplogroups (C, E and A, B, D) happened 0.3 mya, according to Sanna’s team. Haplogroup D (0.24 mya) was the first to be separated, groups A and B (0.17 mya) were further distinguished, and C and E (0.12 mya) were the last [32]. Table 1 provides a comparison of the estimates of five different studies of the three main divergence points preceding the formation of individual mitochondrial haplogroups.
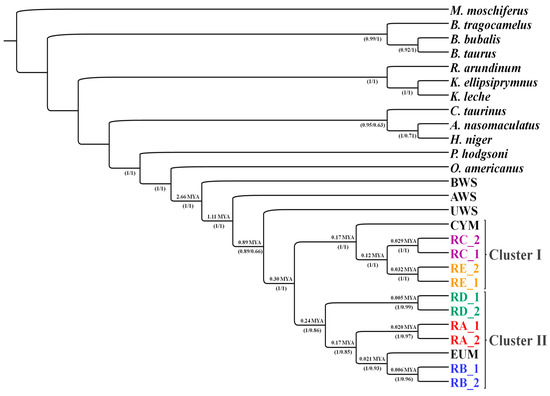
Figure 1.
Root phylogram obtained by Bayesian inference from 28 haplogroups. The labels below the nodes indicate bootstrap values for maximum similarity. Above the nodes are the molecular datings in millions of years. Cluster 1 contains three groups, one with haplogroup E (RE), the other with haplogroup C (RC), and the third with all Cypriot mouflons (CYM) and some Anatolian mouflons. The second cluster has a total of four groups. Three haplogroups of domestic sheep were divided according to haplotypes D, A, B (RD, RA, RB), and European mouflon (EUM). Some Anatolian mouflons are also included in haplogroup A. BWS, AWS, UWS means Ovis canadensis (bighorn sheep), Ovis vignei (urial), and Ovis ammon (argali), respectively. Taken from [32].

Table 1.
Comparison of approximated divergence times in million years ago (mya) of Argali, Urial and main mitochondrial haplogroup branches (A, B, D) between different studies.
In any case, it is not possible to assume a connection between the divergence of any of the five haplogroups and the post-domestication expansion, which according to archaeological findings, dates as far back as the eighth millennium BP [3,36]. Based on molecular genetic data, this may have happened two millennia earlier [14,35]. For lineage B, a primary haplotype may have already been identified in 2019, when a lineage B haplotype was found in the Sardinian mouflon and which was estimated to split about 110,000 years ago, about 30,000 years earlier than the expected division of the European mouflon from sheep lineage B [37].
The sheep probably got to North America by migrating from Asia across the Bering Strait. Bighorn sheep (Ovis canadensis (Shaw, 1804)) and Alaskan sheep (Ovis dalli (Nelson, 1884)) are monophyletic from the Siberian snow sheep (Ovis nivicola (Eschscholtz, 1829)) from which they separated about 1.6 million years ago [13]. Similar conclusions were reached in later studies [32,38].
2.1. Europe
The high diversity of sheep mitochondrial lineages could be due to domestication from several developmentally related ancestors [32]. For this reason, the complete dominance of haplogroup B mtDNA in Europe (Figure 2) indicates the expansion of the European population from only a few individuals. Such a low diversity was already present in herds in the Black Sea area before the Neolithic expansion [39]. With the Neolithic expansion, progress through Europe accelerated. Sheep reached central Anatolia about 10,000 years ago [1]. From there, two main dispersal routes led to Europe, via the Mediterranean Sea and through the Danubian valley. These two European pathways were also confirmed by research of small ruminant lentiviruses (SRLVs) [40]. The Mediterranean route led from Cyprus through the Balkan Peninsula and the Apennine Peninsula to Corsica and Sardinia from which northern Italy and southern France were further inhabited. Domesticated sheep reached the Iberian Peninsula around 7500 BP. The Danube road led through river valleys to Central Europe [1]. Sheep entered the Alps, either way, more than 5000 years ago [41]. However, there was probably another dispersal route to Europe, which led through Caucasus, Russia to northern Europe [11]. Lineage B was probably the first to reach Finland, followed by lineage A in the early Middle Ages [42]. Lineage A has spread across Europe through wool-refining efforts, but the origin of lineages C and D in Central Europe remains unclear. They could have come to Europe with a prehistoric man or much later, for example, during the Ottoman expansion [43]. This is consistent with the current findings of these two lineages, mainly in the Balkans and the Iberian Peninsula (Figure 3), which has been under Arab rule for almost seven centuries.
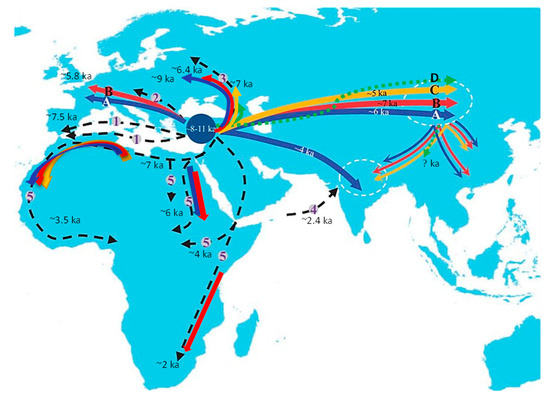
Figure 2.
The main dispersal routes of sheep from the place of domestication over Eurasia and Africa estimated in thousands of years BP. A, B, C, D = routes of major mitochondrial lines [34,44]; 1 = Mediterranean route [1,9]; 2 = Danubian route [1,9]; 3 = route to northern Europe [45]; 4 = routes of ancient sea transport to the Indian subcontinent [46]; 5 = African routes [47,48,49,50]. Taken and modified from [34].
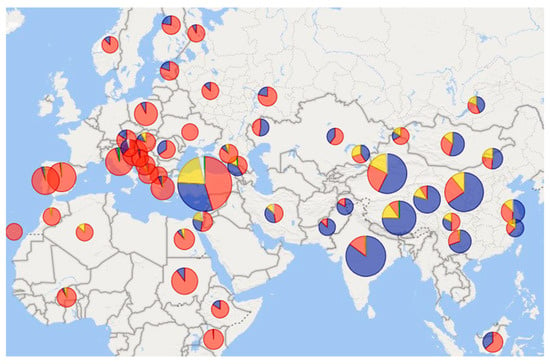
Figure 3.
Types and frequencies of mitochondrial haplogroups in different regions of the Eastern Hemisphere. Color resolution of haplogroups: A, B, C, D, E. The data used to create this map diagram with Power BI are a compilation of data from studies: [12,17,18,19,20,25,26,29,34,42,43,44,45,46,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76]. On a scale of 21–866 samples per pie chart.
2.2. Asia
The colonization of Asia was a little more complicated (Figure 2). The study evaluated the optimal model based on the ABC analysis of mitochondrial lineages, which most likely occurred during the colonization of Asia, and revealed the next three steps [34]:
- Lineage A spread first to the Mongolian Plateau and the Indian subcontinent. Later, it expanded from the Mongolian Plateau to northern and southwestern China. According to [77], lineage A was the most abundant lineage in ancient Bronze Age China (95.5%). Its abundance increased from west to east.
