Why Do Some Sex Chromosomes Degenerate More Slowly Than Others? The Odd Case of Ratite Sex Chromosomes


Abstract
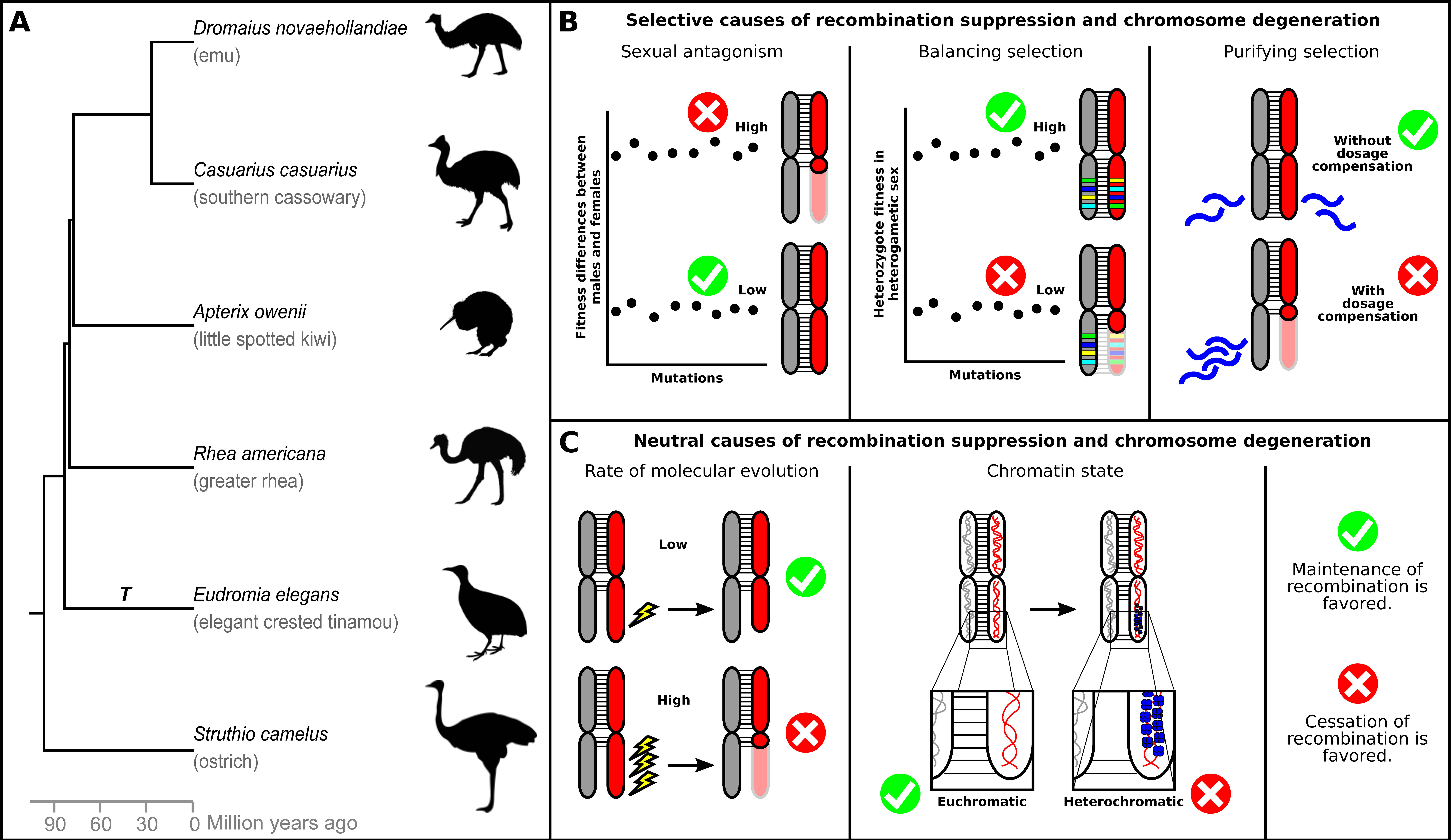
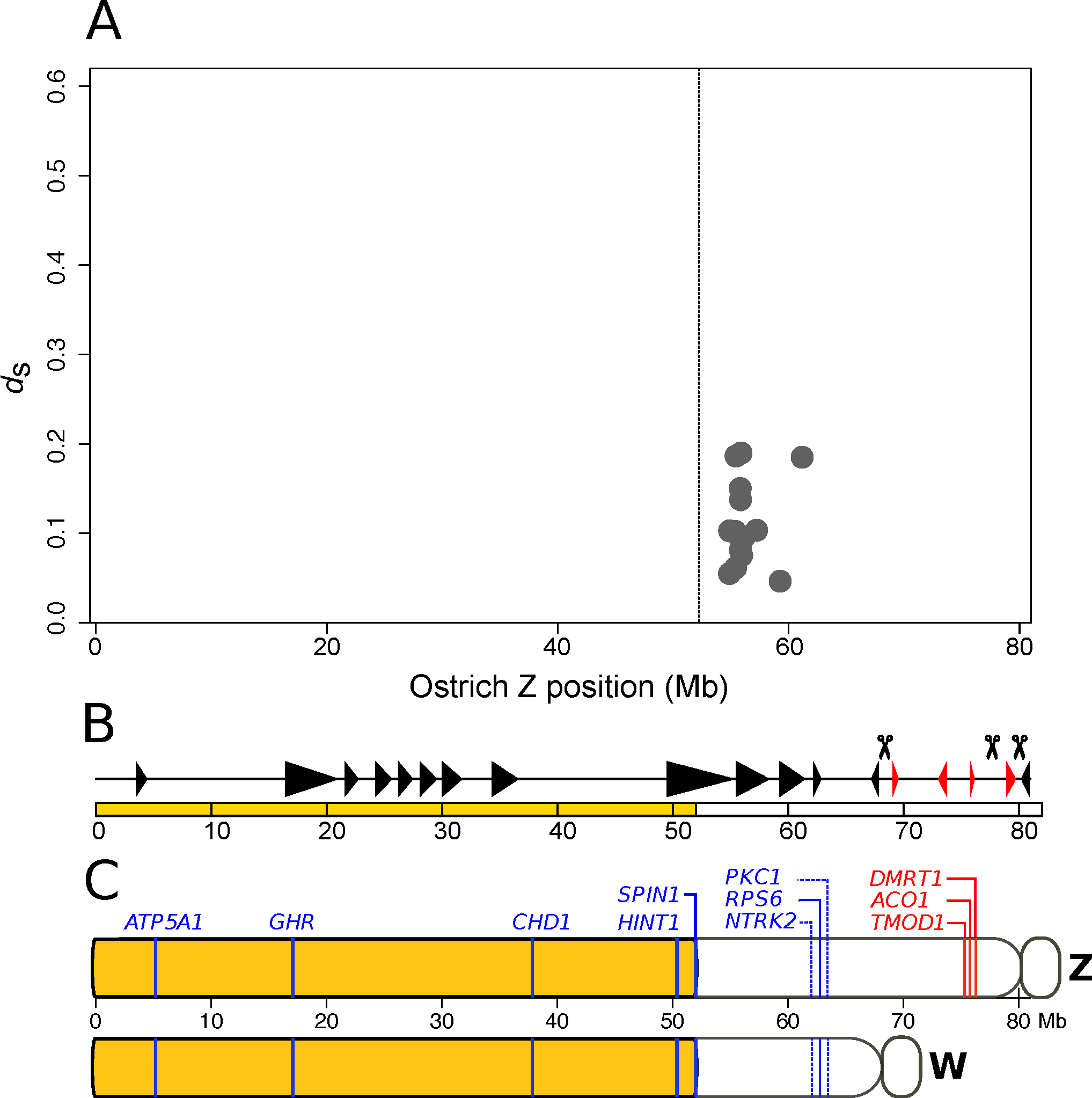
1. Introduction
2. Selective Processes
2.1. Sexually Antagonistic Selection
2.2. Balancing Selection
2.3. Purifying Selection
3. Neutral Processes
3.1. Rate of Molecular Evolution
3.2. Chromatin State of Sex Chromosomes
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sinclair, A.H.; Berta, P.; Palmer, M.S.; Hawkins, J.R.; Griffiths, B.L.; Smith, M.J.; Foster, J.W.; Frischauf, A.M.; Lovell-Badge, R.; Goodfellow, P.N. A gene from the human sex-determining region encodes a protein with homology to a conserved DNA-binding motif. Nature 1990, 346, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.A.; Roeszler, K.N.; Ohnesorg, T.; Cummins, D.M.; Farlie, P.G.; Doran, T.J.; Sinclair, A.H. The avian Z-linked gene DMRT1 is required for male sex determination in the chicken. Nature 2009, 461, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Fridolfsson, A.K.; Cheng, H.; Copeland, N.G.; Jenkins, N.A.; Liu, H.C.; Raudsepp, T.; Woodage, T.; Chowdhary, B.; Halverson, J.; Ellegren, H. Evolution of the avian sex chromosomes from an ancestral pair of autosomes. Proc. Natl. Acad. Sci. USA 1998, 95, 8147–8152. [Google Scholar] [CrossRef]
- Bellott, D.W.; Skaletsky, H.; Cho, T.J.; Brown, L.; Locke, D.; Chen, N.; Galkina, S.; Pyntikova, T.; Koutseva, N.; Graves, T.; et al. Avian W and mammalian Y chromosomes convergently retained dosage-sensitive regulators. Nat. Genet. 2017, 49, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Itoh, Y.; Mizuno, S. Molecular and cytological characterization of SspI-family repetitive sequence on the chicken W chromosome. Chromosome Res. 2002, 10, 499–511. [Google Scholar] [CrossRef]
