
Genome-Wide Identification and Functional Exploration of SBP-Box Gene Family in Black Pepper (Piper nigrum L.)

Abstract
:1. Introduction
2. Methods
2.1. Data Sources
2.2. Identification and Characterization of Pepper SBP Genes
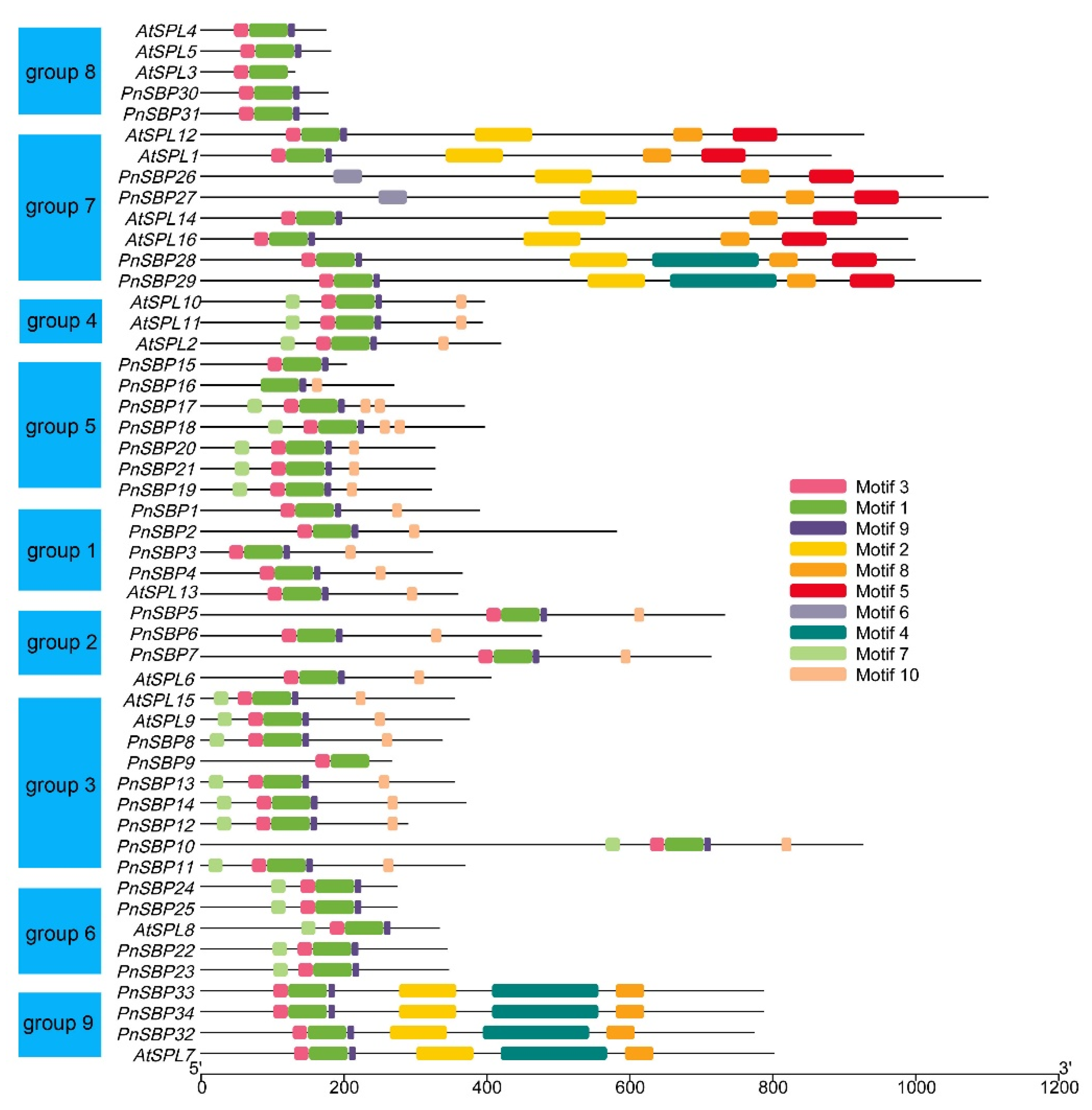
2.3. Phylogenetic Analysis and Conserved Motif Prediction
2.4. Chromosomal Localization and Detection of Gene Duplication
2.5. Estimation of Synonymous (Ks) and Nonsynonymous (Ka) Substitutions per Site and Their Ratio (Ka/Ks)
2.6. MicroRNA and Target Prediction
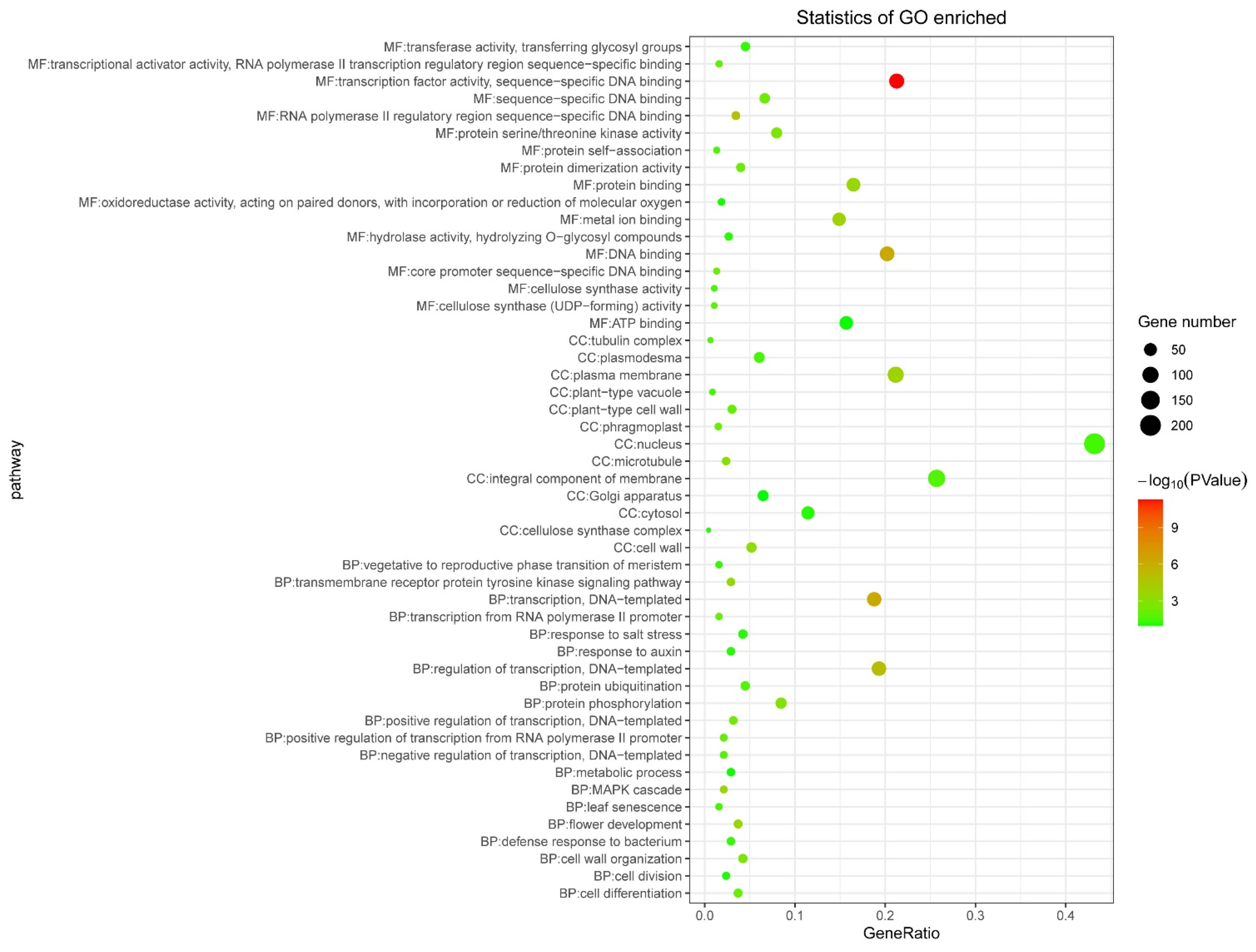
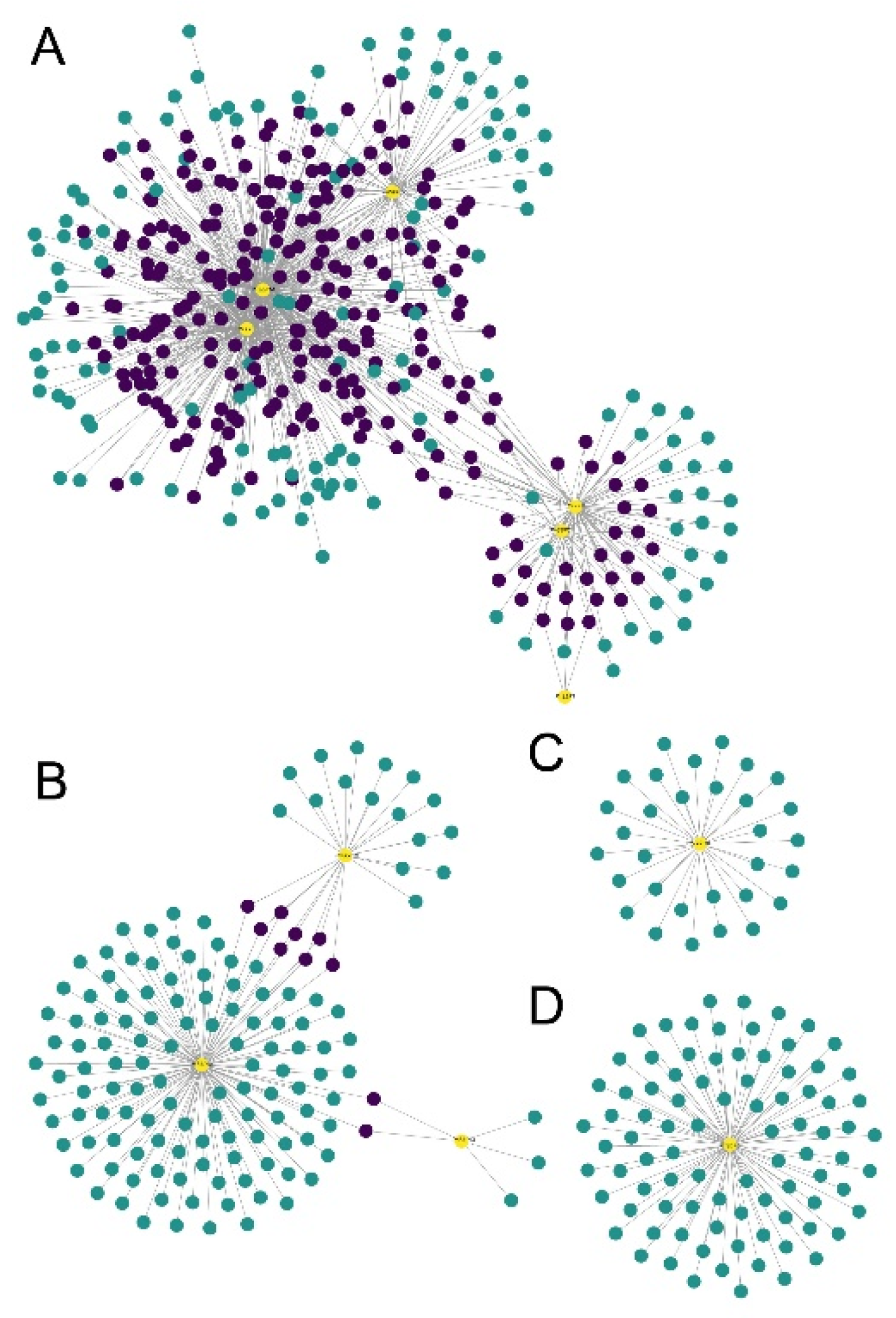
2.7. Tissue Expression, WGCNA Analysis, GO Enrichment, and Co-Expression Network Construction
3. Results
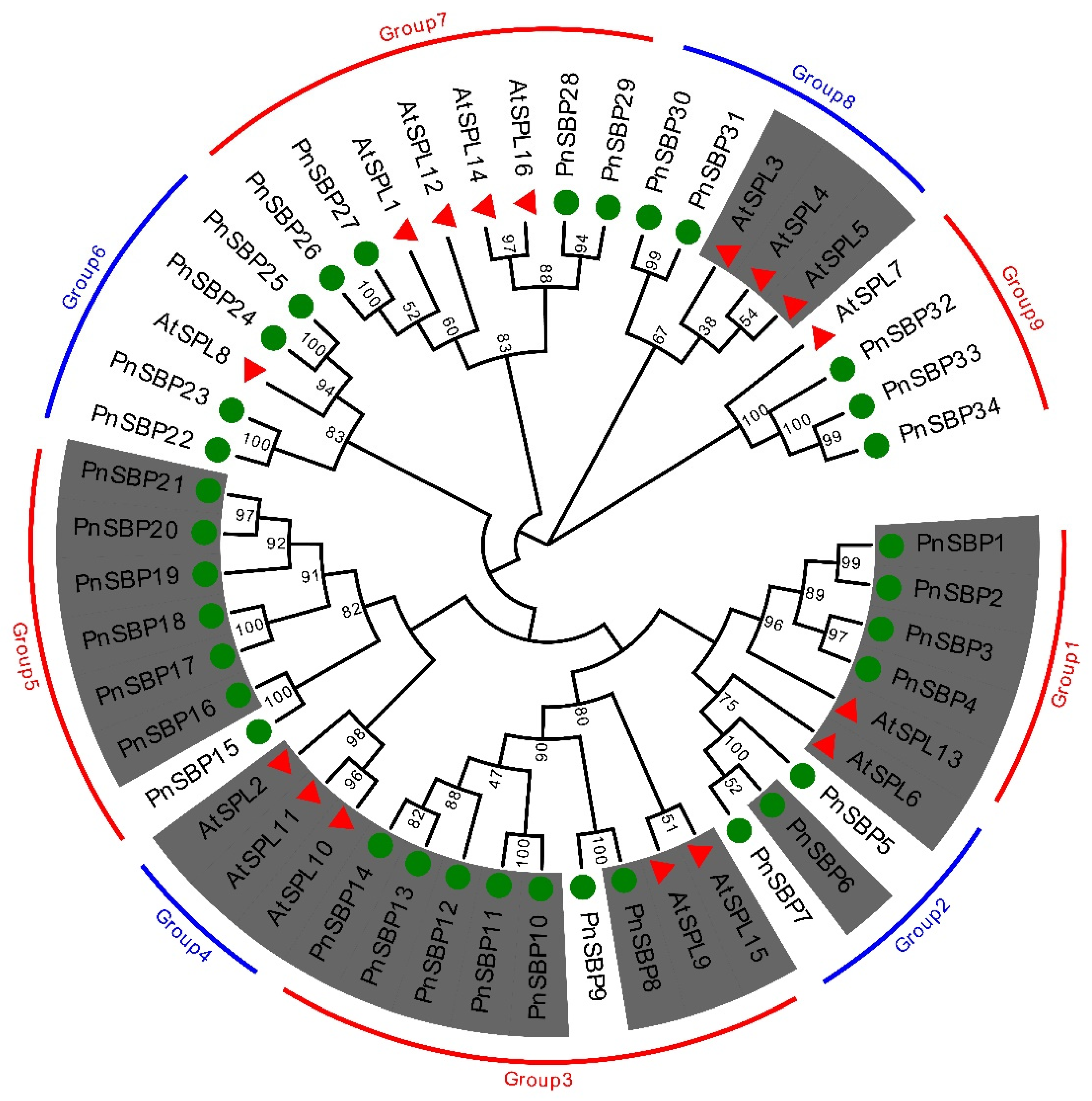
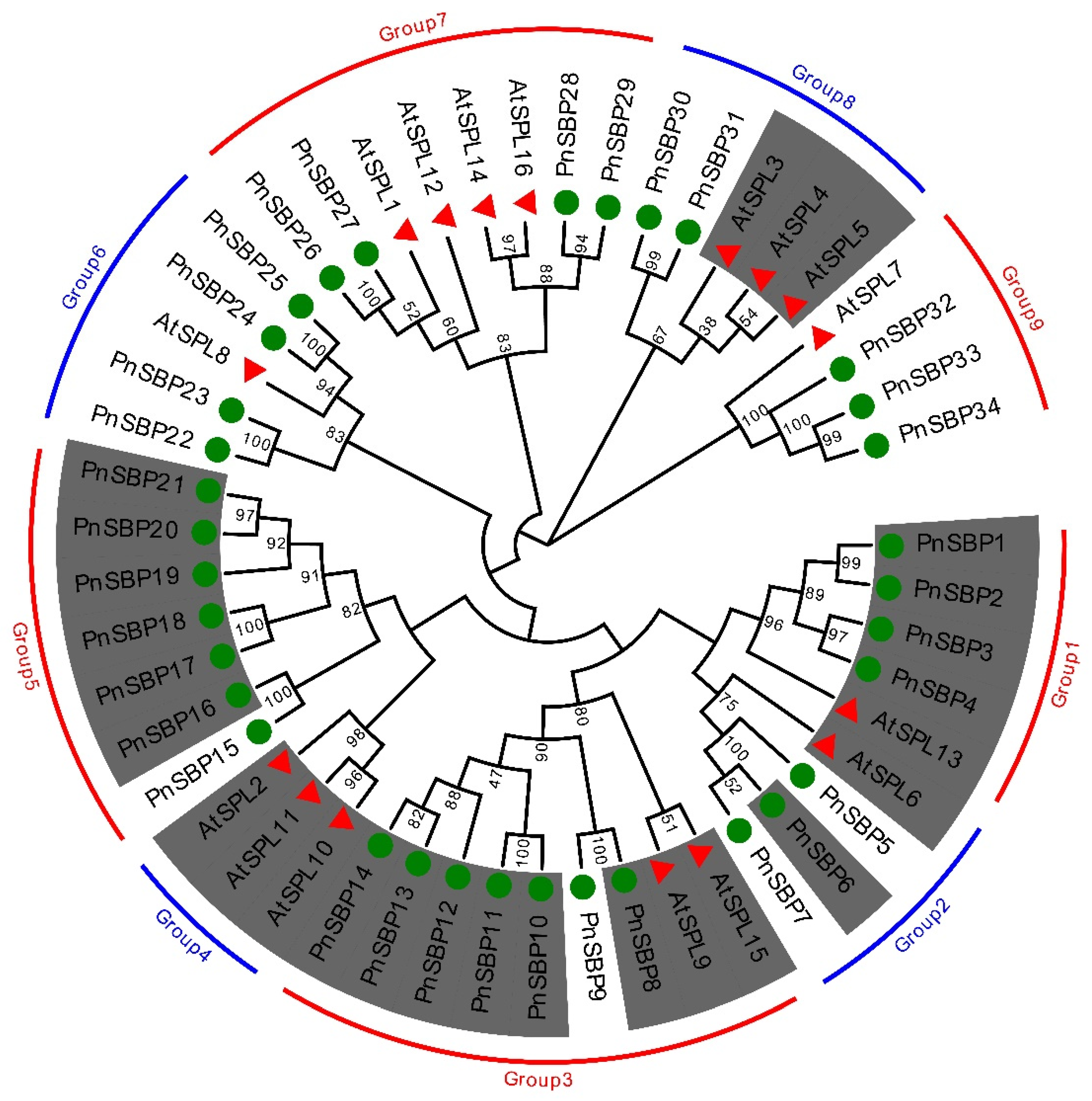
3.1. Phylogenetic Analysis
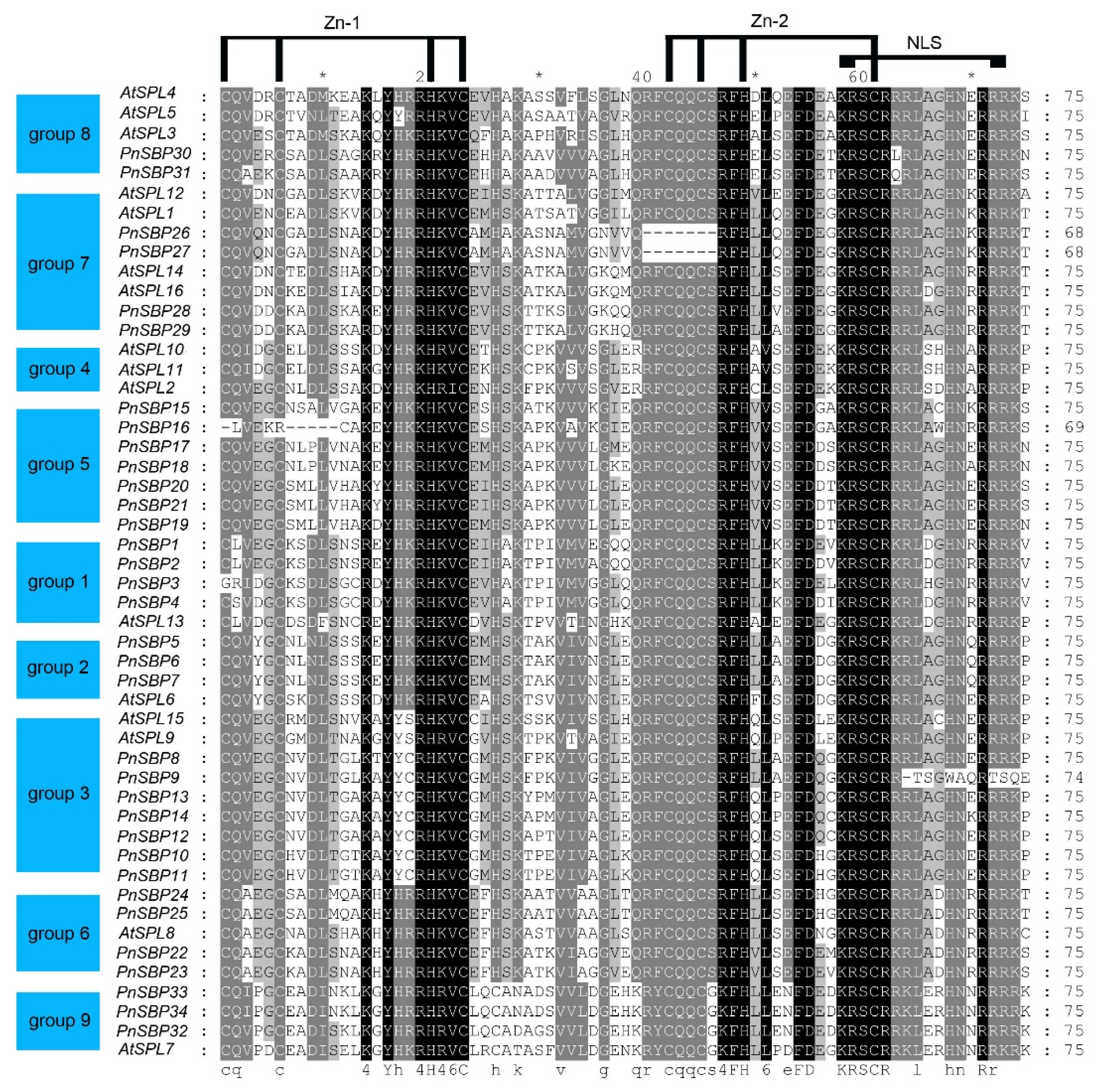
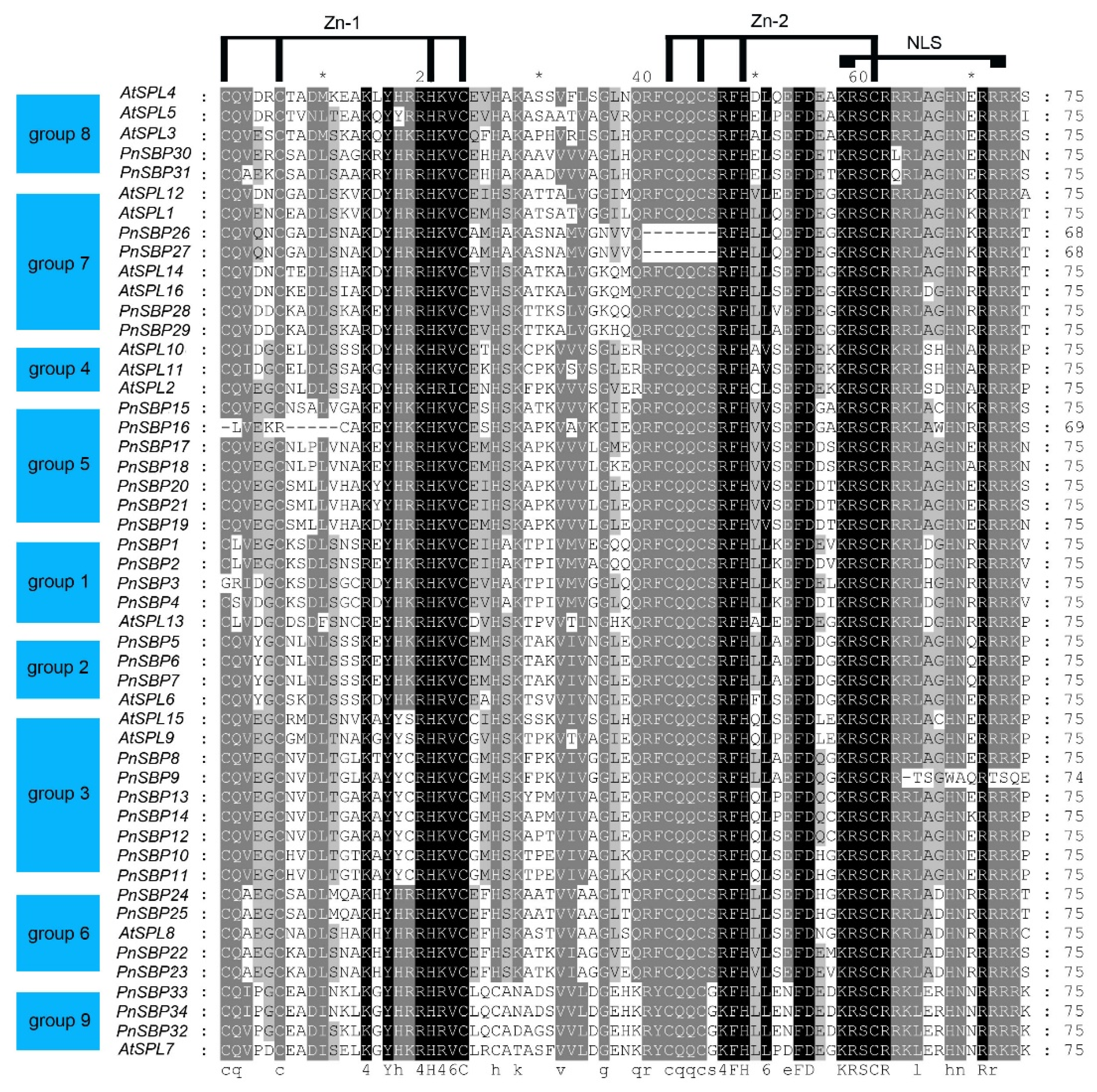
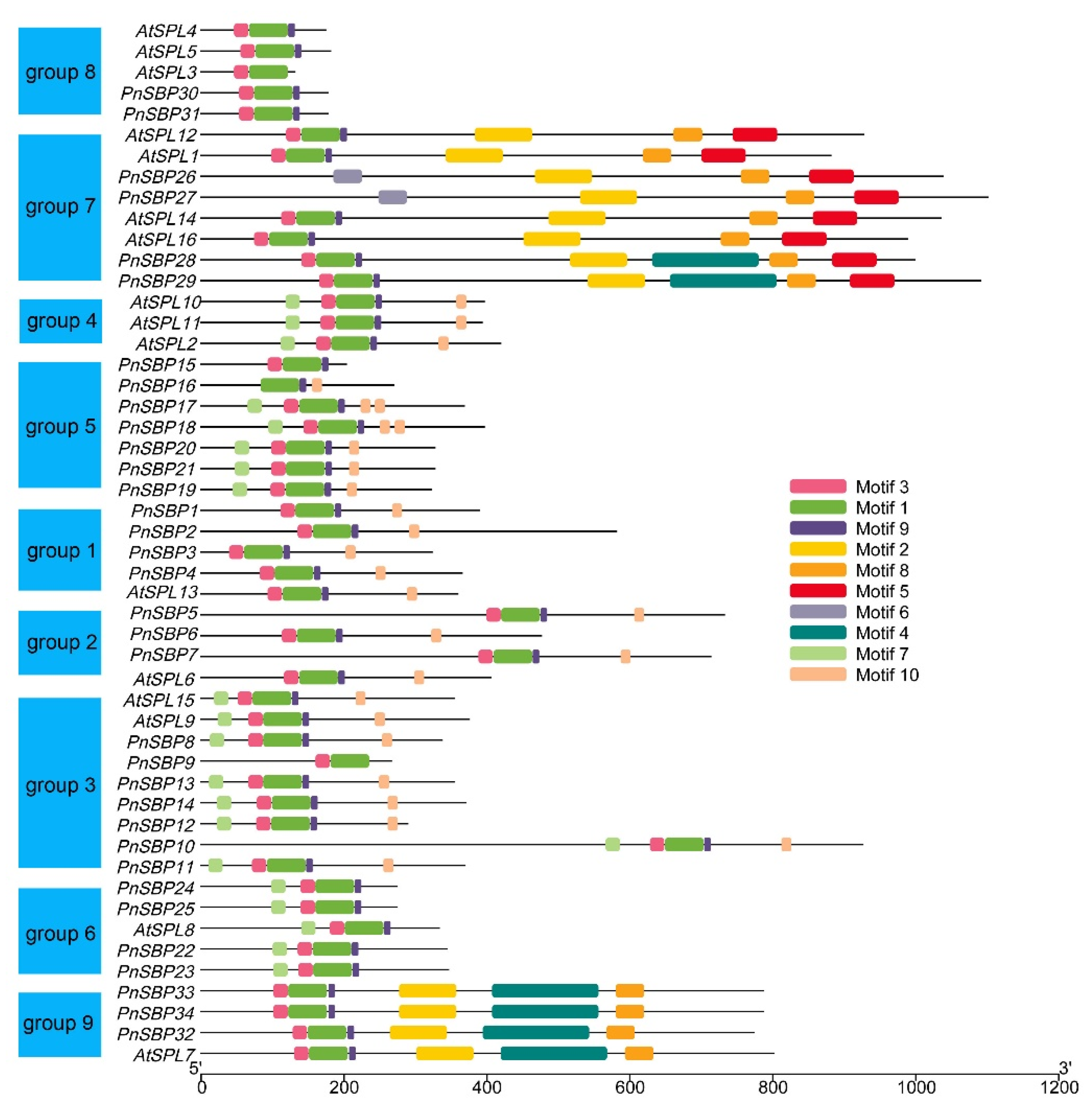
3.2. Multiple Sequence Alignment and Conserved Motif Analysis
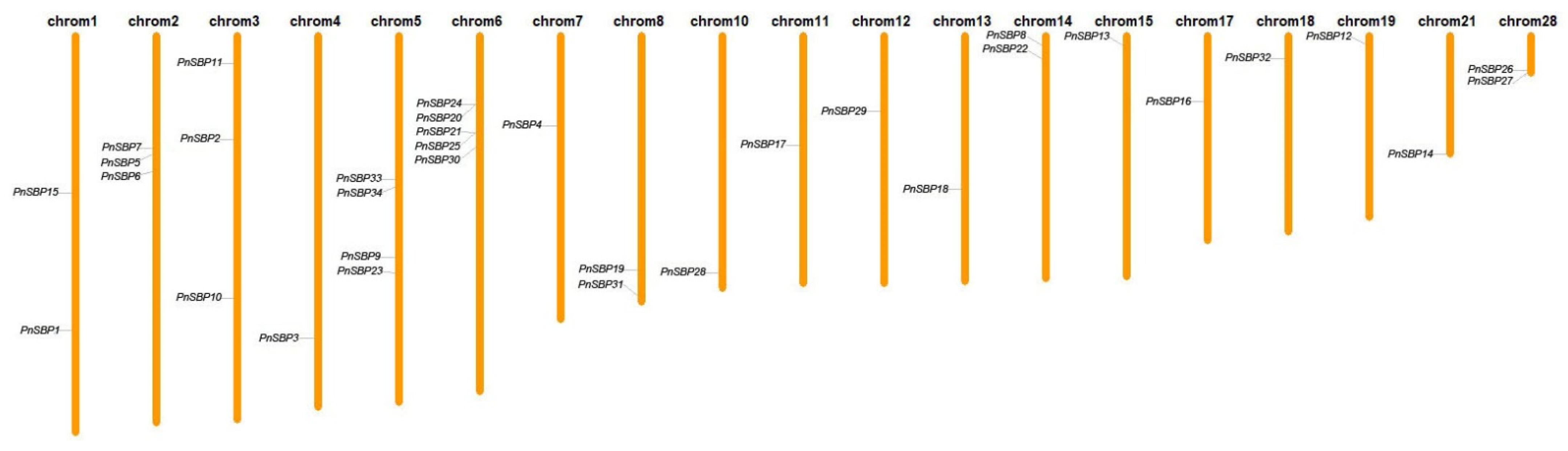
3.3. Chromosomal Localization and Gene Duplication Analysis
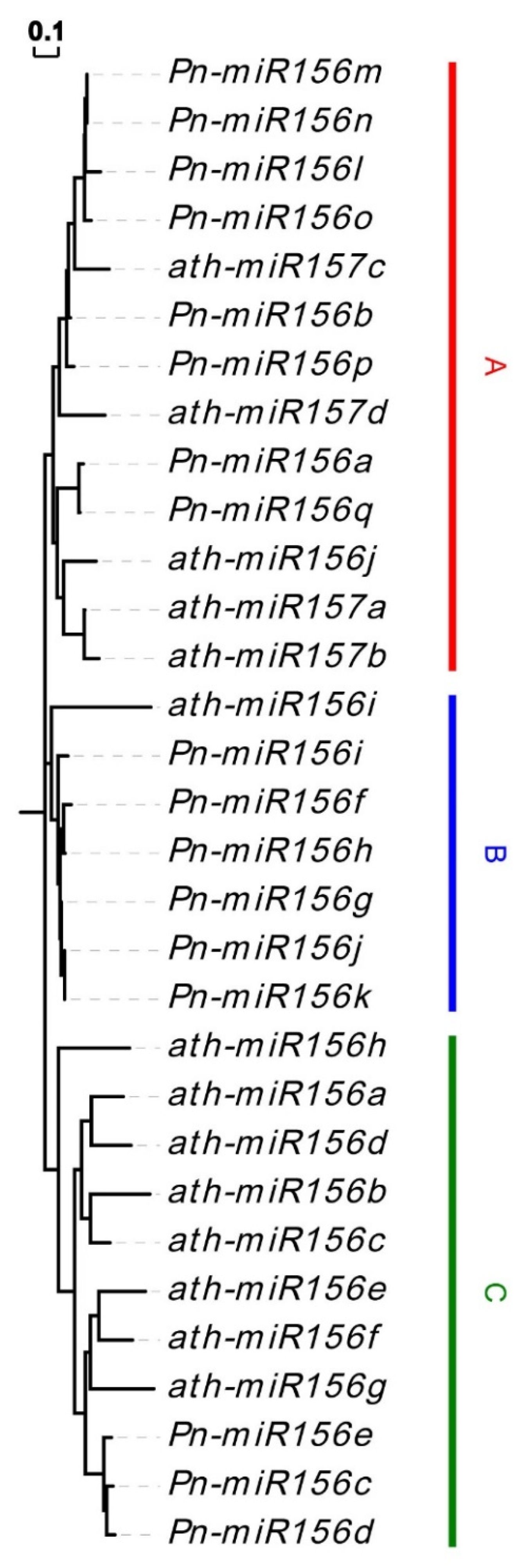
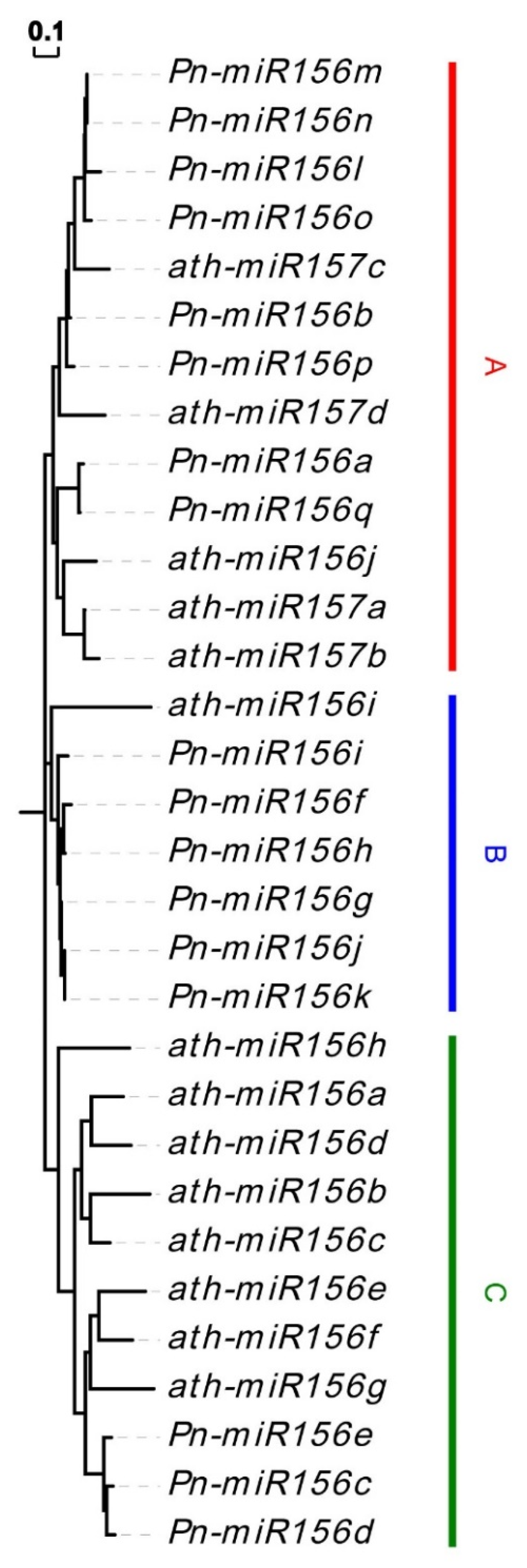
3.4. Prediction of MiR156 and Their Target Sites
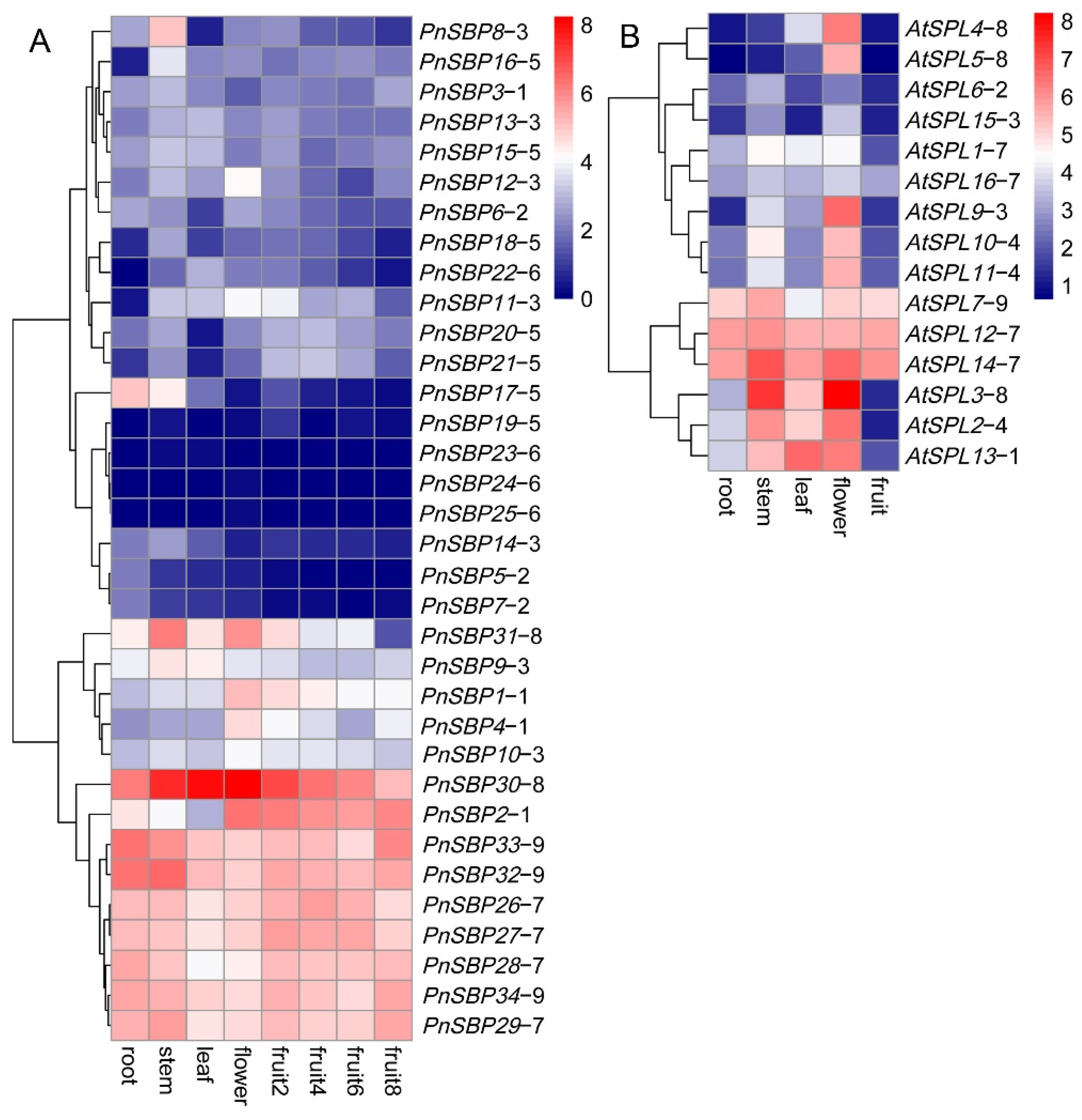
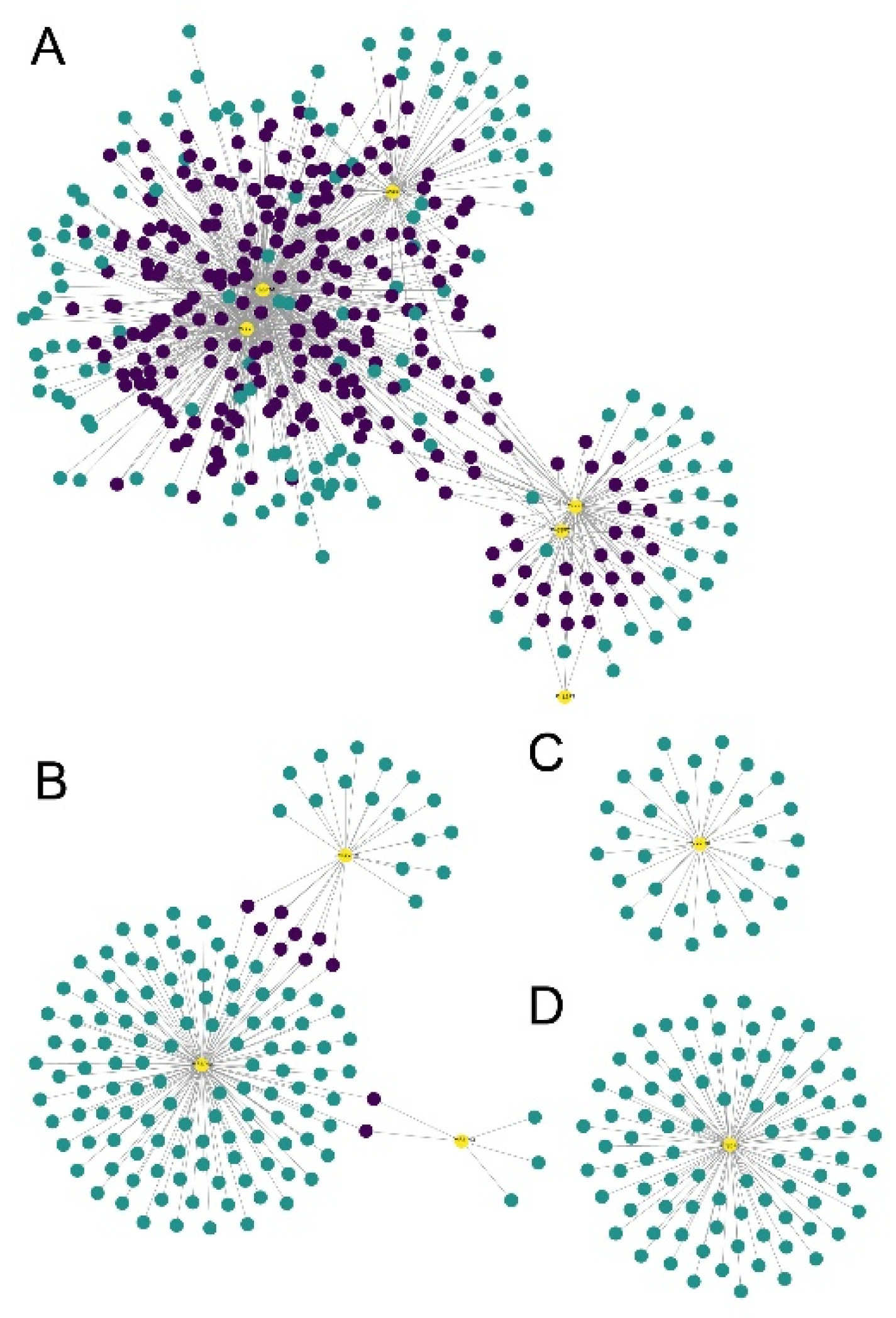
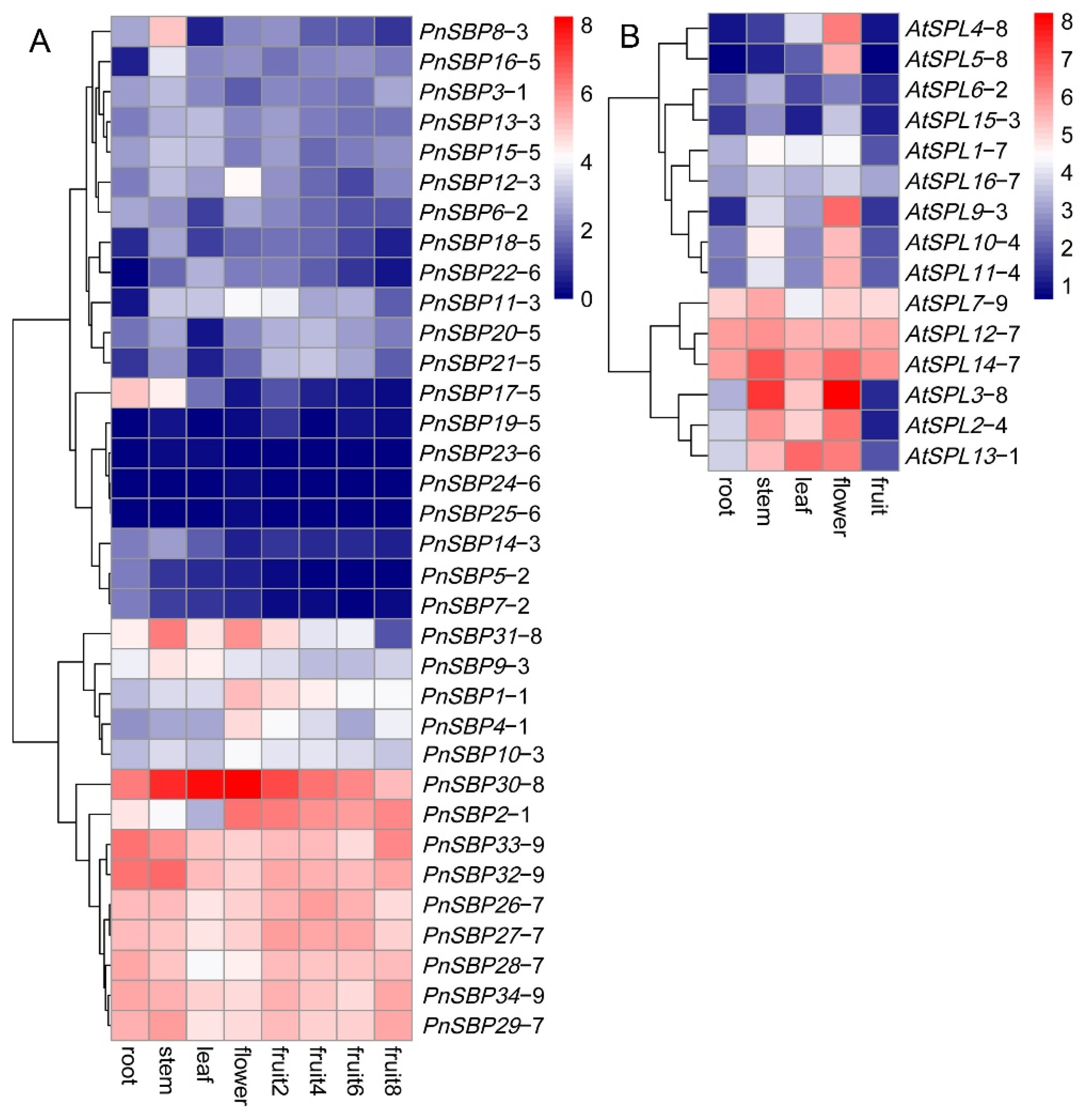
3.5. Tissue Expression and WGCNA Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Lijsebettens, M.; Grasser, K.D. Transcript elongation factors: Shaping transcriptomes after transcript initiation. Trends Plant Sci. 2014, 19, 717–726. [Google Scholar] [CrossRef]