- Lineage B headed first on the Mongolian Plateau and colonized northern and southwestern China and India from there.
- Lineage C also first colonized the Mongolian Plateau. From there it headed to northern China and then to the Indian subcontinent.
The fourth lineage, D, was also found in Central Asia, specifically in the south of the Tibetan Plateau in one of the local breeds, Linzhou [44]. However, currently there are no relevant estimates of the time or route of its arrival at this location.
The mainland route to Asia, however, may not have been the only one. Another possibility of importing the maternal lineages A and B to India seems to be the sea route from the ancient port of Lóthal at the mouth of the Indus river [46]. At the time of Harappan culture (~2.4 thousand BP), this place had trade links with Africa and West Asia. The Mongolian Plateau was identified as the area with the highest variability [34]. Even several cases of heteroplasmy were found there, which must have existed in this area for several millennia [51]. The Mongolian Plateau acted as a migratory hub from which the lineages spread from the Middle East to Asia [34,52]. Specifically for lineages A and B, high nucleotide diversity is found in India [19,34] and for lineage C in northern China [34]. According to some authors, this diversity is so significant that it cannot come from the same domesticated animals that gave rise to these lineages in the more eastern areas. Therefore, independent domestication events could also have occurred on the Indian subcontinent [46] or in China [53,78].
So far, no study has confirmed that the Indonesian region has any original breed of sheep. The current breeds are, therefore, mainly descendants of European breeds imported by the Dutch in the second half of the 19th century [54] or fat-tailed sheep brought by Arab traders in the early 18th century [79]. However, these imported breeds were often crossed with local thin-tailed sheep of unknown origin, which were imported even earlier, probably by traders from Asia. [54]. This is probably the source of haplogroup A in Indonesia.
2.3. Africa
Evidence of the presence of sheep in Africa dates back to a much earlier time than in Southeast Asia [47]. Sheep probably penetrated North Africa through two routes about 7000 years ago. The first is the same colonization dispersion that crossed the Mediterranean Basin, the second led across Sinai, then down to and over the Red Sea [1]. There were several scattering routes on the African continent itself, south to the Middle Nile Valley, west to central Sahara, and north to Libya. Another possibility remains the spread of sheep from the Mediterranean along the northern shores of Africa. The last route discussed in Africa is the direct trade link between East Africa and the Arabian Peninsula [47]. As in Europe, mitochondrial haplogroup B is dominant in Africa (Figure 3), as confirmed in different locations—South Africa [80], Sudan [48], Kenya [49], West Africa, and the Canary Islands [50].
2.4. America and Australia
The settlement of the other two continents of America and Australia is already a matter of modern history. The first sheep brought to Central America by the Spaniards were either hair type (West African furry sheep) or coarse wool type (Churro breed from Iberia), which were later crossed with merino and gave rise to the Creole type of sheep [9]. However, it is highly probable that other breeds from the area of the Iberian Peninsula, such as Manchega, Latxa, Castellana, or Rasa Aragonesa, also contributed to the creation of Creole sheep [81].
The first hairy sheep were brought to America from the Canary Islands by Columbus and the first colonists and later along with slaves from other parts of West Africa [82]. West African sheep arrived in America in the early seventeenth century, and their contribution to the gene pool of contemporary hairy American sheep is the most significant [83]. A strong European influence was revealed in the gene pool of Creole fur sheep, which is, however, most likely caused by a later cross with merino sheep [84]. Whole-genome structural analysis of Spangler et al. showed the main influence of European breeds, especially Creole wool breeds [83]. Based on the mitochondrial maternal lineage, the origin cannot be specified because in the West African, European, and Canary sheep, maternal lineage B, which is also present in America, predominates [50]. However, another mitochondrial analysis found in Mexican Creole sheep several mitochondrial haplotypes common to both Creole sheep and two strains of Spanish sheep—Churro (Churra, Laxta, Churra Galega Mirandesa, Braganana) and Entrefino (Aragonesa, Manchega, Castellana, Castellana Stela) [81]. H2 haplotype was identified as the main ancestral mitochondrial haplotype in Mexican Creole sheep and haplotypes H21 and H32, present in both Cuba and Mexico as the possible original ones derived from hairy sheep [85].
However, it is already very difficult to study the evolution of “native” Creole sheep through genetics. The main reason is a strong disruption of the original gene pool by importing and crossing commercial breeds in the 19th and 20th centuries to Mexico [85,86,87] and other Central and South American countries [88,89].
The first sheep were brought to Australia from India, South Africa (thick-tailed), and Spain (merino) after 1788 and from the British Isles (Saxony Merino, Southdown, Romney) after 1840 [9]. It was, therefore, possible to assume the existence of the same lineages and most of the haplotypes (mt and Y) as in the populations from which Australian breeds originated. This expectation was confirmed by a study carried out on 18 breeds kept in Australia, which revealed 55% abundance of lineage B and 45% abundance of lineage A [69].
3. Haplogroups and Haplotypes of Male Y Chromosomes
The mutation rate of the male-specific region of the Y chromosome (MSY) is about fifty times lower than that of mtDNA, i.e., about 0.93 × 10−10 mutations per generation per site [35]. However, as with mitochondrial haplotypes, their use to study the phylogeny of a species is complicated by the fact that all members of the genus Ovis can interbreed and form fertile hybrids. Such insertion of a distant paternal or maternal lineage into a gene pool of another species often occurs in areas of overlap of distribution areas [13,18,29,64,90,91]. Estimates of urial and argali cleavage time have been performed in sheep based on male haplotypes so far only by [35]. In this case, the urial haplotype grouped with Asian mouflon haplotypes, in contrast to the mitochondrial genome where it formed a separate branch.
A key finding for the use of male haplotypes to study the population expansion of sheep populations was the discovery of eight SNP sites (oY1–oY8) in the sex-determining gene (SRY) on MSY [92]. And only one of them (oY1) showed variability even within the species and not only between them [93]. Subsequently, the microsatellite marker SRYM18 was discovered and thanks to it, the first 18 Y chromosomal haplotypes H1–H18 were defined [93].
In general, not as many studies have been performed in the field of research on male sheep haplotypes as in the mitochondrial genome (Figure 4). The first large worldwide study revealed, with the help of two markers (SRY SNP oY1 and microsatellite SRYM18) in domestic sheep, seven male haplotypes (H4–H10) that form two haplogroups [92]. Further research has taken over this methodological approach, including the nomenclature of the identified haplotypes. Follow-up studies revealed another H12 haplotype-specific for the Turkish Sakiz breed [93,94]. Other new haplotypes were subsequently discovered in Croatia—H18 [58] and northern China—H19, Ha, Hb [95,96]. Paternal genetic diversity of sheep has also been studied in Estonia and Finland on samples from the Bronze and Iron Ages [17]. However, only one SNP marker on the SRY gene (G-oY1) was monitored; thus, it does not provide any further information on the variability of the male genome in Europe [17].
