Impact of Biomaterials on Differentiation and Reprogramming Approaches for the Generation of Functional Cardiomyocytes

Abstract
1. Introduction
2. Cell Sources for the Generation of Cardiomyocytes
2.1. Embryonic Stem Cells (ESCs)
2.2. Induced Pluripotent Stem Cells (iPSCs)
2.3. Mesenchymal Stem Cells (MSCs)
2.4. Cardiac Progenitors Cells (CPCs)
2.5. Indirect and Direct Reprogramming
2.6. New Emerging Approaches for CM Generation
3. Techniques for the Evaluation of the Formation of Functional Cardiomyocytes
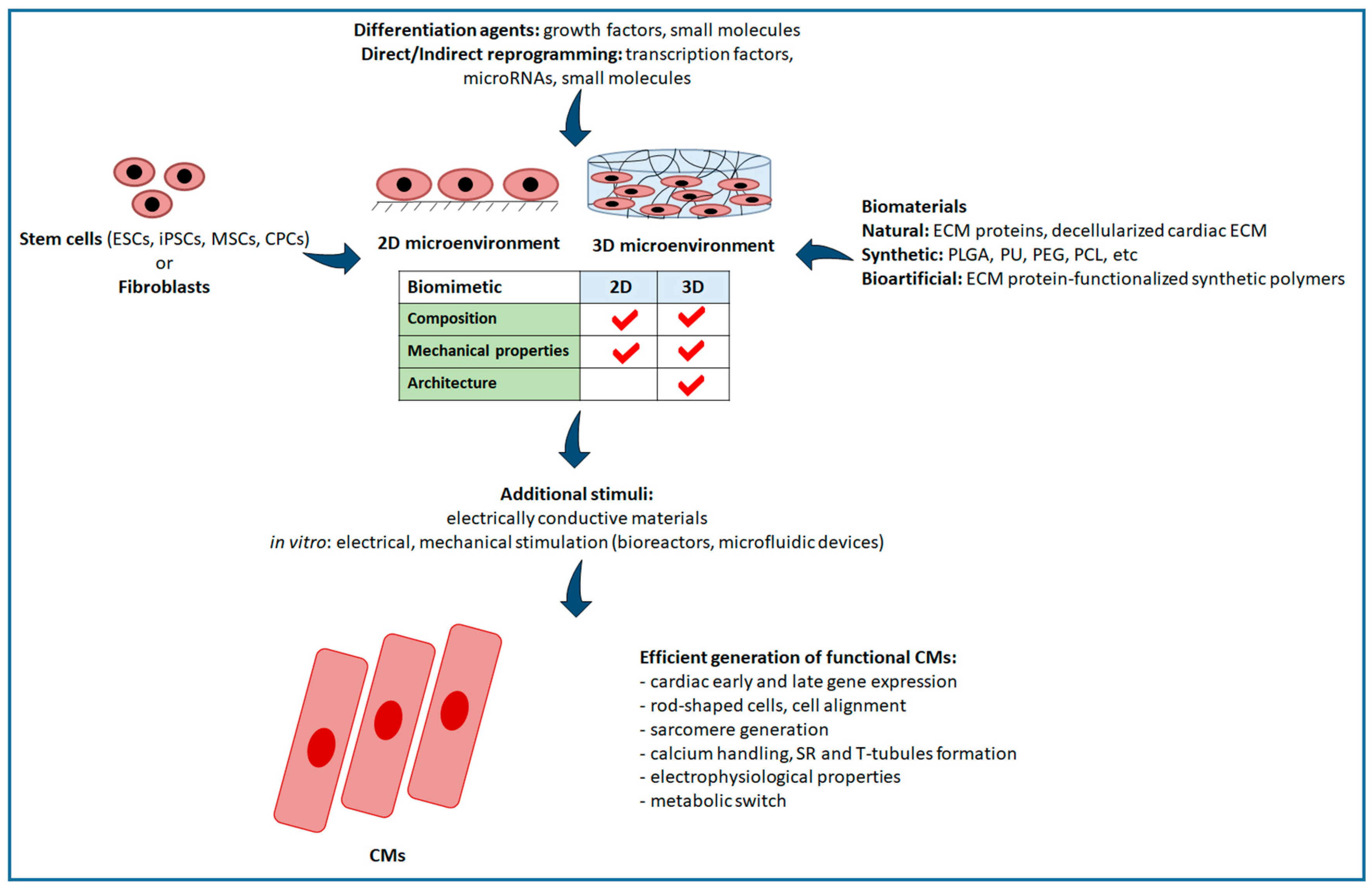
4. Biomaterials for Guiding Cell Behaviour
- (i)
- ECM-mimetic (mainly protein-based) biomaterials;
- (ii)
- decellularized cardiac ECM;
- (iii)
- “bioartificial” materials consisting of synthetic polymers functionalized with cardiac ECM proteins.
4.1. Role of Substrate on Stem Cell Differentiation into CMs
4.1.1. Biomaterials Supporting ESC Differentiation
4.1.2. Biomaterials Supporting iPSC Differentiation
4.1.3. Biomaterials Supporting MSC Differentiation
4.1.4. Biomaterials Supporting CPC Differentiation
4.2. Role of Substrate on Fibroblast Direct and Indirect Reprogramming into CMs
5. Critical Analysis of the State of the Art and Future Perspectives
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Addis, R.C.; Epstein, J.A. Induced regeneration—The progress and promise of direct reprogramming for heart repair. Nat. Med. 2013, 19, 829–836. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.-Z.; Harding, S.E.; Ali, N.N.; Lyon, A.R.; Boccaccini, A.R. Biomaterials in cardiac tissue engineering: Ten years of research survey. Mater. Sci. Eng. R Rep. 2008, 59, 1–37. [Google Scholar] [CrossRef]
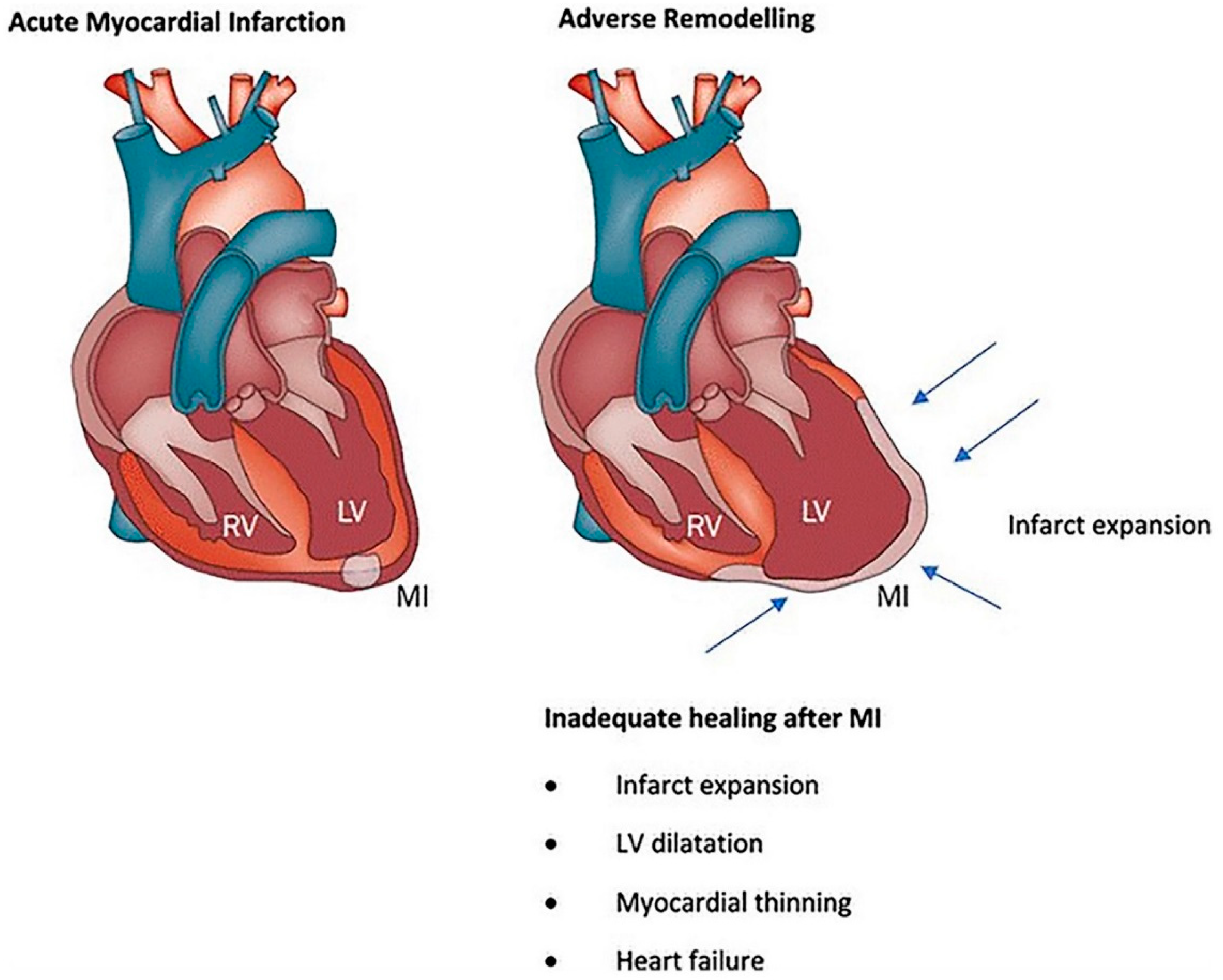
- Curley, D.; Lavin Plaza, B.; Shah, A.M.; Botnar, R.M. Molecular imaging of cardiac remodelling after myocardial infarction. Basic Res. Cardiol. 2018, 113, 10. [Google Scholar] [CrossRef] [PubMed]
- Fraccarollo, D.; Galuppo, P.; Bauersachs, J. Novel therapeutic approaches to post-infarction remodelling. Cardiovasc. Res. 2012, 94, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Pavesi, A.; Adriani, G.; Rasponi, M.; Zervantonakis, I.K.; Fiore, G.B.; Kamm, R.D. Controlled electromechanical cell stimulation on-a-chip. Sci. Rep. 2015, 5, 11800. [Google Scholar] [CrossRef] [PubMed]
- Segers, V.F.M.; Lee, R.T. Stem-cell therapy for cardiac disease. Nature 2008, 451, 937–942. [Google Scholar] [CrossRef] [PubMed]
- Ponikowski, P.; Anker, S.D.; AlHabib, K.F.; Cowie, M.R.; Force, T.L.; Hu, S.; Jaarsma, T.; Krum, H.; Rastogi, V.; Rohde, L.E.; et al. Heart failure: Preventing disease and death worldwide. ESC Heart Fail. 2014, 1, 4–25. [Google Scholar] [CrossRef] [PubMed]
- Silvestri, A.; Boffito, M.; Sartori, S.; Ciardelli, G. Biomimetic Materials and Scaffolds for Myocardial Tissue Regeneration. Macromol. Biosci. 2013, 13, 984–1019. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, J.; Krishnan, U.M.; Sethuraman, S. Hydrogel based injectable scaffolds for cardiac tissue regeneration. Biotechnol. Adv. 2014, 32, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Sanganalmath, S.K.; Bolli, R. Cell Therapy for Heart Failure. Circ. Res. 2013, 113, 810–834. [Google Scholar] [CrossRef] [PubMed]
- Hodgkinson, C.P.; Bareja, A.; Gomez, J.A.; Dzau, V.J. Emerging Concepts in Paracrine Mechanisms in Regenerative Cardiovascular Medicine and Biology. Circ. Res. 2016, 118, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Batty, J.A.; Lima, J.A.C.; Kunadian, V. Direct cellular reprogramming for cardiac repair and regeneration. Eur. J. Heart Fail. 2016, 18, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Doppler, S.; Deutsch, M.-A.; Lange, R.; Krane, M. Direct Reprogramming—The Future of Cardiac Regeneration? Int. J. Mol. Sci. 2015, 16, 17368–17393. [Google Scholar] [CrossRef] [PubMed]
- Nam, Y.-J.; Song, K.; Olson, E.N. Heart repair by cardiac reprogramming. Nat. Med. 2013, 19, 413–415. [Google Scholar] [CrossRef] [PubMed]