- Yamasaki, K.; Kigawa, T.; Seki, M.; Shinozaki, K.; Yokoyama, S. DNA-binding domains of plant-specific transcription factors: Structure, function, and evolution. Trends Plant Sci. 2012, 18, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.; Saedler, H.; Huijser, P. A new family of DNA binding proteins includes putative transcriptional regulators of the Antirrhinum majus floral meristem identity gene SQUAMOSA. Mol. Gen. Genet. 1996, 250, 7–16. [Google Scholar] [PubMed]
- Riese, M.; Höhmann, S.; Saedler, H.; Münster, T.; Huijser, P. Comparative analysis of the SBP-box gene families in P. patens and seed plants. Gene 2007, 401, 28–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Wang, X.; Gu, S.; Hu, Z.; Xu, H.; Xu, C. Comparative study of SBP-box gene family in Arabidopsis and rice. Gene 2008, 407, 1–11. [Google Scholar] [CrossRef]
- Li, J.; Gao, X.; Zhang, X.; Liu, C. Dynamic expansion and functional evolutionary profiles of plant conservative gene family SBP-box in twenty two flowering plants and the origin of miR156. Biomolecules 2020, 10, 757. [Google Scholar] [CrossRef]
- Zhang, H.X.; Jin, J.H.; He, Y.M.; Lu, B.Y.; Li, D.W.; Chai, W.G.; Khan, A.; Gong, Z.H. Genome-wide identification and analysis of the SBP-box family genes under Phytophthora capsici stress in pepper (Capsicum annuum L.). Front. Plant Sci. 2016, 7, 504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Wang, H. The miR156/SPL module, a regulatory hub and versatile toolbox, gears up crops for enhanced agronomic traits. Mol. Plant 2015, 8, 677–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birkenbihl, R.P.; Jach, G.; Saedler, H.; Huijser, P. Functional dissection of the plant-specific SBP-domain: Overlap of the DNA-binding and nuclear localization domains. J. Mol. Biol. 2005, 352, 585–596. [Google Scholar] [CrossRef] [Green Version]
- Rogers, K.; Chen, X. Biogenesis, turnover, and mode of action of plant microRNAs. Plant Cell 2013, 25, 2383–2399. [Google Scholar] [CrossRef] [Green Version]