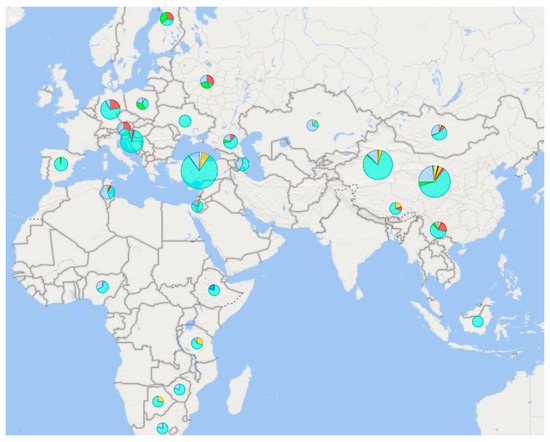
Figure 4.
Species and frequencies of male haplotypes based on microsatellite markers in different areas of the Eastern Hemisphere. Color resolution of haplotypes: H4 H5 H6 H7 H8 H9 H10 H12 H18 H19 Ha Hb. The data are a compilation of the results of five studies [58,92,93,94,95,96]. On a scale of 5–386 samples per pie chart.
The last breakthrough in this area was made in 2020 when whole-genome sequences were used to create a new set of MSY 495 SNPs in sheep [35]. Based on 179 samples of rams sequenced in the whole genome, they identified 49 different Y haplotypes. Based on a selection of 79 SNPs and two others published in previous studies (oY1; oY2), the study identified 58 other different haplotypes for domestic sheep belonging to four haplogroups: y-HA, y-HB, y-HC, and y- HD [35]. A total of 614 sheep from populations all over the world performed this genotyping. However, only native breeds were selected for our review (Figure 5).
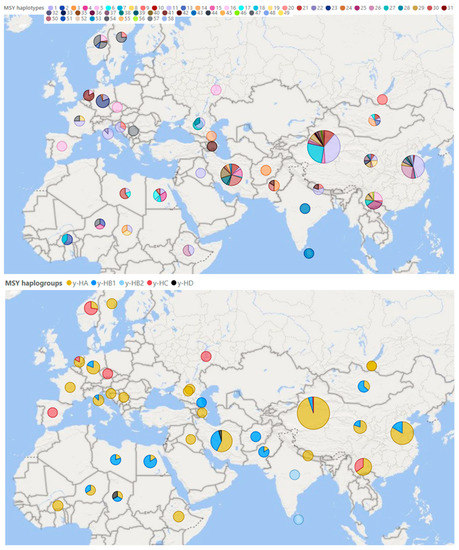
Figure 5.
Male Y haplotypes and haplogroups occurring in native sheep breeds in the Eastern Hemisphere. Illustrated with program Power BI (Microsoft, 2022) [55]. The pictured data were taken over together with the nomenclature of the included haplotypes (H1–11; H13–33; H35–58) and haplogroups (y-HA; y-HB1; y-HB2; y-HC; y-HD) [35]. On a scale of 1–135 samples per pie chart.
Based on the genome-wide study of breeds from all over the world, a certain weak relationship was found between the degree of genetic variability and the distance from the domestication center [8]. For haplotypes inherited solely by paternal or maternal lineage, this phenomenon should be more pronounced because they are not affected by the recombination process and are transmitted from generation to generation in practically unchanged form. So far, however, current research does not suggest anything like this (Figure 4 and Figure 5).
Overall, the highest variability of male haplotypes is in sheep populations from areas close to the original center of domestication in the Middle East [95]. However, more data will be needed to support this assumption. Current findings so far point to the greatest variability in central China; see Figure 4. In contrast, the diversity of mitochondrial lines so far supports this assumption, as all known lines have been found in Turkey and Israel (Figure 3)—in the supposed original place of domestication.
4. Historical Background
It can be concluded that the highest diversity of the genome can be found as close as possible to the place of origin, as has been shown, for example, in humans [97]. For sheep, research in this area is a bit more complicated. Due to the controlled reproduction and trade of animals over long distances practically from the beginning of breeding, it is possible to infer a faster and more rapid spread of genetic material than was the case in humans. Initially, sheep farming focused mainly on meat, and specialization in secondary production elements, such as wool and milk, probably did not occur until many millennia later in Asia (7–6 thousand years BP) and millennia later in Europe [14,35,98]. Specialization in wool production probably originated in Southwest Asia and only then spread to Europe, which is confirmed by the study of retroviruses [14] and by the analysis of DNA of European sheep from the Bronze Age [99]. The introduction of a new breed into Central Europe in the late Stone Age is also indicated by archaeological findings. A comparison with older findings confirmed the increase in the body frame of sheep breeds bred in the area of Bohemia and Moravia [100] and since the beginning of the Bronze age as well in Hungary [101]. Another example is the spread of merino sheep from the Iberian Peninsula since the second half of the 15th century throughout Europe [102]. Most primitive breeds today have withstood the second wave of migration to Europe in a semi-wild or wild state in isolated areas without predators or outside areas economically prone to introgression [14].
The first expansion of sheep went along with man mainly overland to Europe, Africa, and then deeper into Asia during the Mesolithic and Neolithic periods. Sheep reached other continents (America and Australia) much later with the first European colonists. The use of haplotypes to study the distribution of domestic sheep and its breeds in modern history is almost impossible by modern modes of transport. Particularly, commercially used breeds create something like a “global population” in which it is not possible to exclude the genetic proximity of individuals on different continents. Mitochondrial and Y haplotypes do not generate sufficiently genetically unique markers to study genetic diversity at this level. However, their potential for studying the phylogeny of the species, and especially its population dispersion, remains untapped.
5. Conclusions
The aim of this study was to capture the process of monitoring the dispersion and development of domestic sheep populations in different parts of the world through the study of male and female non-recombinant sections of DNA. The current review supports the existence of one domestication center in the Middle East. Nevertheless, crossbreeding with wild sheep species has probably often happened and occasionally continues to occur even now. This could be the reason why central China shows such high variability in male haplotypes. However, it can also be caused by the extinction of these variants in the Middle East, with Central Asia being a kind of reservoir of variability originating from the ancient sheep brought in from the fertile crescent.
The main challenge for the future is to involve more countries and regions while increasing the number of animals used for sequencing. Only based on a larger amount of these data combined with the genetic material from excavations, it will be possible to identify other domestication centers or refute their existence. It would also be necessary to unify the methodology and nomenclature of haplotypes for better comparability of the results of different research. But perhaps we may never know the truth because a vast amount of information is already lost forever.