- Saitoh, Y.; Saitoh, H.; Ohtomo, K.; Mizuno, S. Occupancy of the majority of DNA in the chicken W chromosome by bent-repetitive sequences. Chromosoma 1991, 101, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Stefos, K.; Arrighi, F.E. Heterochromatic nature of W chromosome in birds. Exp. Cell Res. 1971, 68, 228–231. [Google Scholar] [CrossRef]
- Otto, S.P.; Pannell, J.R.; Peichel, C.L.; Ashman, T.L.; Charlesworth, D.; Chippindale, A.K.; Delph, L.F.; Guerrero, R.F.; Scarpino, S.V.; McAllister, B.F. About PAR: The distinct evolutionary dynamics of the pseudoautosomal region. Trends Genet. 2011, 27, 358–367. [Google Scholar] [CrossRef]
- Smeds, L.; Kawakami, T.; Burri, R.; Bolivar, P.; Husby, A.; Qvarnstrom, A.; Uebbing, S.; Ellegren, H. Genomic identification and characterization of the pseudoautosomal region in highly differentiated avian sex chromosomes. Nat. Commun. 2014, 5, 5448. [Google Scholar] [CrossRef]
- Hinch, A.G.; Altemose, N.; Noor, N.; Donnelly, P.; Myers, S.R. Recombination in the human Pseudoautosomal region PAR1. PLoS Genet. 2014, 10, e1004503. [Google Scholar] [CrossRef]
- Ponnikas, S.; Sigeman, H.; Abbott, J.K.; Hansson, B. Why Do Sex Chromosomes Stop Recombining? Trends Genet. 2018, 34, 492–503. [Google Scholar] [CrossRef] [PubMed]
- Lenormand, T. The evolution of sex dimorphism in recombination. Genetics 2003, 163, 811–822. [Google Scholar] [PubMed]
- Rice, W.R. The Accumulation of Sexually Antagonistic Genes as a Selective Agent Promoting the Evolution of Reduced Recombination between Primitive Sex-Chromosomes. Evolution 1987, 41, 911–914. [Google Scholar] [CrossRef] [PubMed]
- Nei, M. Linkage modifications and sex difference in recombination. Genetics 1969, 63, 681–699. [Google Scholar] [PubMed]
- Charlesworth, B.; Charlesworth, D. A Model for the Evolution of Dioecy and Gynodioecy. Am. Nat. 1978, 112, 975–997. [Google Scholar] [CrossRef]
- Charlesworth, B.; Charlesworth, D. The degeneration of Y chromosomes. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2000, 355, 1563–1572. [Google Scholar] [CrossRef]