- Voinnet, O. Origin, biogenesis, and activity of plant microRNAs. Cell 2009, 136, 669–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, A.; Wu, M.F.; Yang, L.; Wu, G.; Poethig, R.S.; Wagner, D. The microRNA-regulated SBP-box transcription factor SPL3 is a direct upstream activator of leafy, FRUITFULL, and APETALA1. Dev. Cell 2009, 17, 268–278. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.H.; Lee, H.J.; Ryu, J.Y.; Park, C.M. SPL3/4/5 integrate developmental aging and photoperiodic signals into the FT-FD module in Arabidopsis flowering. Mol. Plant 2016, 9, 1647–1659. [Google Scholar] [CrossRef] [Green Version]
- Cardon, G.H.; Höhmann, S.; Nettesheim, K.; Saedler, H.; Huijser, P. Functional analysis of the Arabidopsis thaliana SBP-box gene SPL3: A novel gene involved in the floral transition. Plant J. 1997, 12, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Shikata, M.; Koyama, T.; Mitsuda, N.; Ohme-Takagi, M. Arabidopsis SBP-box genes SPL10, SPL11 and SPL2 control morphological change in association with shoot maturation in the reproductive phase. Plant Cell Physiol. 2009, 50, 2133–2145. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, S.; Grande, A.V.; Bujdoso, N.; Saedler, H.; Huijser, P. The microRNA regulated SBP-box genes SPL9 and SPL15 control shoot maturation in Arabidopsis. Plant Mol. Biol 2008, 67, 183–195. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhao, X.; Li, J.; Cai, H.; Deng, X.W.; Li, L. MicroRNA408 is critical for the HY5-SPL7 gene network that mediates the coordinated response to light and copper. Plant Cell 2014, 26, 4933–4953. [Google Scholar] [CrossRef] [Green Version]