Author Contributions
Conceptualization, investigation, visualization, writing—original draft preparation, K.M.; writing—review and editing, A.M.; supervision, L.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Agriculture, Czech Republic No. QK1910156 and by the Czech University of Life Sciences, Czech Republic, SGS grant No. SV21-5-21360.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zeder, M.A. Out of the Fertile Crescent: The Dispersal of Domestic Livestock through Europe and Africa. In Human Dispersal and Species Movement; Boivin, N., Petraglia, M., Crassard, R., Eds.; Cambridge University Press: Cambridge, UK, 2017; pp. 261–303. [Google Scholar]
- Zeder, M.A. Animal Domestication in the Zagros: A Review of Past and Current Research. Paléorient 1999, 25, 11–25. [Google Scholar] [CrossRef]
- Baird, D.; Fairbairn, A.; Jenkins, E.; Martin, L.; Middleton, C.; Pearson, J.; Asouti, E.; Edwards, Y.; Kabukcu, C.; Mustafaoğlu, G.; et al. Agricultural Origins on the Anatolian Plateau. Proc. Natl. Acad. Sci. USA 2018, 115, E3077–E3086. [Google Scholar] [CrossRef] [PubMed]
- Luigi-Sierra, M.G.; Mármol-Sánchez, E.; Amills, M. Comparing the Diversity of the Casein Genes in the Asian Mouflon and Domestic Sheep. Anim. Genet. 2020, 51, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Wright, D. Article Commentary: The Genetic Architecture of Domestication in Animals. Bioinform. Biol. Insights 2015, 9 (Suppl. S4), 11–20. [Google Scholar] [CrossRef]
- Lv, F.-H.; Agha, S.; Kantanen, J.; Colli, L.; Stucki, S.; Kijas, J.W.; Joost, S.; Li, M.-H.; Ajmone Marsan, P. Adaptations to Climate-Mediated Selective Pressures in Sheep. Mol. Biol. Evol. 2014, 31, 3324–3343. [Google Scholar] [CrossRef] [PubMed]
- Salces-Ortiz, J.; González, C.; Martínez, M.; Mayoral, T.; Calvo, J.H.; Serrano, M. Looking for Adaptive Footprints in the HSP90AA1 Ovine Gene. BMC Evol. Biol. 2015, 15, 7. [Google Scholar] [CrossRef]
- Kijas, J.W.; Lenstra, J.A.; Hayes, B.; Boitard, S.; Porto Neto, L.R.; San Cristobal, M.; Servin, B.; McCulloch, R.; Whan, V.; Gietzen, K.; et al. Genome-Wide Analysis of the World’s Sheep Breeds Reveals High Levels of Historic Mixture and Strong Recent Selection. PLoS Biol. 2012, 10, e1001258. [Google Scholar] [CrossRef]
- Ryder, M.L. Sheep. In Evolution of Domesticated Animals; Manson, I.L., Ed.; Longman Group Limited: London, UK; New York, NY, USA, 1984; pp. 63–84. [Google Scholar]
- Hiendleder, S.; Kaupe, B.; Wassmuth, R.; Janke, A. Molecular Analysis of Wild and Domestic Sheep Questions Current Nomenclature and Provides Evidence for Domestication from Two Different Subspecies. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2002, 269, 893–904. [Google Scholar] [CrossRef]
- Tapio, M.; Marzanov, N.; Ozerov, M.; Ćinkulov, M.; Gonzarenko, G.; Kiselyova, T.; Murawski, M.; Viinalass, H.; Kantanen, J. Sheep Mitochondrial DNA Variation in European, Caucasian, and Central Asian Areas. Mol. Biol. Evol. 2006, 23, 1776–1783. [Google Scholar] [CrossRef] [PubMed]
- Mariotti, M.; Valentini, A.; Marsan, P.A.; Pariset, L. Mitochondrial DNA of Seven Italian Sheep Breeds Shows Faint Signatures of Domestication and Suggests Recent Breed Formation. Mitochondrial DNA 2013, 24, 577–583. [Google Scholar] [CrossRef]
- Rezaei, H.R.; Naderi, S.; Chintauan-Marquier, I.C.; Taberlet, P.; Virk, A.T.; Naghash, H.R.; Rioux, D.; Kaboli, M.; Pompanon, F. Evolution and Taxonomy of the Wild Species of the Genus Ovis (Mammalia, Artiodactyla, Bovidae). Mol. Phylogenet. Evol. 2010, 54, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Chessa, B.; Pereira, F.; Arnaud, F.; Amorim, A.; Goyache, F.; Mainland, I.; Kao, R.R.; Pemberton, J.M.; Beraldi, D.; Stear, M.J.; et al. Revealing the History of Sheep Domestication Using Retrovirus Integrations. Science 2009, 324, 532–536. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Chen, L.; Wang, W. Genomic Insights into Ruminant Evolution: From Past to Future Prospects. Zool. Res. 2019, 40, 476–487. [Google Scholar] [CrossRef] [PubMed]
- Kijas, J.W.; Townley, D.; Dalrymple, B.P.; Heaton, M.P.; Maddox, J.F.; McGrath, A.; Wilson, P.; Ingersoll, R.G.; McCulloch, R.; McWilliam, S.; et al. A Genome Wide Survey of SNP Variation Reveals the Genetic Structure of Sheep Breeds. PLoS ONE 2009, 4, e4668. [Google Scholar] [CrossRef] [PubMed]
- Rannamäe, E.; Lõugas, L.; Niemi, M.; Kantanen, J.; Maldre, L.; Kadõrova, N.; Saarma, U. Maternal and Paternal Genetic Diversity of Ancient Sheep in Estonia from the Late Bronze Age to the Post-Medieval Period and Comparison with Other Regions in Eurasia. Anim. Genet. 2016, 47, 208–218. [Google Scholar] [CrossRef]
- Zhao, Y.-X.; Yang, J.; Lv, F.-H.; Hu, X.-J.; Xie, X.-L.; Zhang, M.; Li, W.-R.; Liu, M.-J.; Wang, Y.-T.; Li, J.-Q.; et al. Genomic Reconstruction of the History of Native Sheep Reveals the Peopling Patterns of Nomads and the Expansion of Early Pastoralism in East Asia. Mol. Biol. Evol. 2017, 34, 2380–2395. [Google Scholar] [CrossRef] [PubMed]
- Pardeshi, V.C.; Kadoo, N.Y.; Sainani, M.N.; Meadows, J.R.S.; Kijas, J.W.; Gupta, V.S. Mitochondrial Haplotypes Reveal a Strong Genetic Structure for Three Indian Sheep Breeds. Anim. Genet. 2007, 38, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Nigussie, H.; Mwacharo, J.M.; Osama, S.; Agaba, M.; Mekasha, Y.; Kebede, K.; Abegaz, S.; Pal, S.K. Genetic Diversity and Matrilineal Genetic Origin of Fat-Rumped Sheep in Ethiopia. Trop. Anim. Health Prod. 2019, 51, 1393–1404. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-H.; Zhang, M.; Lv, F.-H.; Ren, X.; Li, W.-R.; Liu, M.-J.; Nam, K.; Bruford, M.W.; Li, M.-H. Contrasting Patterns of Genomic Diversity Reveal Accelerated Genetic Drift but Reduced Directional Selection on X-Chromosome in Wild and Domestic Sheep Species. Genome Biol. Evol. 2018, 10, 1282–1297. [Google Scholar] [CrossRef]
- Chessa, S.; Giambra, I.J.; Brandt, H.; Caroli, A.M.; Gootwine, E.; Erhardt, G. Genetic Diversity within Economically Important Loci in European, Middle Eastern, and African Sheep Breeds: An Insight into Their Development. Small Rumin. Res. 2017, 155, 72–80. [Google Scholar] [CrossRef]
- Brown, W.M.; George, M.; Wilson, A.C. Rapid Evolution of Animal Mitochondrial DNA. Proc. Natl. Acad. Sci. USA 1979, 76, 1967–1971. [Google Scholar] [CrossRef] [PubMed]
- Rafia, P.; Tarang, A. Sequence Variations of Mitochondrial DNA Displacement-Loop in Iranian Indig…: Elektronické Publikace Dostupné Na ČZU. Iran. J. Appl. Anim. Sci. 2016, 6, 363–368. [Google Scholar]
- Mukhametzharova, I.; Islamov, Y.; Shauyenov, S.; Ibrayev, D.; Atavliyeva, S.; Tarlykov, P. Genetic Characterization of Kazakh Native Sheep Breeds Using Mitochondrial DNA. Online J. Biol. Sci. 2018, 18, 341–348. [Google Scholar] [CrossRef]
- Meadows, J.R.S.; Cemal, I.; Karaca, O.; Gootwine, E.; Kijas, J.W. Five Ovine Mitochondrial Lineages Identified from Sheep Breeds of the near East. Genetics 2007, 175, 1371–1379. [Google Scholar] [CrossRef]
- Yağci, S.; Baş, S.; Kiraz, S. Study of Mitochondrial DNA (MtDNA) D-Loop Region Polymorphism in Şavak Akkaraman Sheep. Turk. J. Vet. Anim. Sci. 2020, 44, 323–330. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information NCBI. Available online: https://www.ncbi.nlm.nih.gov/ipg/ADI57605.1 (accessed on 26 June 2020).