- Hodgkinson, C.P.; Kang, M.H.; Dal-Pra, S.; Mirotsou, M.; Dzau, V.J. MicroRNAs and Cardiac Regeneration. Circ. Res. 2015, 116, 1700–1711. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, B. In vivo reprogramming for heart regeneration: A glance at efficiency, environmental impacts, challenges and future directions. J. Mol. Cell. Cardiol. 2017, 108, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Duelen, R.; Sampaolesi, M. Stem Cell Technology in Cardiac Regeneration: A Pluripotent Stem Cell Promise. EBioMedicine 2017, 16, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic stem cell lines derived from human blastocysts. Science 1998, 282, 1145–1147. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.J.H.; Yang, X.; Don, C.W.; Minami, E.; Liu, Y.-W.; Weyers, J.J.; Mahoney, W.M.; Van Biber, B.; Cook, S.M.; Palpant, N.J.; et al. Human embryonic-stem-cell-derived cardiomyocytes regenerate non-human primate hearts. Nature 2014, 510, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Hartman, M.E.; Dai, D.-F.; Laflamme, M.A. Human pluripotent stem cells: Prospects and challenges as a source of cardiomyocytes for in vitro modeling and cell-based cardiac repair. Adv. Drug Deliv. Rev. 2016, 96, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.K.; Kalsan, M.; Kumar, N.; Saini, A.; Chandra, R. Induced pluripotent stem cells: Applications in regenerative medicine, disease modeling, and drug discovery. Front. Cell Dev. Biol. 2015, 3, 2. [Google Scholar] [CrossRef] [PubMed]
- Rojas, S.V.; Kensah, G.; Rotaermel, A.; Baraki, H.; Kutschka, I.; Zweigerdt, R.; Martin, U.; Haverich, A.; Gruh, I.; Martens, A. Transplantation of purified iPSC-derived cardiomyocytes in myocardial infarction. PLoS ONE 2017, 12, e0173222. [Google Scholar] [CrossRef] [PubMed]
- Casini, S.; Verkerk, A.O.; Remme, C.A. Human iPSC-Derived Cardiomyocytes for Investigation of Disease Mechanisms and Therapeutic Strategies in Inherited Arrhythmia Syndromes: Strengths and Limitations. Cardiovasc. Drugs Ther. 2017, 31, 325–344. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Singh, A.; Sen, D. Mesenchymal stem cells in cardiac regeneration: A detailed progress report of the last 6 years (2010–2015). Stem Cell Res. Ther. 2016, 7, 82. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, G.; Fox, J.; Ashton, B.; Middleton, J. Concise Review: Mesenchymal Stem Cells: Their Phenotype, Differentiation Capacity, Immunological Features, and Potential for Homing. Stem Cells 2007, 25, 2739–2749. [Google Scholar] [CrossRef] [PubMed]
- Miao, C.; Lei, M.; Hu, W.; Han, S.; Wang, Q. A brief review: The therapeutic potential of bone marrow mesenchymal stem cells in myocardial infarction. Stem Cell Res. Ther. 2017, 8, 242. [Google Scholar] [CrossRef] [PubMed]
- Rosland, G.V.; Svendsen, A.; Torsvik, A.; Sobala, E.; McCormack, E.; Immervoll, H.; Mysliwietz, J.; Tonn, J.-C.; Goldbrunner, R.; Lonning, P.E.; et al. Long-term Cultures of Bone Marrow-Derived Human Mesenchymal Stem Cells Frequently Undergo Spontaneous Malignant Transformation. Cancer Res. 2009, 69, 5331–5339. [Google Scholar] [CrossRef] [PubMed]
- Mauretti, A.; Spaans, S.; Bax, N.A.M.; Sahlgren, C.; Bouten, C.V.C. Cardiac Progenitor Cells and the Interplay with Their Microenvironment. Stem Cells Int. 2017, 2017, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Le, T.; Chong, J. Cardiac progenitor cells for heart repair. Cell Death Discov. 2016, 2, 16052. [Google Scholar] [CrossRef] [PubMed]
- Zaruba, M.M.; Soonpaa, M.; Reuter, S.; Field, L.J. Cardiomyogenic Potential of C-Kit+-Expressing Cells Derived from Neonatal and Adult Mouse Hearts. Circulation 2010, 121, 1992–2000. [Google Scholar] [CrossRef] [PubMed]
- Soonpaa, M.H.; Rubart, M.; Field, L.J. Challenges measuring cardiomyocyte renewal. Biochim. Biophys. Acta 2013, 1833, 799–803. [Google Scholar] [CrossRef] [PubMed]
- Leri, A.; Kajstura, J.; Anversa, P. Role of Cardiac Stem Cells in Cardiac Pathophysiology: A Paradigm Shift in Human Myocardial Biology. Circ. Res. 2011, 109, 941–961. [Google Scholar] [CrossRef] [PubMed]
- Anversa, P.; Kajstura, J.; Rota, M.; Leri, A. Regenerating new heart with stem cells. J. Clin. Investig. 2013, 123, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Loughran, J.H.; Elmore, J.B.; Waqar, M.; Chugh, A.R.; Bolli, R. Cardiac Stem Cells in Patients with Ischemic Cardiomyopathy: Discovery, Translation, and Clinical Investigation. Curr. Atheroscler. Rep. 2012, 14, 491–503. [Google Scholar] [CrossRef] [PubMed]