- Muller, H.J. The Relation of Recombination to Mutational Advance. Mutat. Res. 1964, 106, 2–9. [Google Scholar] [CrossRef]
- Felsenstein, J. The evolutionary advantage of recombination. Genetics 1974, 78, 737–756. [Google Scholar]
- Charlesworth, B.; Morgan, M.T.; Charlesworth, D. The effect of deleterious mutations on neutral molecular variation. Genetics 1993, 134, 1289–1303. [Google Scholar]
- Maynard-Smith, J.; Haigh, J. Hitch-Hiking Effect of a Favorable Gene. Genet. Res. 1974, 23, 23–35. [Google Scholar] [CrossRef]
- Hill, W.G.; Robertson, A. Effect of Linkage on Limits to Artificial Selection. Genet. Res. 1966, 8, 269–294. [Google Scholar] [CrossRef] [PubMed]
- Smeds, L.; Warmuth, V.; Bolivar, P.; Uebbing, S.; Burri, R.; Suh, A.; Nater, A.; Bures, S.; Garamszegi, L.Z.; Hogner, S.; et al. Evolutionary analysis of the female-specific avian W chromosome. Nat. Commun. 2015, 6, 7330. [Google Scholar] [CrossRef]
- Xu, L.; Wa Sin, S.Y.; Grayson, P.; Edwards, S.V.; Sackton, T.B. Evolutionary Dynamics of Sex Chromosomes of Paleognathous Birds. Genome Biol. Evol. 2019, 11, 2376–2390. [Google Scholar] [CrossRef] [PubMed]
- Peona, V.; Palacios-Gimenez, O.M.; Blommaert, J.; Liu, J.; Haryoko, T.; Jønsson, K.A.; Irestedt, M.; Zhou, Q.; Jern, P.; Suh, A. The avian W chromosome is a refugium for endogenous retroviruses with likely effects on female-biased mutational load and genetic incompatibilities. BioRxiv 2020. [Google Scholar] [CrossRef]
- Yazdi, H.P.; Ellegren, H. A Genetic Map of Ostrich Z Chromosome and the Role of Inversions in Avian Sex Chromosome Evolution. Genome Biol. Evol. 2018, 10, 2049–2060. [Google Scholar] [CrossRef]
- del Priore, L.; Pigozzi, M.I. Broad-scale recombination pattern in the primitive bird Rhea americana (Ratites, Palaeognathae). PLoS ONE 2017, 12, e0187549. [Google Scholar] [CrossRef]