- Demirci, S.; Koban Baştanlar, E.; Dağtaş, N.D.; Pişkin, E.; Engin, A.; Özer, F.; Yüncü, E.; Doğan, Ş.A.; Togan, İ. Mitochondrial DNA Diversity of Modern, Ancient and Wild Sheep (Ovis gmelinii anatolica) from Turkey: New Insights on the Evolutionary History of Sheep. PLoS ONE 2013, 8, e81952. [Google Scholar] [CrossRef]
- Dymova, M.A.; Zadorozhny, A.V.; Mishukova, O.V.; Khrapov, E.A.; Druzhkova, A.S.; Trifonov, V.A.; Kichigin, I.G.; Tishkin, A.A.; Grushin, S.P.; Filipenko, M.L. Mitochondrial DNA Analysis of Ancient Sheep from Altai. Anim. Genet. 2017, 48, 615–618. [Google Scholar] [CrossRef]
- Wood, N.J.; Phua, S.H. Variation in the Control Region Sequence of the Sheep Mitochondrial Genome. Anim. Genet. 2009, 27, 25–33. [Google Scholar] [CrossRef]
- Sanna, D.; Barbato, M.; Hadjisterkotis, E.; Cossu, P.; Decandia, L.; Trova, S.; Pirastru, M.; Leoni, G.G.; Naitana, S.; Francalacci, P.; et al. The First Mitogenome of the Cyprus Mouflon (Ovis Gmelini Ophion): New Insights into the Phylogeny of the Genus Ovis. PLoS ONE 2015, 10, e0144257. [Google Scholar] [CrossRef]
- Meadows, J.R.S.; Hiendleder, S.; Kijas, J.W. Haplogroup Relationships between Domestic and Wild Sheep Resolved Using a Mitogenome Panel. Heredity 2011, 106, 700–706. [Google Scholar] [CrossRef]
- Lv, F.-H.; Peng, W.-F.; Yang, J.; Zhao, Y.-X.; Li, W.-R.; Liu, M.-J.; Ma, Y.-H.; Zhao, Q.-J.; Yang, G.-L.; Wang, F.; et al. Mitogenomic Meta-Analysis Identifies Two Phases of Migration in the History of Eastern Eurasian Sheep. Mol. Biol. Evol. 2015, 32, 2515–2533. [Google Scholar] [CrossRef]
- Deng, J.; Xie, X.-L.; Wang, D.-F.; Zhao, C.; Lv, F.-H.; Li, X.; Yang, J.; Yu, J.-L.; Shen, M.; Gao, L.; et al. Paternal Origins and Migratory Episodes of Domestic Sheep. Curr. Biol. 2020, 30, 4085–4095.e6. [Google Scholar] [CrossRef]
- Çakırlar, C. The Evolution of Animal Husbandry in Neolithic Central-West Anatolia: The Zooarchaeological Record from Ulucak Höyük (c. 7040–5660 Cal. BC, Izmir, Turkey). Anatol. Stud. 2012, 62, 1–33. [Google Scholar] [CrossRef]
- Mereu, P.; Pirastru, M.; Barbato, M.; Satta, V.; Hadjisterkotis, E.; Manca, L.; Naitana, S.; Leoni, G.G. Identification of an Ancestral Haplotype in the Mitochondrial Phylogeny of the Ovine Haplogroup B. PeerJ 2019, 7, e7895. [Google Scholar] [CrossRef]
- Dotsev, A.V.; Kunz, E.; Shakhin, A.V.; Petrov, S.N.; Kostyunina, O.V.; Okhlopkov, I.M.; Deniskova, T.E.; Barbato, M.; Bagirov, V.A.; Medvedev, D.G.; et al. The First Complete Mitochondrial Genomes of Snow Sheep (Ovis Nivicola) and Thinhorn Sheep (Ovis Dalli ) and Their Phylogenetic Implications for the Genus Ovis. Mitochondrial DNA Part B 2019, 4, 1332–1333. [Google Scholar] [CrossRef]
- Scheu, A.; Geörg, C.; Schulz, A.; Burger, J.; Benecke, N. The Arrival of Domesticated Animals in South-Eastern Europe as Seen from Ancient DNA. In Population Dynamics in Prehistory and Early History; Kaiser, E., Burger, J., Schier, W., Eds.; Walter de Gruyter: Berlin, Germany, 2012; pp. 45–54. [Google Scholar] [CrossRef]
- Molaee, V.; Bazzucchi, M.; De Mia, G.M.; Otarod, V.; Abdollahi, D.; Rosati, S.; Lühken, G. Phylogenetic Analysis of Small Ruminant Lentiviruses in Germany and Iran Suggests Their Expansion with Domestic Sheep. Sci. Rep. 2020, 10, 2243. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, C.; Ermini, L.; Rizzi, E.; Corti, G.; Luciani, S.; Marota, I.; De Bellis, G.; Rollo, F. Phylogenetic Position of a Copper Age Sheep (Ovis aries) Mitochondrial DNA. PLoS ONE 2012, 7, e33792. [Google Scholar] [CrossRef]
- Niemi, M.; Bläuer, A.; Iso-Touru, T.; Nyström, V.; Harjula, J.; Taavitsainen, J.-P.; Storå, J.; Lidén, K.; Kantanen, J. Mitochondrial DNA and Y-Chromosomal Diversity in Ancient Populations of Domestic Sheep (Ovis aries) in Finland: Comparison with Contemporary Sheep Breeds. Genet. Sel. Evol. 2013, 45, 2. [Google Scholar] [CrossRef] [PubMed]
- Gáspárdy, A.; Berger, B.; Zabavnik-Piano, J.; Kovács, E.; Annus, K.; Zenke, P.; Sáfár, L.; Maróti-Agóts, Á. Comparison of MtDNA Control Region among Descendant Breeds of the Extinct Zaupel Sheep Revealed Haplogroup C and D in Central Europe. Vet. Med. Sci. 2021, 7, 2330–2338. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ding, X.; Zeng, Y.; Yue, Y.; Guo, X.; Guo, T.; Chu, M.; Wang, F.; Han, J.; Feng, R.; et al. Genetic Diversity and Phylogenetic Evolution of Tibetan Sheep Based on MtDNA D-Loop Sequences. PLoS ONE 2016, 11, e0159308. [Google Scholar] [CrossRef]