- Dey, D.; Han, L.; Bauer, M.; Sanada, F.; Oikonomopoulos, A.; Hosoda, T.; Unno, K.; De Almeida, P.; Leri, A.; Wu, J.C. Dissecting the Molecular Relationship Among Various Cardiogenic Progenitor Cells. Circ. Res. 2013, 112, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Hastings, C.L.; Roche, E.T.; Ruiz-Hernandez, E.; Schenke-Layland, K.; Walsh, C.J.; Duffy, G.P. Drug and cell delivery for cardiac regeneration. Adv. Drug Deliv. Rev. 2015, 84, 85–106. [Google Scholar] [CrossRef] [PubMed]
- van Berlo, J.H.; Kanisicak, O.; Maillet, M.; Vagnozzi, R.J.; Karch, J.; Lin, S.-C.J.; Middleton, R.C.; Marbán, E.; Molkentin, J.D. c-kit+ cells minimally contribute cardiomyocytes to the heart. Nature 2014, 509, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Srivastava, D. Direct Cardiac Reprogramming: From Developmental Biology to Cardiac Regeneration. Circ. Res. 2013, 113, 915–921. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, Z.; Zhao, Z.-A.; Shen, Z. Direct reprogramming of fibroblasts into cardiomyocytes. Stem Cell Res. Ther. 2017, 8, 118. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Tu, W.; Fu, Y.; Wang, J.; Xie, X. Chemical-induced cardiac reprogramming in vivo. Cell Res. 2018, 1. [Google Scholar] [CrossRef] [PubMed]
- Ieda, M.; Fu, J.-D.; Delgado-Olguin, P.; Vedantham, V.; Hayashi, Y.; Bruneau, B.G.; Srivastava, D. Direct Reprogramming of Fibroblasts into Functional Cardiomyocytes by Defined Factors. Cell 2010, 142, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Londono, P.; Cao, Y.; Sharpe, E.J.; Proenza, C.; O’Rourke, R.; Jones, K.L.; Jeong, M.Y.; Walker, L.A.; Buttrick, P.M.; et al. High-efficiency reprogramming of fibroblasts into cardiomyocytes requires suppression of pro-fibrotic signalling. Nat. Commun. 2015, 6, 8243. [Google Scholar] [CrossRef] [PubMed]
- Gnecchi, M.; Pisano, F.; Bariani, R. MicroRNA and Cardiac Regeneration. Adv. Exp. Med. Biol. 2015, 887, 119–141. [Google Scholar] [CrossRef] [PubMed]
- Jayawardena, T.M.; Egemnazarov, B.; Finch, E.A.; Zhang, L.; Payne, J.A.; Pandya, K.; Zhang, Z.; Rosenberg, P.; Mirotsou, M.; Dzau, V.J. MicroRNA-Mediated In Vitro and In Vivo Direct Reprogramming of Cardiac Fibroblasts to Cardiomyocytes. Circ. Res. 2012, 110, 1465–1473. [Google Scholar] [CrossRef] [PubMed]
- Lalit, P.A.; Salick, M.R.; Nelson, D.O.; Squirrell, J.M.; Shafer, C.M.; Patel, N.G.; Saeed, I.; Schmuck, E.G.; Markandeya, Y.S.; Wong, R.; et al. Lineage Reprogramming of Fibroblasts into Proliferative Induced Cardiac Progenitor Cells by Defined Factors. Cell Stem Cell 2016, 18, 354–367. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Han, P.; Yang, H.; Ouyang, K.; Lee, D.; Lin, Y.-F.; Ocorr, K.; Kang, G.; Chen, J.; Stainier, D.Y.R.; et al. In vivo cardiac reprogramming contributes to zebrafish heart regeneration. Nature 2013, 498, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Giacca, M.; Zacchigna, S. Harnessing the microRNA pathway for cardiac regeneration. J. Mol. Cell. Cardiol. 2015, 89, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Zacchigna, S.; Giacca, M. Extra- and intracellular factors regulating cardiomyocyte proliferation in postnatal life. Cardiovasc. Res. 2014, 102, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Zacchigna, S.; Martinelli, V.; Moimas, S.; Colliva, A.; Anzini, M.; Nordio, A.; Costa, A.; Rehman, M.; Vodret, S.; Pierro, C.; et al. Paracrine effect of regulatory T cells promotes cardiomyocyte proliferation during pregnancy and after myocardial infarction. Nat. Commun. 2018, 9, 2432. [Google Scholar] [CrossRef] [PubMed]
- Scuderi, G.J.; Butcher, J. Naturally Engineered Maturation of Cardiomyocytes. Front. Cell Dev. Biol. 2017, 5, 50. [Google Scholar] [CrossRef] [PubMed]
- McCulley, D.J.; Black, B.L. Transcription factor pathways and congenital heart disease. Curr. Top. Dev. Biol. 2012, 100, 253–277. [Google Scholar] [CrossRef] [PubMed]
- He, A.; Gu, F.; Hu, Y.; Ma, Q.; Ye, L.Y.; Akiyama, J.A.; Visel, A.; Pennacchio, L.A.; Pu, W.T. Dynamic GATA4 enhancers shape the chromatin landscape central to heart development and disease. Nat. Commun. 2014, 5, 4907. [Google Scholar] [CrossRef] [PubMed]