- Takagi, N.; Ito, M.; Sasaki, M. Chromosome studies in four species of Ratitae (Aves). Chromosoma 1972, 36, 281–291. [Google Scholar] [CrossRef]
- Ansari, H.A.; Takagi, N.; Sasaki, M. Morphological-Differentiation of Sex-Chromosomes in 3 Species of Ratite Birds. Cytogenet. Cell Genet. 1988, 47, 185–188. [Google Scholar] [CrossRef]
- Nanda, I.; Schlegelmilch, K.; Haaf, T.; Schartl, M.; Schmid, M. Synteny conservation of the Z chromosome in 14 avian species (11 families) supports a role for Z dosage in avian sex determination. Cytogenet. Genome Res. 2008, 122, 150–156. [Google Scholar] [CrossRef]
- Tsuda, Y.; Nishida-Umehara, C.; Ishijima, J.; Yamada, K.; Matsuda, Y. Comparison of the Z and W sex chromosomal architectures in elegant crested tinamou (Eudromia elegans) and ostrich (Struthio camelus) and the process of sex chromosome differentiation in palaeognathous birds. Chromosoma 2007, 116, 159–173. [Google Scholar] [CrossRef]
- Pigozzi, M.I. Diverse stages of sex-chromosome differentiation in tinamid birds: Evidence from crossover analysis in Eudromia elegans and Crypturellus tataupa. Genetica 2011, 139, 771–777. [Google Scholar] [CrossRef] [PubMed]
- Schartl, M. Sex chromosome evolution in non-mammalian vertebrates. Curr. Opin. Genet. Dev. 2004, 14, 634–641. [Google Scholar] [CrossRef] [PubMed]
- Perrin, N. Sex reversal: A fountain of youth for sex chromosomes? Evolution 2009, 63, 3043–3049. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.J.; Haddrath, O.; McPherson, J.D.; Cloutier, A. Genomic support for a moa-tinamou clade and adaptive morphological convergence in flightless ratites. Mol. Biol. Evol. 2014, 31, 1686–1696. [Google Scholar] [CrossRef] [PubMed]
- Cloutier, A.; Sackton, T.B.; Grayson, P.; Clamp, M.; Baker, A.J.; Edwards, S.V. Whole-Genome Analyses Resolve the Phylogeny of Flightless Birds (Palaeognathae) in the Presence of an Empirical Anomaly Zone. Syst. Biol. 2019, 68, 937–955. [Google Scholar] [CrossRef]