- Salehi, B.; Zakaria, Z.A.; Gyawali, R.; Ibrahim, S.A.; Rajkovic, J.; Shinwari, Z.K.; Khan, T.; Sharifi-Rad, J.; Ozleyen, A.; Turkdonmez, E.; et al. Piper species: A comprehensive review on their phytochemistry, biological activities and applications. Molecules 2019, 24, 1364. [Google Scholar] [CrossRef] [Green Version]
- Takooree, H.; Aumeeruddy, M.Z.; Rengasamy, K.R.R.; Venugopala, K.N.; Jeewon, R.; Zengin, G.A.-O.; Mahomoodally, M.F. A systematic review on black pepper (Piper nigrum L.): From folk uses to pharmacological applications. Crit. Rev. Food Sci. Nutr. 2019, 59, S210–S243. [Google Scholar] [CrossRef] [PubMed]
- Butt, M.S.; Pasha, I.; Sultan, M.T.; Randhawa, M.A.; Saeed, F.; Ahmed, W. Black pepper and health claims: A comprehensive treatise. Crit Rev. Food Sci. Nutr. 2013, 53, 875–886. [Google Scholar] [CrossRef]
- Dosoky, N.S.; Satyal, P.; Barata, L.M.; da Silva, J.K.R.; Setzer, W.N. Volatiles of black pepper fruits (Piper nigrum L.). Molecules 2019, 24, 4244. [Google Scholar] [CrossRef] [Green Version]
- Group, T.A.P.; Chase, M.W.; Christenhusz, M.J.M.; Fay, M.F.; Byng, J.W.; Judd, W.S.; Soltis, D.E.; Mabberley, D.J.; Sennikov, A.N.; Soltis, P.S.; et al. An update of the angiosperm phylogeny group classification for the orders and families of flowering plants: Apg IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar]
- Hu, L.; Xu, Z.A.-O.; Wang, M.A.-O.; Fan, R.; Yuan, D.A.-O.; Wu, B.; Wu, H.; Qin, X.; Yan, L.; Tan, L.; et al. The chromosome-scale reference genome of black pepper provides insight into piperine biosynthesis. Nat. Commun. 2019, 10, 4702. [Google Scholar] [CrossRef]
- Lamesch, P.; Berardini, T.Z.; Li, D.; Swarbreck, D.; Wilks, C.; Sasidharan, R.; Muller, R.; Dreher, K.; Alexander, D.L.; Garcia-Hernandez, M.; et al. The Arabidopsis information resource (tair): Improved gene annotation and new tools. Nucleic Acids Res. 2011, 40, D1202–D1210. [Google Scholar] [CrossRef]