- Bai, F.; Ho Lim, C.; Jia, J.; Santostefano, K.; Simmons, C.; Kasahara, H.; Wu, W.; Terada, N.; Jin, S. Directed Differentiation of Embryonic Stem Cells into Cardiomyocytes by Bacterial Injection of Defined Transcription Factors. Sci. Rep. 2015, 5, 15014. [Google Scholar] [CrossRef] [PubMed]
- Kolanowski, T.J.; Antos, C.L.; Guan, K. Making human cardiomyocytes up to date: Derivation, maturation state and perspectives. Int. J. Cardiol. 2017, 241, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, D. Making or Breaking the Heart: From Lineage Determination to Morphogenesis. Cell 2006, 126, 1037–1048. [Google Scholar] [CrossRef] [PubMed]
- Robertson, C.; Tran, D.D.; George, S.C. Concise review: Maturation phases of human pluripotent stem cell-derived cardiomyocytes. Stem Cells 2013, 31, 829–837. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Pabon, L.; Murry, C.E. Engineering Adolescence: Maturation of Human Pluripotent Stem Cell-Derived Cardiomyocytes. Circ. Res. 2014, 114, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, O.; Zdunek, S.; Alkass, K.; Druid, H.; Bernard, S.; Frisén, J. Identification of cardiomyocyte nuclei and assessment of ploidy for the analysis of cell turnover. Exp. Cell Res. 2011, 317, 188–194. [Google Scholar] [CrossRef] [PubMed]
- McCain, M.L.; Parker, K.K. Mechanotransduction: The role of mechanical stress, myocyte shape, and cytoskeletal architecture on cardiac function. Pflüg. Arch. Eur. J. Physiol. 2011, 462, 89–104. [Google Scholar] [CrossRef] [PubMed]
- Manfra, O.; Frisk, M.; Louch, W.E. Regulation of Cardiomyocyte T-Tubular Structure: Opportunities for Therapy. Curr. Heart Fail. Rep. 2017, 14, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Scharnhorst, K.S.; Stieg, A.Z.; Gimzewski, J.K.; Minami, I.; Nakatsuji, N.; Nakano, H.; Nakano, A. Two dimensional electrophysiological characterization of human pluripotent stem cell-derived cardiomyocyte system. Sci. Rep. 2017, 7, 43210. [Google Scholar] [CrossRef] [PubMed]
- Angst, B.D.; Khan, L.U.; Severs, N.J.; Whitely, K.; Rothery, S.; Thompson, R.P.; Magee, A.I.; Gourdie, R.G. Dissociated spatial patterning of gap junctions and cell adhesion junctions during postnatal differentiation of ventricular myocardium. Circ. Res. 1997, 80, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Xi, Y.; Zheng, Y.; Wang, X.; Cooney, A.J. Generation of electrophysiologically functional cardiomyocytes from mouse induced pluripotent stem cells. Stem Cell Res. 2016, 16, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Piquereau, J.; Caffin, F.; Novotova, M.; Lemaire, C.; Veksler, V.; Garnier, A.; Ventura-Clapier, R.; Joubert, F. Mitochondrial dynamics in the adult cardiomyocytes: Which roles for a highly specialized cell? Front. Physiol. 2013, 4, 102. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, L.; Liu, Z.; Alimohamadi, S.; Yin, C.; Liu, J.; Qian, L. Comparative Gene Expression Analyses Reveal Distinct Molecular Signatures between Differentially Reprogrammed Cardiomyocytes. Cell Rep. 2017, 20, 3014–3024. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, V.; Rezende, N.C.; Scotland, K.B.; Shaffer, S.M.; Persson, J.L.; Gudas, L.J.; Mongan, N.P. Regulation of Stem Cell Pluripotency and Differentiation Involves a Mutual Regulatory Circuit of the Nanog, OCT4, and SOX2 Pluripotency Transcription Factors with Polycomb Repressive Complexes and Stem Cell microRNAs. Stem Cells Dev. 2009, 18, 1093–1108. [Google Scholar] [CrossRef] [PubMed]
- Gattazzo, F.; Urciuolo, A.; Bonaldo, P. Extracellular matrix: A dynamic microenvironment for stem cell niche. Biochim. Biophys. Acta Gen. Subj. 2014, 1840, 2506–2519. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.S.; Fitzgerald, W.; Zimmerberg, J.; Kleinman, H.K.; Margolis, L. Cell-Cell and Cell-Extracellular Matrix Interactions Regulate Embryonic Stem Cell Differentiation. Stem Cells 2007, 25, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix Elasticity Directs Stem Cell Lineage Specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-B.; Dembo, M.; Wang, Y.-L. Substrate flexibility regulates growth and apoptosis of normal but not transformed cells. Am. J. Physiol. Physiol. 2000, 279, C1345–C1350. [Google Scholar] [CrossRef] [PubMed]