- Tapio, I.; Värv, S.; Bennewitz, J.; Maleviciute, J.; Fimland, E.; Grislis, Z.; Meuwissen, T.H.E.; Miceikiene, I.; Olsaker, I.; Viinalass, H.; et al. Prioritization for Conservation of Northern European Cattle Breeds Based on Analysis of Microsatellite Data. Conserv. Biol. 2006, 20, 1768–1779. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Kumar, S., Jr.; Kolte, A.P.; Kumar, S. Extensive Variation and Sub-Structuring in Lineage A MtDNA in Indian Sheep: Genetic Evidence for Domestication of Sheep in India. PLoS ONE 2013, 8, e77858. [Google Scholar] [CrossRef] [PubMed]
- Muigai, A.W.T.; Hanotte, O. The Origin of African Sheep: Archaeological and Genetic Perspectives. Afr. Archaeol. Rev. 2013, 30, 39–50. [Google Scholar] [CrossRef]
- Gornas, N.; Weimann, C.; El Hussien, A.; Erhardt, G. Genetic Characterization of Local Sudanese Sheep Breeds Using DNA Markers. Small Rumin. Res. 2011, 95, 27–33. [Google Scholar] [CrossRef]
- Resende, A.; Gonçalves, J.; Muigai, A.W.T.; Pereira, F. Mitochondrial DNA Variation of Domestic Sheep (Ovis aries) in Kenya. Anim. Genet. 2016, 47, 377–381. [Google Scholar] [CrossRef]
- Álvarez, I.; Capote, J.; Traoré, A.; Fonseca, N.; Pérez, K.; Cuervo, M.; Fernández, I.; Goyache, F. Mitochondrial Analysis Sheds Light on the Origin of Hair Sheep. Anim. Genet. 2013, 44, 344–347. [Google Scholar] [CrossRef]
- Kim, Y.S.; Tseveen, K.; Batsukh, B.; Seong, J.; Kong, H.S. Origin-Related Study of Genetic Diversity and Heteroplasmy of Mongolian Sheep (Ovis arie) Using Mitochondrial DNA. J. Anim. Reprod. Biotechnol. 2020, 35, 198–206. [Google Scholar] [CrossRef]
- Ganbold, O.; Lee, S.-H.; Seo, D.; Paek, W.K.; Manjula, P.; Munkhbayar, M.; Lee, J.H. Genetic Diversity and the Origin of Mongolian Native Sheep. Livest. Sci. 2019, 220, 17–25. [Google Scholar] [CrossRef]
- Xiao, P.; Niu, L.L.; Zhao, Q.J.; Chen, X.Y.; Wang, L.J.; Li, L.; Zhang, H.P.; Guo, J.Z.; Xu, H.Y.; Zhong, T. New Insights into Mitogenomic Phylogeny and Copy Number in Eight Indigenous Sheep Populations Based on the ATP Synthase and Cytochrome c Oxidase Genes. Animal 2018, 12, 1341–1349. [Google Scholar] [CrossRef]
- Ibrahim, A.; Budisatria, I.G.S.; Widayanti, R.; Artama, W.T. The Genetic Profiles and Maternal Origin of Local Sheep Breeds on Java Island (Indonesia) Based on Complete Mitochondrial DNA D-Loop Sequences. Vet. World 2020, 13, 2625–2634. [Google Scholar] [CrossRef]
- Arora, R.; Yadav, H.S.; Mishra, B.P. Mitochondrial DNA Diversity in Indian Sheep. Livest. Sci. 2013, 153, 50–55. [Google Scholar] [CrossRef]
- Ćinkulov, M.; Popovski, Z.; Porcu, K.; Tanaskovska, B.; Hodžić, A.; Bytyqi, H.; Mehmeti, H.; Margeta, V.; Djedović, R.; Hoda, A.; et al. Genetic Diversity and Structure of the West Balkan Pramenka Sheep Types as Revealed by Microsatellite and Mitochondrial DNA Analysis. J. Anim. Breed. Genet. 2008, 125, 417–426. [Google Scholar] [CrossRef]
- Dudu, A.; Ghiţă, E.; Costache, M.; Georgescu, S.E. Origin and Genetic Diversity of Romanian Racka Sheep Using Mitochondrial Markers. Small Rumin. Res. 2016, 144, 276–282. [Google Scholar] [CrossRef]
- Ferencakovic, M.; Curik, I.; Pérez-Pardal, L.; Royo, L.J.; Cubric-Curik, V.; Fernández, I.; Álvarez, I.; Kostelic, A.; Sprem, N.; Krapinec, K.; et al. Mitochondrial DNA and Y-Chromosome Diversity in East Adriatic Sheep. Anim. Genet. 2013, 44, 184–192. [Google Scholar] [CrossRef]
- Ghernouti, N.; Bodinier, M.; Ranebi, D.; Maftah, A.; Petit, D.; Gaouar, S.B.S. Control Region of MtDNA Identifies Three Migration Events of Sheep Breeds in Algeria. Small Rumin. Res. 2017, 155, 66–71. [Google Scholar] [CrossRef]
- Gorkhali, N.A.; Han, J.L.; Ma, Y.H. Mitochondrial DNA Variation in Indigenous Sheep (Ovis aries) Breeds of Nepal. Trop. Agric. Res. 2015, 26, 632. [Google Scholar] [CrossRef]
- Guangxin, E.; Yong-Ju, Z.; Ri-Su, N.; Yue-Hui, M.; Jia-Hua, Z.; Li-Peng, C.; Xiao-Yu, Q.; Zhong-Quan, Z.; Ya-Wang, S.; Xin, W.; et al. Meta-Analysis Evidence of Maternal Lineages in Chinese Tibetan Sheep Using MtDNA D-Loop Panel. Mitochondrial DNA Part A 2017, 28, 579–583. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Du, L.-X.; Ma, Y.-H.; Guan, W.-J.; Li, H.-B.; Zhao, Q.-J.; Li, X.; Rao, S.-Q. A Novel Maternal Lineage Revealed in Sheep (Ovis aries). Anim. Genet. 2005, 36, 331–336. [Google Scholar] [CrossRef]
- Hamadalahmad, A.; Almeziad, M.; Javadmanesh, A. Genetic Similarity Comparison between Some Iranian and Middle Eastern Sheep Breeds Using Mitochondrial Control Region Sequencing. DYSONA Life Sci. 2020, 1, 20–24. [Google Scholar] [CrossRef]
- Hussain, T.; Babar, M.E.; Wajid, A. Extra Nuclear DNA Control Region and Cytochome b Gene Based Phylogeny Kashmir: Implications Towards Conservation. JAPS J. Anim. Plant Sci. 2016, 26, 1890–1893. [Google Scholar]
- Kandoussi, A.; Boujenane, I.; Auger, C.; Serranito, B.; Germot, A.; Piro, M.; Maftah, A.; Badaoui, B.; Petit, D. The Origin of Sheep Settlement in Western Mediterranean. Sci. Rep. 2020, 10, 10225. [Google Scholar] [CrossRef]