- Ghahramani, Z. An introduction to hidden markov models and bayesian networks. In Hidden markov models. World Sci. 2001, 45, 9–41. [Google Scholar]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. Blast+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The pfam protein families database in 2019. Nucleic Acids Res. 2018, 47, D427–D432. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. 20 years of the smart protein domain annotation resource. Nucleic Acids Res. 2017, 46, D493–D496. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. Expasy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wallace, I.M.; Wilm, A.; et al. Clustal W and clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Koichiro, T.; Glen, S.; Sudhir, K. MEGA11: Molecular evolutionary genetics analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, B.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W.H. Evolview v3: A webserver for visualization, annotation, and management of phylogenetic trees. Nucleic Acids Res. 2019, 47, W270–W275. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME suite: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Lee, T.H.; Tang, H.; Wang, X.; Paterson, A.H.; Paterson, A.H. PGDD: A database of gene and genome duplication in plants. Nucleic Acids Res. 2013, 41, D1152–D1158. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. Mcscanx: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Löytynoja, A. Phylogeny-aware alignment with prank and pagan. Methods Mol. Biol. 2021, 2231, 17–37. [Google Scholar]
- Wang, D.; Zhang, Y.; Zhang, Z.; Zhu, J.; Yu, J. Kaks_calculator 2.0: A toolkit incorporating γ-series methods and sliding window strategies. Genom. Proteom. Bioinf. 2010, 8, 77–80. [Google Scholar] [CrossRef] [Green Version]