- Zhou, Q.; Zhang, J.L.; Bachtrog, D.; An, N.; Huang, Q.F.; Jarvis, E.D.; Gilbert, M.T.P.; Zhang, G.J. Complex evolutionary trajectories of sex chromosomes across bird taxa. Science 2014, 346, 1332–1342. [Google Scholar] [CrossRef]
- Yazdi, H.P.; Ellegren, H. Old but not (so) degenerated—Slow evolution of largely homomorphic sex chromosomes in ratites. Mol. Biol. Evol. 2014, 31, 1444–1453. [Google Scholar] [CrossRef]
- Shetty, S.; Kirby, P.; Zarkower, D.; Graves, J.A. DMRT1 in a ratite bird: Evidence for a role in sex determination and discovery of a putative regulatory element. Cytogenet. Genome Res. 2002, 99, 245–251. [Google Scholar] [CrossRef]
- Vicoso, B.; Kaiser, V.B.; Bachtrog, D. Sex-biased gene expression at homomorphic sex chromosomes in emus and its implication for sex chromosome evolution. Proc. Natl. Acad. Sci. USA 2013, 110, 6453–6458. [Google Scholar] [CrossRef]
- Jarvis, E.D.; Mirarab, S.; Aberer, A.J.; Li, B.; Houde, P.; Li, C.; Ho, S.Y.; Faircloth, B.C.; Nabholz, B.; Howard, J.T.; et al. Whole-genome analyses resolve early branches in the tree of life of modern birds. Science 2014, 346, 1320–1331. [Google Scholar] [CrossRef]
- Schartl, M.; Schmid, M.; Nanda, I. Dynamics of vertebrate sex chromosome evolution: From equal size to giants and dwarfs. Chromosoma 2016, 125, 553–571. [Google Scholar] [CrossRef] [PubMed]
- Coddington, C.L.; Cockburn, A. The Mating System of Free-Living Emus. Aust. J. Zool. 1995, 43, 365–372. [Google Scholar] [CrossRef]
- Codenotti, T.L.; Alvarez, F. Mating behavior of the male Greater Rhea. Wilson Bull. 2001, 113, 85–89. [Google Scholar] [CrossRef]
- del Hoyo, J.; Elliott, A.; Sargatal, J.; Christie, D.A.; Kirwan, G.E. Handbook of the Birds of the World Alive; Lynx Edicions: Barcelona, Spain, 2019. [Google Scholar]
- IUCN 2019. The IUCN Red List of Threatened Species. Version 2019-3. Available online: http://www.iucnredlist.org (accessed on 10 December 2019).