- Miskon, A.; Mahara, A.; Uyama, H.; Yamaoka, T. A suspension induction for myocardial differentiation of rat mesenchymal stem cells on various extracellular matrix proteins. Tissue Eng. Part C Methods 2010, 16, 979–987. [Google Scholar] [CrossRef] [PubMed]
- Lockhart, M.; Wirrig, E.; Phelps, A.; Wessels, A. Extracellular matrix and heart development. Birth Defects Res. Part A Clin. Mol. Teratol. 2011, 91, 535–550. [Google Scholar] [CrossRef] [PubMed]
- Xi, J.; Khalil, M.; Shishechian, N.; Hannes, T.; Pfannkuche, K.; Liang, H.; Fatima, A.; Haustein, M.; Suhr, F.; Bloch, W.; et al. Comparison of contractile behavior of native murine ventricular tissue and cardiomyocytes derived from embryonic or induced pluripotent stem cells. FASEB J. 2010, 24, 2739–2751. [Google Scholar] [CrossRef] [PubMed]
- Marinkovic, M.; Block, T.J.; Rakian, R.; Li, Q.; Wang, E.; Reilly, M.A.; Dean, D.D.; Chen, X.-D. One size does not fit all: Developing a cell-specific niche for in vitro study of cell behavior. Matrix Biol. 2016, 52–54, 426–441. [Google Scholar] [CrossRef] [PubMed]
- Martino, S.; D’Angelo, F.; Armentano, I.; Kenny, J.M.; Orlacchio, A. Stem cell-biomaterial interactions for regenerative medicine. Biotechnol. Adv. 2012, 30, 338–351. [Google Scholar] [CrossRef] [PubMed]
- Ikonen, L. Analysis of Different Natural and Synthetic Biomaterials to Support Cardiomyocyte Growth. J. Clin. Exp. Cardiol. 2013, 04. [Google Scholar] [CrossRef]
- Morez, C.; Noseda, M.; Abreu, M.; Belian, E.; Schneider, M.D.; Stevens, M.M. Biomaterials Enhanced ef fi ciency of genetic programming toward cardiomyocyte creation through topographical cues. Biomaterials 2015, 70, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Engler, A.J.; Carag-Krieger, C.; Johnson, C.P.; Raab, M.; Tang, H.-Y.; Speicher, D.W.; Sanger, J.W.; Sanger, J.M.; Discher, D.E. Embryonic cardiomyocytes beat best on a matrix with heart-like elasticity: Scar-like rigidity inhibits beating. J. Cell Sci. 2008, 121, 3794–3802. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Xiao, Z.; Ren, X.; Long, H.; Ma, K.; Qian, H.; Guo, Y. Obtaining spontaneously beating cardiomyocyte-like cells from adipose-derived stromal vascular fractions cultured on enzyme-crosslinked gelatin hydrogels. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Cutts, J.; Nikkhah, M.; Brafman, D.A. Biomaterial Approaches for Stem Cell-Based Myocardial Tissue Engineering. Biomark. Insights 2015, 10, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-W.; Lee, H.J.; Hwang, H.S.; Ko, K.; Han, D.W.; Ko, K. Optimization of Matrigel-based culture for expansion of neural stem cells. Anim. Cells Syst. 2015, 19, 175–180. [Google Scholar] [CrossRef]
- Hinderer, S.; Layland, S.L.; Schenke-Layland, K. ECM and ECM-like materials—Biomaterials for applications in regenerative medicine and cancer therapy. Adv. Drug Deliv. Rev. 2016, 97, 260–269. [Google Scholar] [CrossRef] [PubMed]
- Moroni, F.; Mirabella, T. Decellularized matrices for cardiovascular tissue engineering. Am. J. Stem Cells 2014, 3, 1–20. [Google Scholar] [PubMed]
- Chachques, J.C.; Pradas, M.M.; Bayes-Genis, A.; Semino, C. Creating the bioartificial myocardium for cardiac repair: Challenges and clinical targets. Expert Rev. Cardiovasc. Ther. 2013, 11, 1701–1711. [Google Scholar] [CrossRef] [PubMed]
- Lakshmanan, R.; Krishnan, U.M.; Sethuraman, S. Polymeric Scaffold Aided Stem Cell Therapeutics for Cardiac Muscle Repair and Regeneration. Macromol. Biosci. 2013, 13, 1119–1134. [Google Scholar] [CrossRef] [PubMed]
- Sartori, S.; Chiono, V.; Tonda-Turo, C.; Mattu, C.; Gianluca, C. Biomimetic polyurethanes in nano and regenerative medicine. J. Mater. Chem. B 2014, 2, 5128–5144. [Google Scholar] [CrossRef]
- Chiono, V.; Mozetic, P.; Boffito, M.; Sartori, S.; Gioffredi, E.; Silvestri, A.; Rainer, A.; Giannitelli, S.M.; Trombetta, M.; Nurzynska, D.; et al. Polyurethane-based scaffolds for myocardial tissue engineering. Interface Focus 2013, 4, 20130045. [Google Scholar] [CrossRef] [PubMed]
- Carmagnola, I.; Ranzato, E.; Chiono, V. Scaffold functionalization to support a tissue biocompatibility. In Functional 3D Tissue Engineering Scaffolds; Elsevier/Woodhead Publishing: Amsterdam, The Netherlands, 2018; pp. 255–277, Chapter 11; ISBN 9780081009796. [Google Scholar]