- Griffiths-Jones, S.; Grocock, R.J.; van Dongen, S.; Bateman, A.; Enright, A.J. miRBase: microRNA sequences, targets and gene nomenclature. Nucleic Acids Res. 2006, 34, D140–D144. [Google Scholar] [CrossRef]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- Patanun, O.; Lertpanyasampatha, M.; Sojikul, P.; Viboonjun, U.; Narangajavana, J. Computational identification of microRNAs and their targets in cassava (Manihot esculenta Crantz.). Mol. Biotechnol. 2013, 53, 257–269. [Google Scholar] [CrossRef]
- Dai, X.; Zhuang, Z.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server (2017 release). Nucleic Acids Res. 2018, 46, W49–W54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Sherman, B.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Doncheva, N.A.-O.; Morris, J.A.-O.; Gorodkin, J.; Jensen, L.A.-O.X. Cytoscape stringapp: Network analysis and visualization of proteomics data. J. Proteome Res. 2019, 18, 623–632. [Google Scholar] [CrossRef]
- Meghwal, M.; Goswami, T.K. Piper nigrum and piperine: An update. Phytother. Res. 2013, 27, 1121–1130. [Google Scholar] [CrossRef] [PubMed]
- Smilkov, K.; Ackova, D.G.; Cvetkovski, A.; Ruskovska, T.; Vidovic, B.; Atalay, M. Piperine: Old spice and new nutraceutical? Curr. Pharm. Des. 2019, 25, 1729–1739. [Google Scholar] [CrossRef]
- Panchy, N.A.-O.; Lehti-Shiu, M.A.-O.; Shiu, S.A.-O.X. Evolution of gene duplication in plants. Plant Physiol. 2016, 171, 2294–2316. [Google Scholar] [CrossRef] [Green Version]
- Ren, R.; Wang, H.; Guo, C.; Zhang, N.; Zeng, L.; Chen, Y.; Ma, H.; Qi, J. Widespread whole genome duplications contribute to genome complexity and species diversity in angiosperms. Mol. Plant 2018, 11, 414–428. [Google Scholar] [CrossRef] [Green Version]