- Myers, P.; Espinosa, R.; Parr, C.S.; Jones, T.; Hammond, G.S.; Dewey, T.A. The Animal Diversity Web. Available online: https://animaldiversity.org/ (accessed on 10 December 2019).
- Kawakami, T.; Smeds, L.; Backstrom, N.; Husby, A.; Qvarnstrom, A.; Mugal, C.F.; Olason, P.; Ellegren, H. A high-density linkage map enables a second-generation collared flycatcher genome assembly and reveals the patterns of avian recombination rate variation and chromosomal evolution. Mol. Ecol. 2014, 23, 4035–4058. [Google Scholar] [CrossRef] [PubMed]
- Otto, S.P. Selective maintenance of recombination between the sex chromosomes. J. Evol. Biol. 2014, 27, 1431–1442. [Google Scholar] [CrossRef]
- Claramunt, S.; Cracraft, J. A new time tree reveals Earth history’s imprint on the evolution of modern birds. Sci. Adv. 2015, 1, e1501005. [Google Scholar] [CrossRef]
- Torres, E.M.; Williams, B.R.; Amon, A. Aneuploidy: Cells losing their balance. Genetics 2008, 179, 737–746. [Google Scholar] [CrossRef]
- Tang, Y.C.; Amon, A. Gene Copy-Number Alterations: A Cost-Benefit Analysis. Cell 2013, 152, 394–405. [Google Scholar] [CrossRef]
- Khan, Z.; Ford, M.J.; Cusanovich, D.A.; Mitrano, A.; Pritchard, J.K.; Gilad, Y. Primate transcript and protein expression levels evolve under compensatory selection pressures. Science 2013, 342, 1100–1104. [Google Scholar] [CrossRef]
- Ishikawa, K.; Makanae, K.; Iwasaki, S.; Ingolia, N.T.; Moriya, H. Post-Translational Dosage Compensation Buffers Genetic Perturbations to Stoichiometry of Protein Complexes. PLoS Genet. 2017, 13, e1006554. [Google Scholar] [CrossRef]
- Dang, V.T.; Kassahn, K.S.; Marcos, A.E.; Ragan, M.A. Identification of human haploinsufficient genes and their genomic proximity to segmental duplications. Eur. J. Hum. Genet. 2008, 16, 1350–1357. [Google Scholar] [CrossRef] [PubMed]
- Graves, J.A.M. Evolution of vertebrate sex chromosomes and dosage compensation. Nat. Rev. Genet. 2016, 17, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Brockdorff, N.; Turner, B.M. Dosage compensation in mammals. Cold Spring Harb. Perspect. Biol. 2015, 7, a019406. [Google Scholar] [CrossRef]
- Mank, J.E.; Ellegren, H. All dosage compensation is local: Gene-by-gene regulation of sex-biased expression on the chicken Z chromosome. Heredity 2009, 102, 312–320. [Google Scholar] [CrossRef]
- Uebbing, S.; Kunstner, A.; Makinen, H.; Ellegren, H. Transcriptome sequencing reveals the character of incomplete dosage compensation across multiple tissues in flycatchers. Genome Biol. Evol. 2013, 5, 1555–1566. [Google Scholar] [CrossRef]
- Adolfsson, S.; Ellegren, H. Lack of Dosage Compensation Accompanies the Arrested Stage of Sex Chromosome Evolution in Ostriches. Mol. Biol. Evol. 2013, 30, 806–810. [Google Scholar] [CrossRef]
- Stuart, R.O.; Bush, K.T.; Nigam, S.K. Changes in global gene expression patterns during development and maturation of the rat kidney. Proc. Natl. Acad. Sci. USA 2001, 98, 5649–5654. [Google Scholar] [CrossRef]