- Chiono, V.; Nardo, T.; Ciardelli, G. Bioartificial materials for regenerative medicine applications. In Regenerative Medicine Applications in Organ Transplantation; Elsevier/AP: Amsterdam, The Netherlands, 2014; pp. 113–136, Chapter 9; ISBN 9780123985231. [Google Scholar]
- Boffito, M.; Di Meglio, F.; Mozetic, P.; Giannitelli, S.M.; Carmagnola, I.; Castaldo, C.; Nurzynska, D.; Sacco, A.M.; Miraglia, R.; Montagnani, S.; et al. Surface functionalization of polyurethane scaffolds mimicking cardiac microenvironment to support cardiac primitive cells. PLoS ONE 2018, 13, e0199896. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Shadrin, I.Y.; Lam, J.; Xian, H.-Q.; Snodgrass, H.R.; Bursac, N. Tissue-engineered cardiac patch for advanced functional maturation of human ESC-derived cardiomyocytes. Biomaterials 2013, 34, 5813–5820. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, S.; Lin, Q.; Wang, J.; Lim, T.K.; Joshi, S.B.; Anand, G.S.; Chung, M.C.M.; Sheetz, M.P.; Fujita, H. Heart extracellular matrix supports cardiomyocyte differentiation of mouse embryonic stem cells. J. Biosci. Bioeng. 2013, 115, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Prabhakaran, M.P.; Mobarakeh, L.G.; Kai, D.; Karbalaie, K.; Nasr-Esfahani, M.H.; Ramakrishna, S. Differentiation of embryonic stem cells to cardiomyocytes on electrospun nanofibrous substrates. J. Biomed. Mater. Res. Part B Appl. Biomater. 2014, 102, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Sa, S.; Wong, L.; McCloskey, K.E. Combinatorial Fibronectin and Laminin Signaling Promote Highly Efficient Cardiac Differentiation of Human Embryonic Stem Cells. BioRes. Open Access 2014, 3, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zeng, D.; Ding, L.; Li, X.L.; Liu, X.T.; Li, W.J.; Wei, T.; Yan, S.; Xie, J.H.; Wei, L.; et al. Three-dimensional poly-(ε-caprolactone) nanofibrous scaffolds directly promote the cardiomyocyte differentiation of murine-induced pluripotent stem cells through Wnt/β-catenin signaling. BMC Cell Biol. 2015, 16, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Fong, A.H.; Romero-López, M.; Heylman, C.M.; Keating, M.; Tran, D.; Sobrino, A.; Tran, A.Q.; Pham, H.H.; Fimbres, C.; Gershon, P.D.; et al. Three-Dimensional Adult Cardiac Extracellular Matrix Promotes Maturation of Human Induced Pluripotent Stem Cell-Derived Cardiomyocytes. Tissue Eng. Part A 2016, 22, 1016–1025. [Google Scholar] [CrossRef] [PubMed]
- Hirata, M.; Yamaoka, T. Effect of stem cell niche elasticity/ECM protein on the self-beating cardiomyocyte differentiation of induced pluripotent stem (iPS) cells at different stages. Acta Biomater. 2018, 65, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Tan, G.; Shim, W.; Gu, Y.; Qian, L.; Ying Chung, Y.; Yun Lim, S.; Yong, P.; Sim, E.; Wong, P. Differential effect of myocardial matrix and integrins on cardiac differentiation of human mesenchymal stem cells. Differentiation 2010, 79, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Guan, J.; Wang, F.; Li, Z.; Chen, J.; Guo, X.; Liao, J.; Moldovan, N.I. The stimulation of the cardiac differentiation of mesenchymal stem cells in tissue constructs that mimic myocardium structure and biomechanics. Biomaterials 2011, 32, 5568–5580. [Google Scholar] [CrossRef] [PubMed]
- Santhakumar, R.; Vidyasekar, P.; Verma, R.S. Cardiogel: A Nano-Matrix Scaffold with Potential Application in Cardiac Regeneration Using Mesenchymal Stem Cells. PLoS ONE 2014, 9, e114697. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Park, S.; Ryu, S.; Bhang, S.H.; Kim, J.; Yoon, J.-K.; Park, Y.H.; Cho, S.-P.; Lee, S.; Hong, B.H.; et al. Graphene-Regulated Cardiomyogenic Differentiation Process of Mesenchymal Stem Cells by Enhancing the Expression of Extracellular Matrix Proteins and Cell Signaling Molecules. Adv. Healthc. Mater. 2014, 3, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.P.; Bache-Wiig, M.K.; Provenzano, P.P.; Ogle, B.M. Heterogeneous Differentiation of Human Mesenchymal Stem Cells in 3D Extracellular Matrix Composites. BioRes. Open Access 2016, 5, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Rashedi, I.; Talele, N.; Wang, X.-H.; Hinz, B.; Radisic, M.; Keating, A. Collagen scaffold enhances the regenerative properties of mesenchymal stromal cells. PLoS ONE 2017, 12, e0187348. [Google Scholar] [CrossRef] [PubMed]