- Kirikci, K.; Noce, A.; Cam, M.A.; Mercan, L.; Amills, M. The Analysis of Mitochondrial Data Indicates the Existence of Population Substructure in Karayaka Sheep. Small Rumin. Res. 2018, 162, 25–29. [Google Scholar] [CrossRef]
- Koseniuk, A.; Słota, E. Mitochondrial Control Region Diversity in Polish Sheep Breeds. Arch. Anim. Breed. 2016, 59, 227–233. [Google Scholar] [CrossRef]
- Liu, J.; Lu, Z.; Yuan, C.; Wang, F.; Yang, B. Phylogeography and Phylogenetic Evolution in Tibetan Sheep Based on MT-CYB Sequences. Animals 2020, 10, 1177. [Google Scholar] [CrossRef] [PubMed]
- Meadows, J.R.S.; Li, K.; Kantanen, J.; Tapio, M.; Sipos, W.; Pardeshi, V.; Gupta, V.; Calvo, J.H.; Whan, V.; Norris, B.; et al. Mitochondrial Sequence Reveals High Levels of Gene Flow Between Breeds of Domestic Sheep from Asia and Europe. J. Hered. 2005, 96, 494–501. [Google Scholar] [CrossRef][Green Version]
- Mustafa, S.I.; Schwarzacher, T.; Heslop-Harrison, J.S. Complete Mitogenomes from Kurdistani Sheep: Abundant Centromeric Nuclear Copies Representing Diverse Ancestors. Mitochondrial DNA Part A 2018, 29, 1180–1193. [Google Scholar] [CrossRef]
- Oner, Y.; Calvo, J.H.; Elmaci, C. Investigation of the Genetic Diversity among Native Turkish Sheep Breeds Using MtDNA Polymorphisms. Trop. Anim. Health Prod. 2013, 45, 947–951. [Google Scholar] [CrossRef]
- Othman, O.E.; Pariset, L.; Balabel, E.A.; Marioti, M. Genetic Characterization of Egyptian and Italian Sheep Breeds Using Mitochondrial DNA. J. Genet. Eng. Biotechnol. 2015, 13, 79–86. [Google Scholar] [CrossRef]
- Pariset, L.; Mariotti, M.; Gargani, M.; Joost, S.; Negrini, R.; Perez, T.; Bruford, M.; Ajmone Marsan, P.; Valentini, A. Genetic Diversity of Sheep Breeds from Albania, Greece, and Italy Assessed by Mitochondrial DNA and Nuclear Polymorphisms (SNPs). Sci. World J. 2011, 11, 1641–1659. [Google Scholar] [CrossRef]
- Sharma, R.; Ahlawat, S.; Sharma, H.; Sharma, P.; Panchal, P.; Arora, R.; Tantia, M.S. Microsatellite and Mitochondrial DNA Analyses Unveil the Genetic Structure of Native Sheep Breeds from Three Major Agro-Ecological Regions of India. Sci. Rep. 2020, 10, 20422. [Google Scholar] [CrossRef]
- Sulaiman, Y.; Wu, C.; Zhao, C. Phylogeny of 19 Indigenous Sheep Populations in Northwestern China Inferred from Mitochondrial DNA Control Region. Asian J. Anim. Vet. Adv. 2011, 6, 71–79. [Google Scholar] [CrossRef]
- Yüncü, E.; Demirci, S.; Koban Baştanlar, E.; Doğan, Ş.A.; Taşdemir, U.; Togan, İ. Comparative Study of Three Simple Molecular Approaches in Search of MtDNA Haplogroup Identification of Domestic Sheep. Small Rumin. Res. 2013, 114, 64–71. [Google Scholar] [CrossRef]
- Cai, D.; Tang, Z.; Yu, H.; Han, L.; Ren, X.; Zhao, X.; Zhu, H.; Zhou, H. Early History of Chinese Domestic Sheep Indicated by Ancient DNA Analysis of Bronze Age Individuals. J. Archaeol. Sci. 2011, 38, 896–902. [Google Scholar] [CrossRef]
- Chen, S.-Y.; Duan, Z.-Y.; Sha, T.; Xiangyu, J.; Wu, S.-F.; Zhang, Y.-P. Origin, Genetic Diversity, and Population Structure of Chinese Domestic Sheep. Gene 2006, 376, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Udo, H.M.J.; Budisatria, I.G.S. Fat-Tailed Sheep in Indonesia; an Essential Resource for Smallholders. Trop. Anim. Health Prod. 2011, 43, 1411–1418. [Google Scholar] [CrossRef]
- Horsburgh, K.A.; Rhines, A. Genetic Characterization of an Archaeological Sheep Assemblage from South Africa’s Western Cape. J. Archaeol. Sci. 2010, 37, 2906–2910. [Google Scholar] [CrossRef]
- Alonso, R.A.; Ulloa-Arvizu, R.; Gayosso-Vázquez, A. Mitochondrial DNA Sequence Analysis of the Mexican Creole Sheep (Ovis aries) Reveals a Narrow Iberian Maternal Origin. Mitochondrial DNA Part A 2017, 28, 793–800. [Google Scholar] [CrossRef]
- Delgado, J.V.; Perezgrovas, R.; Camacho, M.E.; Fresno, M.; Barba, C. The Wool-Less Canary Sheep and Their Relationship with the Present Breeds in America. Anim. Genet. Resour. Inf. 2000, 28, 27–34. [Google Scholar] [CrossRef]
- Spangler, G.L.; Rosen, B.D.; Ilori, M.B.; Hanotte, O.; Kim, E.-S.; Sonstegard, T.S.; Burke, J.M.; Morgan, J.L.M.; Notter, D.R.; Van Tassell, C.P. Whole Genome Structural Analysis of Caribbean Hair Sheep Reveals Quantitative Link to West African Ancestry. PLoS ONE 2017, 12, e0179021. [Google Scholar] [CrossRef] [PubMed]
- Muigai, A.W.T.; Hirbo, J.; Sharkey, S.; Rege, J.E.O.; Blackburn, H.; Hanotte, O. Genetic diversity and relationships of hair sheep breeds of the Americas: First results. In Proceedings of the 7th World Congress on Genetics Applied to Livestock Production, Montpellier, France, 19–23 August 2002; pp. 1–4. [Google Scholar]