- Xing, S.; Quodt, V.; Chandler, J.; Höhmann, S.; Berndtgen, R.; Huijser, P. SPL8 acts together with the brassinosteroid-signaling component bim1 in controlling Arabidopsis thaliana male fertility. Plants 2013, 2, 416–428. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Schwarz, S.; Saedler, H.; Huijser, P. SPL8, a local regulator in a subset of gibberellin-mediated developmental processes in Arabidopsis. Plant Mol. Biol. 2007, 63, 429–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.D.; Ling, L.Z.; Yi, T.S. Evolution and divergence of SBP-box genes in land plants. BMC Genom. 2015, 16, 787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.W.; Czech, B.; Weigel, D. miR156-regulated SPL transcription factors define an endogenous flowering pathway in Arabidopsis thaliana. Cell 2009, 138, 738–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]

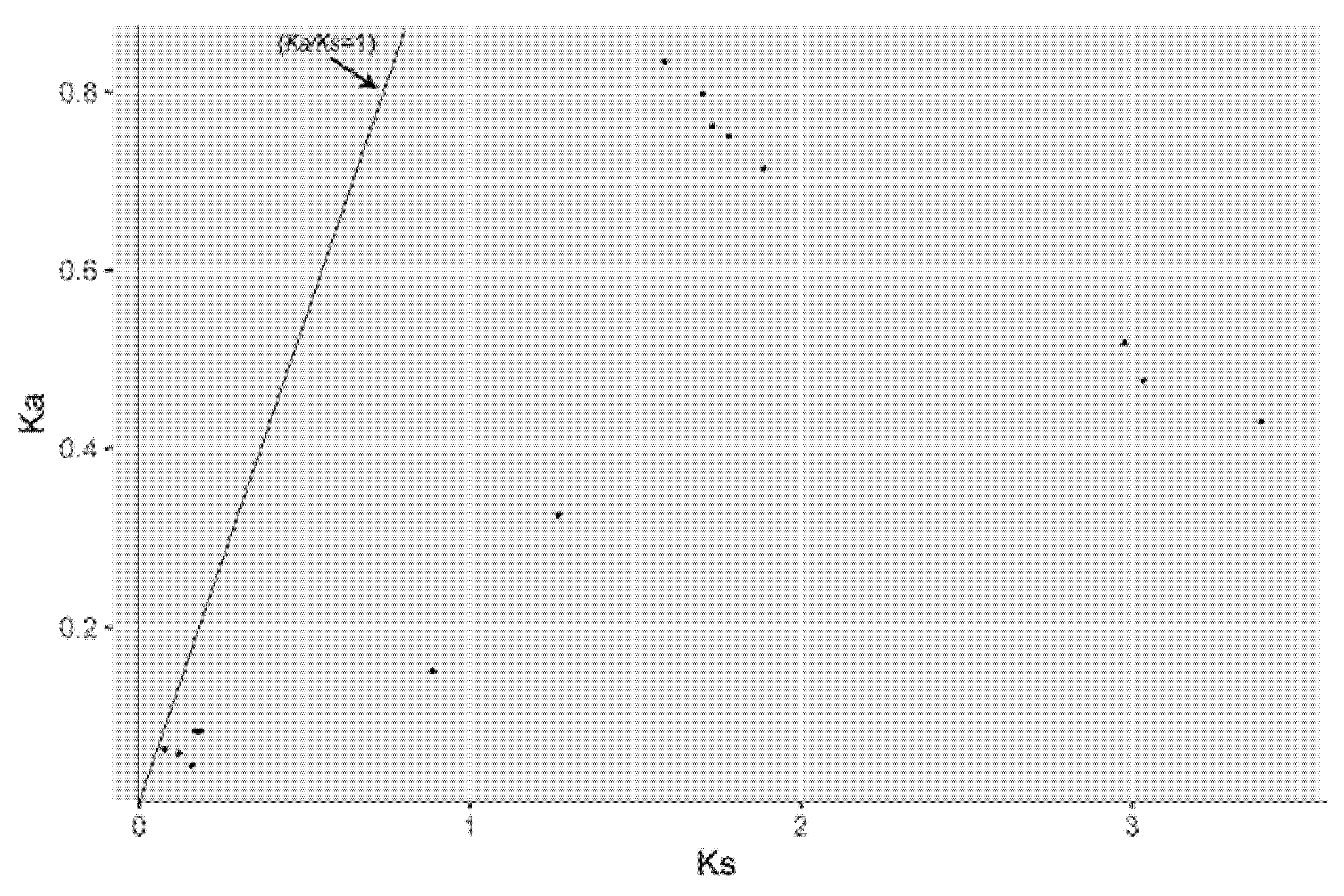


{kind=link}
{kind=link}
{kind=link}
{kind=link}
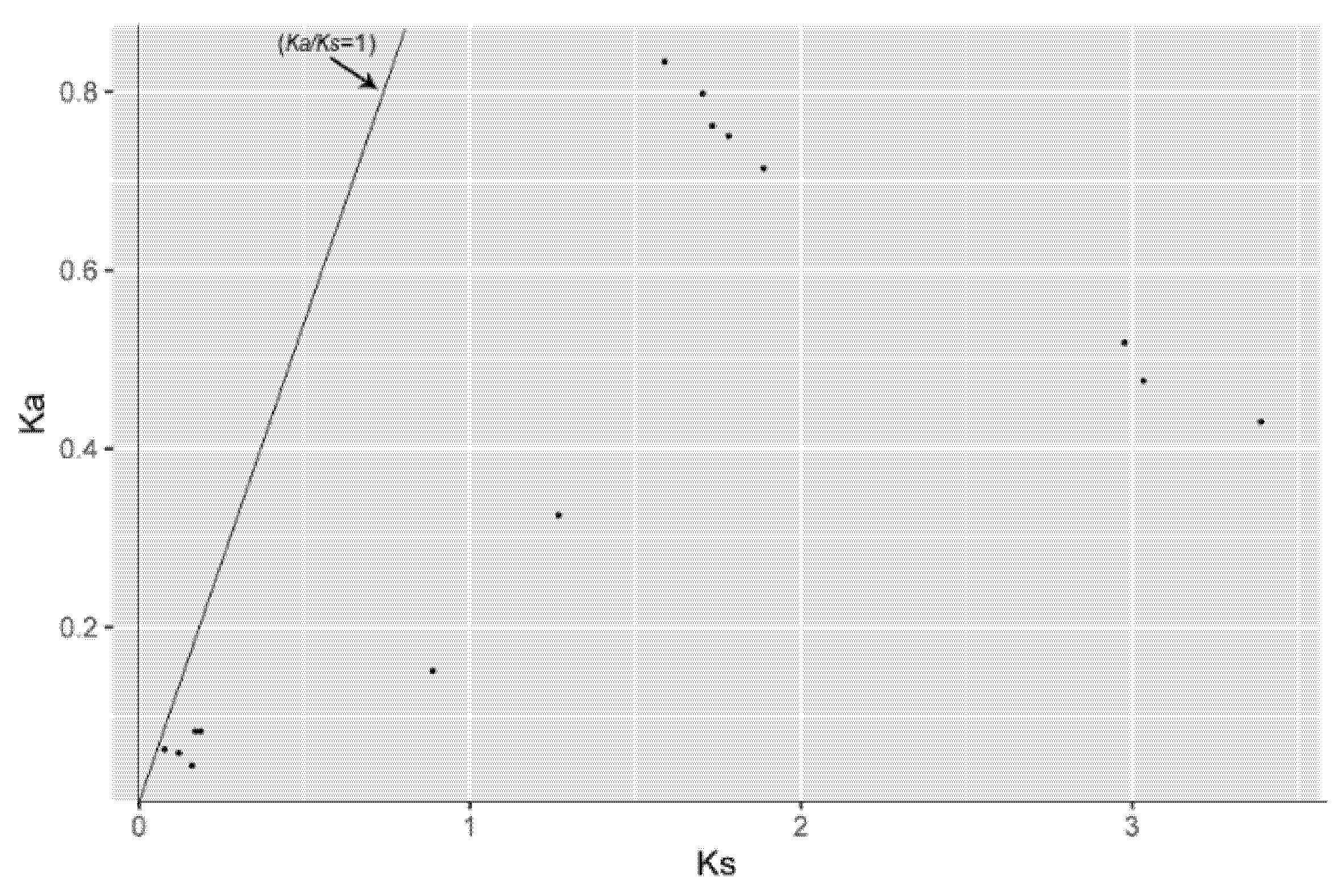{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Names | Gene | Length (aa) | MV (kDa) | pI |
|---|---|---|---|---|
| PnSBP8 | Pn14.164 | 337 | 36.7 | 8.72 |
| PnSBP13 | Pn15.136 | 354 | 38.2 | 8.62 |
| PnSBP4 | Pn7.344 | 365 | 40.3 | 8.72 |
| PnSBP28 | Pn10.2075 | 999 | 110.2 | 7.29 |
| PnSBP34 | Pn5.1721 | 787 | 88.1 | 6.40 |
| PnSBP33 | Pn5.1651 | 787 | 88.0 | 6.47 |
| PnSBP32 | Pn18.304 | 774 | 86.1 | 6.65 |
| PnSBP16 | Pn17.1041 | 269 | 30.7 | 9.61 |
| PnSBP9 | Pn5.2134 | 266 | 30.1 | 8.49 |
| PnSBP26 | Pn28.549 | 1038 | 114.4 | 6.30 |
| PnSBP27 | Pn28.583 | 1101 | 120.9 | 6.80 |
| PnSBP29 | Pn12.1036 | 1091 | 120.6 | 7.91 |
| PnSBP15 | Pn1.1351 | 203 | 23.2 | 10.28 |
| PnSBP1 | Pn1.2504 | 389 | 44.3 | 8.05 |