- French, K.M.; Boopathy, A.V.; DeQuach, J.A.; Chingozha, L.; Lu, H.; Christman, K.L.; Davis, M.E. A naturally derived cardiac extracellular matrix enhances cardiac progenitor cell behavior in vitro. Acta Biomater. 2012, 8, 4357–4364. [Google Scholar] [CrossRef] [PubMed]
- Castaldo, C.; Di Meglio, F.; Miraglia, R.; Sacco, A.M.; Romano, V.; Bancone, C.; Della Corte, A.; Montagnani, S.; Nurzynska, D. Cardiac Fibroblast-Derived Extracellular Matrix (Biomatrix) as a Model for the Studies of Cardiac Primitive Cell Biological Properties in Normal and Pathological Adult Human Heart. BioMed Res. Int. 2013, 2013, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Gaetani, R.; Yin, C.; Srikumar, N.; Braden, R.; Doevendans, P.A.; Sluijter, J.P.G.; Christman, K.L. Cardiac-Derived Extracellular Matrix Enhances Cardiogenic Properties of Human Cardiac Progenitor Cells. Cell Transplant. 2016, 25, 1653–1663. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.P.; Carrion, B.; Singh, R.K.; Putnam, A.J. Matrix identity and tractional forces influence indirect cardiac reprogramming. Sci. Rep. 2013, 3, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.W.; Hoyne, J.D.; Nguyen, P.K.; McCreedy, D.A.; Aly, H.; Efimov, I.R.; Rentschler, S.; Elbert, D.L. Direct reprogramming of mouse fibroblasts to cardiomyocyte-like cells using Yamanaka factors on engineered poly(ethylene glycol) (PEG) hydrogels. Biomaterials 2013, 34, 6559–6571. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Dal-Pra, S.; Mirotsou, M.; Jayawardena, T.M.; Hodgkinson, C.P.; Bursac, N.; Dzau, V.J. Tissue-engineered 3-dimensional (3D) microenvironment enhances the direct reprogramming of fibroblasts into cardiomyocytes by microRNAs. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Vanwinkle, W.B.; Snuggs, M.B.; Buja, L.M. Cardiogel: A biosynthetic extracellular matrix for cardiomyocyte culture. In Vitro Cell. Dev. Biol. Anim. 1996, 32, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Yuan, T.; Li, K.; Guo, L.; Fan, H.; Zhang, X. Modulation of immunological properties of allogeneic mesenchymal stem cells by collagen scaffolds in cartilage tissue engineering. J. Biomed. Mater. Res. A 2011, 98, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Huang, Y.; Spencer, C.I.; Foley, A.; Vedantham, V.; Liu, L.; Conway, S.J.; Fu, J.D.; Srivastava, D. In vivo reprogramming of murine cardiac fibroblasts into induced cardiomyocytes. Nature 2012, 485, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Schuldiner, M.; Yanuka, O.; Itskovitz-Eldor, J.; Melton, D.A.; Benvenisty, N. Effects of eight growth factors on the differentiation of cells derived from human embryonic stem cells. Proc. Natl. Acad. Sci. USA 2000, 97, 11307–11312. [Google Scholar] [CrossRef] [PubMed]
- Monaghan, M.G.; Holeiter, M.; Layland, S.L.; Schenke-Layland, K. Cardiomyocyte generation from somatic sources—Current status and future directions. Curr. Opin. Biotechnol. 2016, 40, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Miner, J.H.; Yurchenco, P.D. Laminin Functions In Tissue Morphogenesis. Annu. Rev. Cell Dev. Biol. 2004, 20, 255–284. [Google Scholar] [CrossRef] [PubMed]
- Castaldo, C.; Di Meglio, F.; Nurzynska, D.; Romano, G.; Maiello, C.; Bancone, C.; Müller, P.; Böhm, M.; Cotrufo, M.; Montagnani, S. CD117-Positive Cells in Adult Human Heart Are Localized in the Subepicardium, and Their Activation Is Associated with Laminin-1 and α6 Integrin Expression. Stem Cells 2008, 26, 1723–1731. [Google Scholar] [CrossRef] [PubMed]
- Stoppel, W.L.; Kaplan, D.L.; Black, L.D. Electrical and mechanical stimulation of cardiac cells and tissue constructs. Adv. Drug Deliv. Rev. 2016, 96, 135–155. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
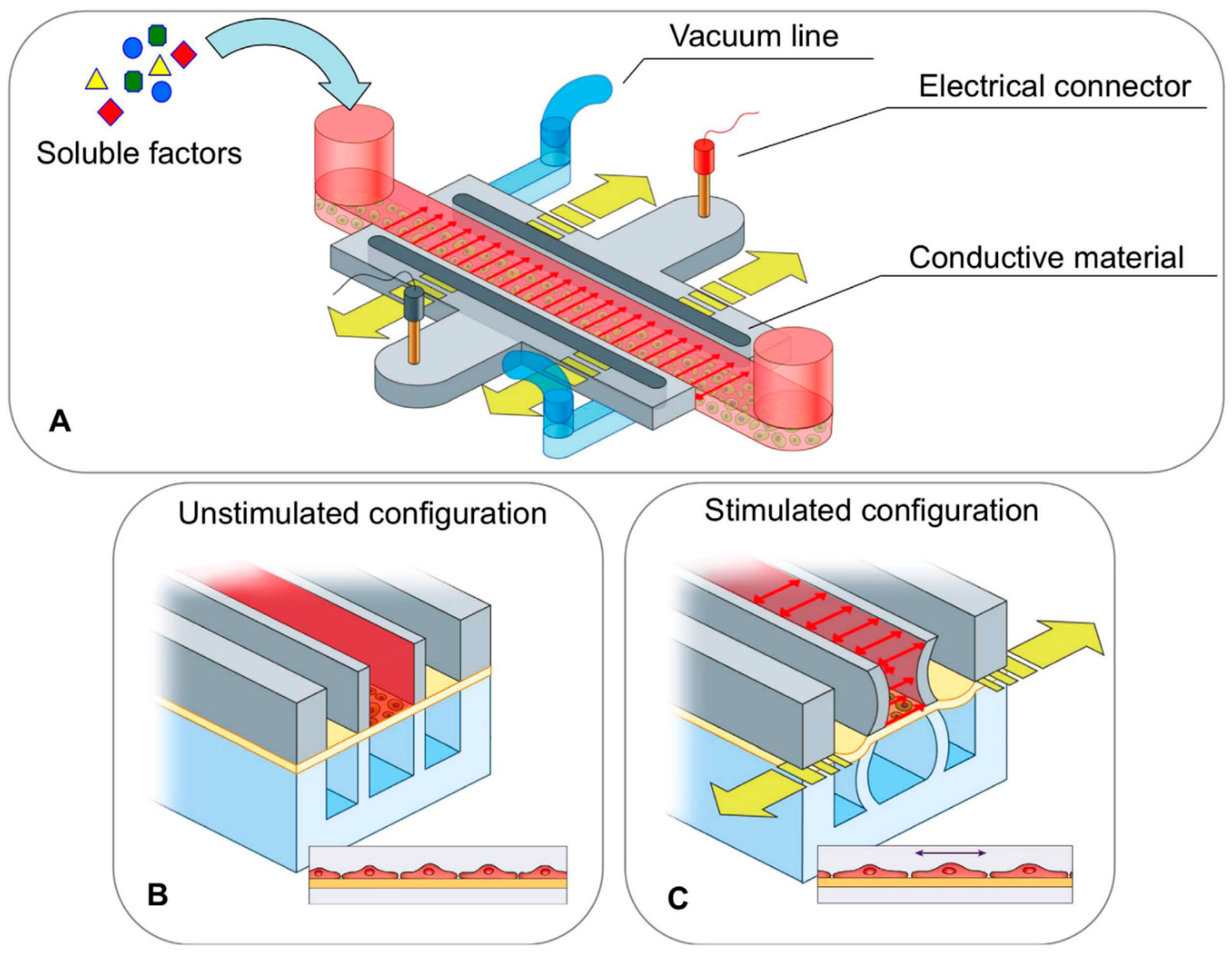{kind=link}
| Cell Types | Substrate Type | Main Results | References |
|---|---|---|---|
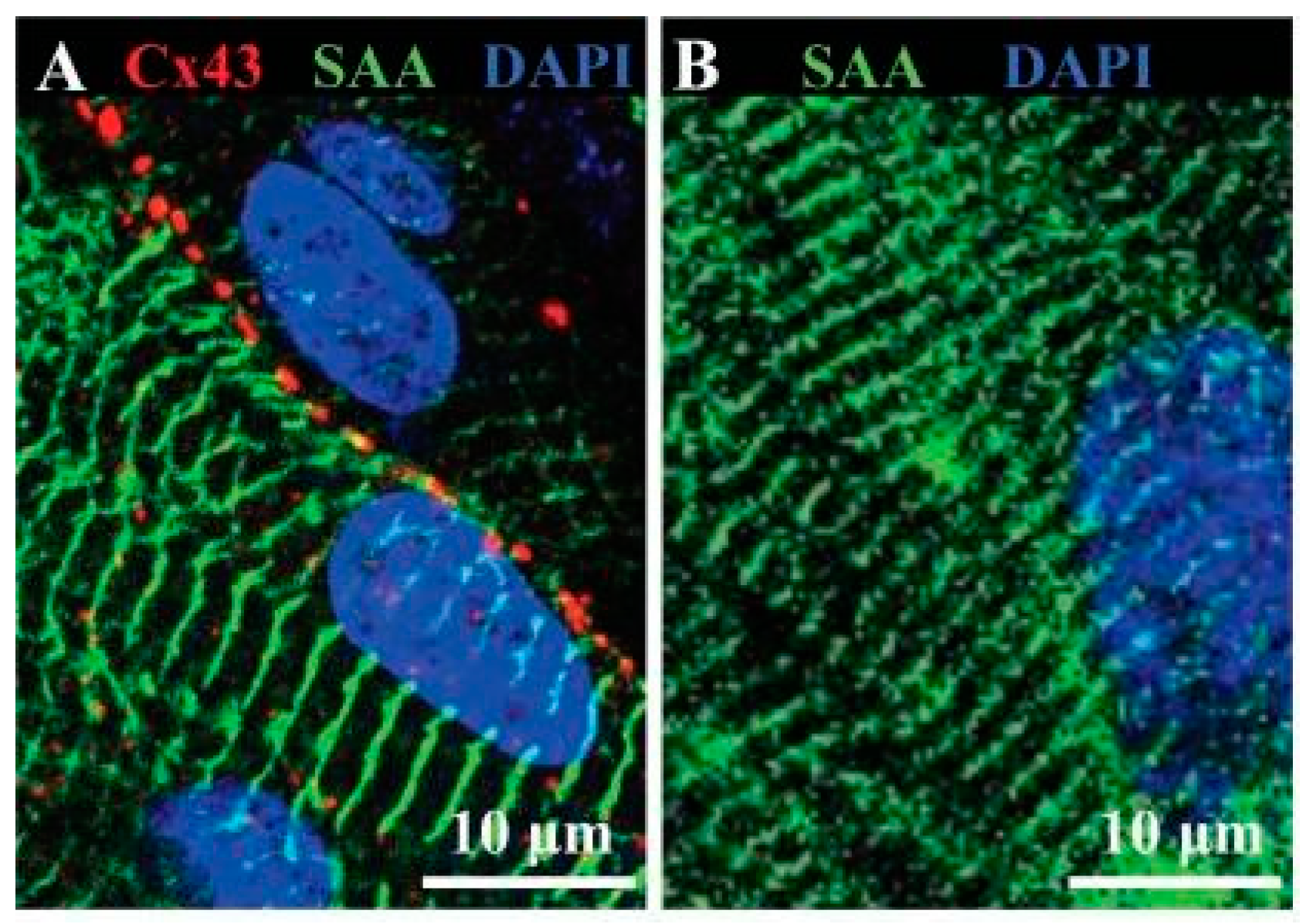