- Bellott, D.W.; Page, D.C. Dosage-sensitive functions in embryonic development drove the survival of genes on sex-specific chromosomes in snakes, birds, and mammals. BioRxiv 2020. [Google Scholar] [CrossRef]
- Lahn, B.T.; Page, D.C. Four evolutionary strata on the human X chromosome. Science 1999, 286, 964–967. [Google Scholar] [CrossRef]
- Huang, Y.; Liu, Q.; Tang, B.; Lin, L.; Liu, W.; Zhang, L.; Li, N.; Hu, X. A preliminary microsatellite genetic map of the ostrich (Struthio camelus). Cytogenet. Genome Res. 2008, 121, 130–136. [Google Scholar] [CrossRef]
- Figuet, E.; Nabholz, B.; Bonneau, M.; Carrio, E.M.; Nadachowska-Brzyska, K.; Ellegren, H.; Galtier, N. Life History Traits, Protein Evolution, and the Nearly Neutral Theory in Amniotes. Mol. Biol. Evol. 2016, 33, 1517–1527. [Google Scholar] [CrossRef]
- Lartillot, N.; Poujol, R. A phylogenetic model for investigating correlated evolution of substitution rates and continuous phenotypic characters. Mol. Biol. Evol. 2011, 28, 729–744. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.J.; Li, C.; Li, Q.Y.; Li, B.; Larkin, D.M.; Lee, C.; Storz, J.F.; Antunes, A.; Greenwold, M.J.; Meredith, R.W.; et al. Comparative genomics reveals insights into avian genome evolution and adaptation. Science 2014, 346, 1311–1320. [Google Scholar] [CrossRef]
- Kapusta, A.; Suh, A.; Feschotte, C. Dynamics of genome size evolution in birds and mammals. Proc. Natl. Acad. Sci. USA 2017, 114, E1460–E1469. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, J.; Xu, X.; Witt, C.; Deng, Y.; Chen, G.; Meng, G.; Feng, S.; Szekely, T.; Zhang, G.; et al. Phylogeny, transposable element and sex chromosome evolution of the basal lineage of birds. BioRxiv 2019. [Google Scholar] [CrossRef]
- Konkel, M.K.; Batzer, M.A. A mobile threat to genome stability: The impact of non-LTR retrotransposons upon the human genome. In Seminars in Cancer Biology; Academic Press: New York, NY, USA, 2010; Volume 20, pp. 211–221. [Google Scholar]
- Darolti, I.; Wright, A.E.; Mank, J.E. Guppy Y Chromosome Integrity Maintained by Incomplete Recombination Suppression. Genome Biol. Evol. 2020, 12, 965–977. [Google Scholar] [CrossRef]
- Kent, T.V.; Uzunovic, J.; Wright, S.I. Coevolution between transposable elements and recombination. Philos. Trans. R. Soc. B 2017, 372, 20160458. [Google Scholar] [CrossRef]
- Choi, J.Y.; Lee, Y.C.G. Double-edged sword: The evolutionary consequences of the epigenetic silencing of transposable elements. PLoS Genet. 2020, 16, e1008872. [Google Scholar] [CrossRef]
- Hollister, J.D.; Gaut, B.S. Epigenetic silencing of transposable elements: A trade-off between reduced transposition and deleterious effects on neighboring gene expression. Genome Res. 2009, 19, 1419–1428. [Google Scholar] [CrossRef]
- Lee, Y.C.G.; Karpen, G.H. Pervasive epigenetic effects of Drosophila euchromatic transposable elements impact their evolution. Elife 2017, 6, e25762. [Google Scholar] [CrossRef]