- Campos, E.; Cuéllar, J.; Salvador, O.; García-Trejo, E.A.; Pereira, F. The Genetic Diversity and Phylogeography of Mexican Domestic Sheep. Small Rumin. Res. 2020, 187, 106109. [Google Scholar] [CrossRef]
- Parsons, D.; Nicholson, C.F. Assessing Policy Options for Agricultural Livestock Development: A Case Study of Mexico’s Sheep Sector. Cogent Food Agric. 2017, 3, 1313360. [Google Scholar] [CrossRef]
- Macias-Cruz, U.; Álvarez-Valenzuela, F.D.; Correa-Calderón, A.; Molina-Ramírez, L.; González-Reyna, A.; Soto-Navarro, S.; Avendaño-Reyes, L. Pelibuey Ewe Productivity and Subsequent Pre-Weaning Lamb Performance Using Hair-Sheep Breeds Under a Confinement System. J. Appl. Anim. Res. 2009, 36, 255–260. [Google Scholar] [CrossRef]
- Revelo, H.A.; López-Alvarez, D.; Landi, V.; Rizzo, L.; Alvarez, L.A. Mitochondrial DNA Variations in Colombian Creole Sheep Confirm an Iberian Origin and Shed Light on the Dynamics of Introduction Events of African Genotypes. Animals 2020, 10, 1594. [Google Scholar] [CrossRef] [PubMed]
- Bravo, S.; Larama, G.; Quiñones, J.; Paz, E.; Rodero, E.; Sepúlveda, N. Genetic Diversity and Phylogenetic Relationship among Araucana Creole Sheep and Spanish Sheep Breeds. Small Rumin. Res. 2019, 172, 23–30. [Google Scholar] [CrossRef]
- Hu, X.-J.; Yang, J.; Xie, X.-L.; Lv, F.-H.; Cao, Y.-H.; Li, W.-R.; Liu, M.-J.; Wang, Y.-T.; Li, J.-Q.; Liu, Y.-G.; et al. The Genome Landscape of Tibetan Sheep Reveals Adaptive Introgression from Argali and the History of Early Human Settlements on the Qinghai–Tibetan Plateau. Mol. Biol. Evol. 2019, 36, 283–303. [Google Scholar] [CrossRef]
- Barbato, M.; Hailer, F.; Orozco-terWengel, P.; Kijas, J.; Mereu, P.; Cabras, P.; Mazza, R.; Pirastru, M.; Bruford, M.W. Genomic Signatures of Adaptive Introgression from European Mouflon into Domestic Sheep. Sci. Rep. 2017, 7, 7623. [Google Scholar] [CrossRef]
- Meadows, J.R.S.; Hanotte, O.; Drögemüller, C.; Calvo, J.; Godfrey, R.; Coltman, D.; Maddox, J.F.; Marzanov, N.; Kantanen, J.; Kijas, J.W. Globally Dispersed Y Chromosomal Haplotypes in Wild and Domestic Sheep. Anim. Genet. 2006, 37, 444–453. [Google Scholar] [CrossRef]
- Meadows, J.R.S.; Kijas, J.W. Re-Sequencing Regions of the Ovine Y Chromosome in Domestic and Wild Sheep Reveals Novel Paternal Haplotypes. Anim. Genet. 2009, 40, 119–123. [Google Scholar] [CrossRef]
- Parmaksiz, A.; Oymak, A.; Yüncü, E.; Demirci, S.; Koban Baştanlar, E.; Özkan Ünal, E.; Togan, İ.; Özer, F. Türkiye’den 12 Yerli, Karagül, Karacabey Merinosu ve Anadolu Yaban Koyununda (Ovis gmelinii anatolica) Y-Kromozom Polimorfizmleri. Kafkas Univ. Vet. Fak. Derg. 2018, 24, 821–828. [Google Scholar] [CrossRef]
- Zhang, M.; Peng, W.-F.; Yang, G.-L.; Lv, F.-H.; Liu, M.-J.; Li, W.-R.; Liu, Y.-G.; Li, J.-Q.; Wang, F.; Shen, Z.-Q.; et al. Y Chromosome Haplotype Diversity of Domestic Sheep (Ovis aries) in Northern Eurasia. Anim. Genet. 2014, 45, 903–907. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, L.; Yan, W.; Li, S.; Wang, J.; Liu, X.; Hu, J.; Luo, Y. Y Chromosomal Haplotype Characteristics of Domestic Sheep (Ovis aries) in China. Gene 2015, 565, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Z.; Absher, D.M.; Tang, H.; Southwick, A.M.; Casto, A.M.; Ramachandran, S.; Cann, H.M.; Barsh, G.S.; Feldman, M.; Cavalli-Sforza, L.L.; et al. Worldwide Human Relationships Inferred from Genome-Wide Patterns of Variation. Science 2008, 319, 1100–1104. [Google Scholar] [CrossRef] [PubMed]
- Becker, C.; Benecke, N.; Grabundžija, A.; Küchelmann, H.-C.; Pollock, S.; Schier, W.; Schoch, C.; Schrakamp, I.; Schütt, B.; Schumacher, M. The Textile Revolution. Research into the Origin and Spread of Wool Production between the Near East and Central Europe. eTopoi J. Anc. Stud. 2016, 6, 102–151. [Google Scholar]
- Sabatini, S.; Bergerbrant, S.; Brandt, L.Ø.; Margaryan, A.; Allentoft, M.E. Approaching Sheep Herds Origins and the Emergence of the Wool Economy in Continental Europe during the Bronze Age. Archaeol. Anthropol. Sci. 2019, 11, 4909–4925. [Google Scholar] [CrossRef]
- Kyselý, R. The Size of Domestic Cattle, Sheep, Goats and Pigs in the Czech Neolithic and Eneolithic Periods: Temporal Variations and Their Causes. Archaeofauna 2016, 25, 33–78. [Google Scholar]
- Bokonyi, S. The Development and History of Domestic Animals in Hungary: The Neolithic Through the Middle Ages. Am. Anthropol. 1971, 73, 640–674. [Google Scholar] [CrossRef]
- Landi, V.; Lasagna, E.; Ceccobelli, S.; Martinez, A.; Santos-Silva, F.; Vega-Pla, J.L.; Panella, F.; Allain, D.; Palhiere, I.; Murawski, M.; et al. An Historical and Biogeographical Assessment of European Merino Sheep Breeds by Microsatellite Markers. Small Rumin. Res. 2019, 177, 76–81. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).