| PnSBP17 | Pn11.1178 | 368 | 40.0 | 8.44 |
| PnSBP18 | Pn13.776 | 396 | 43.8 | 9.62 |
| PnSBP22 | Pn14.389 | 344 | 38.7 | 7.63 |
| PnSBP12 | Pn19.85 | 289 | 30.9 | 8.94 |
| PnSBP5 | Pn2.1033 | 732 | 80.1 | 7.82 |
| PnSBP6 | Pn2.1339 | 476 | 52.3 | 8.73 |
| PnSBP7 | Pn2.939 | 713 | 78.9 | 8.66 |
| PnSBP14 | Pn21.1417 | 370 | 39.7 | 7.76 |
| PnSBP2 | Pn3.1618 | 581 | 64.4 | 8.54 |
| PnSBP10 | Pn3.4074 | 926 | 102.0 | 6.38 |
| PnSBP11 | Pn3.426 | 369 | 39.7 | 9.42 |
| PnSBP3 | Pn4.2537 | 323 | 35.9 | 8.43 |
| PnSBP23 | Pn5.2416 | 346 | 38.7 | 7.97 |
| PnSBP30 | Pn6.1128 | 177 | 19.8 | 9.47 |
| PnSBP24 | Pn6.473 | 274 | 29.8 | 8.75 |
| PnSBP20 | Pn6.475 | 327 | 35.5 | 8.43 |
| PnSBP21 | Pn6.937 | 327 | 35.6 | 8.37 |
| PnSBP25 | Pn6.939 | 274 | 29.8 | 8.75 |
| PnSBP19 | Pn8.2451 | 322 | 35.2 | 9.25 |
| PnSBP31 | Pn8.2831 | 177 | 19.9 | 9.18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Fan, R.; Wu, B.; Ji, X.; Hao, C. Genome-Wide Identification and Functional Exploration of SBP-Box Gene Family in Black Pepper (Piper nigrum L.). Genes 2021, 12, 1740. https://doi.org/10.3390/genes12111740
Li J, Fan R, Wu B, Ji X, Hao C. Genome-Wide Identification and Functional Exploration of SBP-Box Gene Family in Black Pepper (Piper nigrum L.). Genes. 2021; 12(11):1740. https://doi.org/10.3390/genes12111740
Chicago/Turabian StyleLi, Jing, Rui Fan, Baoduo Wu, Xunzhi Ji, and Chaoyun Hao. 2021. "Genome-Wide Identification and Functional Exploration of SBP-Box Gene Family in Black Pepper (Piper nigrum L.)" Genes 12, no. 11: 1740. https://doi.org/10.3390/genes12111740
APA StyleLi, J., Fan, R., Wu, B., Ji, X., & Hao, C. (2021). Genome-Wide Identification and Functional Exploration of SBP-Box Gene Family in Black Pepper (Piper nigrum L.). Genes, 12(11), 1740. https://doi.org/10.3390/genes12111740