- Kirkpatrick, M. How and why chromosome inversions evolve. PLoS Biol. 2010, 8, e1000501. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, M.; Guerrero, R.F.; Scarpino, S.V. Patterns of neutral genetic variation on recombining sex chromosomes. Genetics 2010, 184, 1141–1152. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, M.; Guerrero, R.F. Signatures of sex-antagonistic selection on recombining sex chromosomes. Genetics 2014, 197, 531–541. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Species | Chromosome Number (2n) | Centromere Position ZW | W Chromatin State | PAR Length | Gametolog dS Range | Gametolog dNdS Range |
|---|---|---|---|---|---|---|
| Ostrich | 80 1 | Acrocentric 4 | Euchromatic 5 | 52.5 Mb 8 | 0.036–2.616 6,7 | 0.001–0.858 6,7 |
| Emu | 80 2 | Acrocentric 2 | Euchromatic 2 | 59.3 Mb 8 | 0.128–1.435 6 | 0.015–0.917 6 |
| Cassowary | - | - | - | 59.3 Mb 8 | 0.016–0.019 8 | 0.218–0.273 8 |
| Little spotted kiwi | - | - | - | 53.6 Mb 8 | 0.002–0.002 8 | 0.191–0.268 8 |
| Greater rhea | 82 2 | Acrocentric 2 | Euchromatic 2 | 52.55 Mb 8 | 0.007–0.008 8 | 0.175–0.225 8 |
| Elegant crested tinamou | 80 3 | Acrocentric 3 | Heterochromatic with terminal euchromatic segment on its long arm 3 | 32.2 Mb 8 | 0.017–0.706 6 | 0.001–2.064 6 |
| Tataupa tinamou | 80 3 | Acrocentric 3 | Completely heterochromatic 3 | - | - | - |
| Species | Mating System | Generation Time (Year) | Sexual Maturity (Year) | Body Mass (kg) |
|---|---|---|---|---|
| Ostrich | Polygynandrous * | 16.8 † | 3–4 * | 100–156 (male) * 90–110 (female) * |
| Emu | Females successive polyandry * | 10.5 † | 2–3 * | 30–55 * |
| Cassowary | Polyandrous * | 12.5 † | 3 ‡ | 29–34 (male) * 58 (female) * |
| Little spotted kiwi | Monogamous * | 9 † | 2–3 * | 0.88–1.356 (male) * 1–1.950 (female) * |
| Greater rhea | Males simultaneously polygynous, females serially polyandrous * | 10.5 † | 2–3 * | 20–27 * |
| White-throated tinamou | - | 6.8 † | - | 0.623–0.652 (male) * 0.680–0.8 (female) * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yazdi, H.P.; Silva, W.T.A.F.; Suh, A. Why Do Some Sex Chromosomes Degenerate More Slowly Than Others? The Odd Case of Ratite Sex Chromosomes. Genes 2020, 11, 1153. https://doi.org/10.3390/genes11101153
Yazdi HP, Silva WTAF, Suh A. Why Do Some Sex Chromosomes Degenerate More Slowly Than Others? The Odd Case of Ratite Sex Chromosomes. Genes. 2020; 11(10):1153. https://doi.org/10.3390/genes11101153
Chicago/Turabian StyleYazdi, Homa Papoli, Willian T. A. F. Silva, and Alexander Suh. 2020. "Why Do Some Sex Chromosomes Degenerate More Slowly Than Others? The Odd Case of Ratite Sex Chromosomes" Genes 11, no. 10: 1153. https://doi.org/10.3390/genes11101153
APA StyleYazdi, H. P., Silva, W. T. A. F., & Suh, A. (2020). Why Do Some Sex Chromosomes Degenerate More Slowly Than Others? The Odd Case of Ratite Sex Chromosomes. Genes, 11(10), 1153. https://doi.org/10.3390/genes11101153