| hESC | 3D fibrin/Matrigel hydrogel cardiac tissue patch vs. 2D fibrin/Matrigel culture | Longer sarcomere length, increased action potential conduction velocity, expression of cTnT, α-MHC, SERCA2, CASQ2 and Cx43 | Zhang et al., 2013 [92] |
| mESCs | Mouse decellularized heart ECM vs. liver ECM | Increased expression of cTnI, α-MHC and MCL3 | Higuchi et al., 2013 [93] |
| mESC | Collagen Type I blended PLGA electrospun scaffold vs. PLGA and TCP | Acquisition of a spindle-like cardiomyocyte morphology, expression of α actinin and Cx43 | Prabhakaran et al., 2014 [94] |
| hESC | Fibronectin (70%) and laminin (30%) combination substrate | Generation of 60% of cTnI and Nkx2.5 positive cells | Sa et al., 2014 [95] |
| miPSC | 3D gelatin-coated electrospun PCL scaffold vs. TCP | Increased expression of MESP1, Gata-4, Nkx2.5 and cTnT | Chen et al., 2015 [96] |
| hiPSC | 2D vs. 3D bovine decellularized cardiac foetal and adult ECM | 3D adult ECM enhanced genes related to calcium handling (JNC), inward rectifier potassium ion channels (KCNJ2/Kir2.1), CaV1.2, Cx43 and Myl2 | Fong et al., 2016 [97] |
| miPSCs | Polyacrylamide gels with different elastic moduli and functionalized with collagen Type I, fibronectin and gelatin vs. TCP | Expression of Gata-4, Mef2c and Tbx5 increased on gelatin/fibronectin-TCP; α-MHC, cTnT and cTnI increased on Es20 collagen gel. | Hirata et al., 2018 [98] |
| hBMSCs | Collagen Type V matrix | Expression of both early cardiac genes, such as Gata-4 and Nkx2.5 and their downstream genes, such as α skeletal actinin, cTnT and cTnC. | Tan et al., 2010 [99] |
| hMSCs | 3D tissue PECUU fibrous construct vs. TCP | Cardiac differentiation with increased expression of Mef2c, Nkx2.5 and Gata-4 | Guan et al., 2011 [100] |
| mMSCs | Cardiogel (fibroblast-derived ECM enriched in collagen Type I and III, laminin and fibronectin) | Approximately 15–20% of mouse BMSCs cultured for 4-weeks on Cardiogel developed three-dimensional myotubule-like multinucleated structure. Cardiac cell commitment was further confirmed by the expression of Gata-4, α-sarcomeric actinin and Cx43. | Santhakumar et al., 2014 [101] |
| hMSCs | Graphene substrate | Enhanced Gata-4, β-MHC, cTnT, Mlc2a, Mlc2v and Cx43 expression. | Park et al., 2014 [102] |
| hMSCs | 3D collagen Type I substrate | Enhanced cardiogenic gene expression compared to TCP coated with collagen Type I or laminin, which induced cardiogenic, osteogenic and adipogenic gene expression | Jung et al., 2016 [103] |
| hMSCs | Collagen Type I 3D patch | Increased expression of α-MHC and cTnT | Rashedi et al., 2017 [104] |
| rCPCs | Porcine-derived cardiac decellularized ECM vs. collagen matrix | Enhanced proliferation, adhesion and apoptosis reduction. Increased expression of Nkx2.5, Gata-4, cTnT and α-MHC | French et al., 2012 [105] |
| hCPCs | Biomatrix obtained from adult human cardiac fibroblasts isolated from healthy and pathological heart tissues. | Biomatrix stimulated migration and protected cells from apoptotic processes. | Castaldo et al., 2013 [106] |
| hCPCs | Polyurethane-based scaffold | Scaffold supported in vitro CPC adhesion and viability; it did not support cell proliferation. | Chiono et al., 2013 [88] |
| hCPCs | 3D collagen-based vs. porcine cardiac ECM-based hydrogel. | Cardiac ECM hydrogel enhanced Gata-4, Mlc2v, Vegfr2 expression in foetal CPCs and Nkx2.5, Mef2c, CD31, Vegfr2 expression in adult CPCs. | Gaetani et al., 2016 [107] |
| hCPC | 3D PU-based scaffold with laminin-1 surface functionalization | Increased CPC proliferation, decreased CPC apoptosis. Expression of cardiac markers (Mef2c and α sarcomeric actinin), smooth muscle cell markers (Gata-6 and SMA) and endothelial cell markers (ETS1 and FVIII) | Boffito et al., 2018 [91] |
| Mouse embryonic fibroblasts (MEFs) | Indirect reprogramming through different ECM protein based-hydrogels (Matrigel, collagen Type I and fibrin) | Fibrin hydrogel supported both dedifferentiation and differentiation phase; collagen Type I /fibrin gel increased the percentage of contractile colonies. | Kong et al., 2013 [108] |
| MEFs | Direct reprogramming using PEG hydrogels functionalized with laminin and RGD at different concentration vs. TCP | High concentration of laminin and RGD supported MEF reprogramming | Smith et al., 2013 [109] |
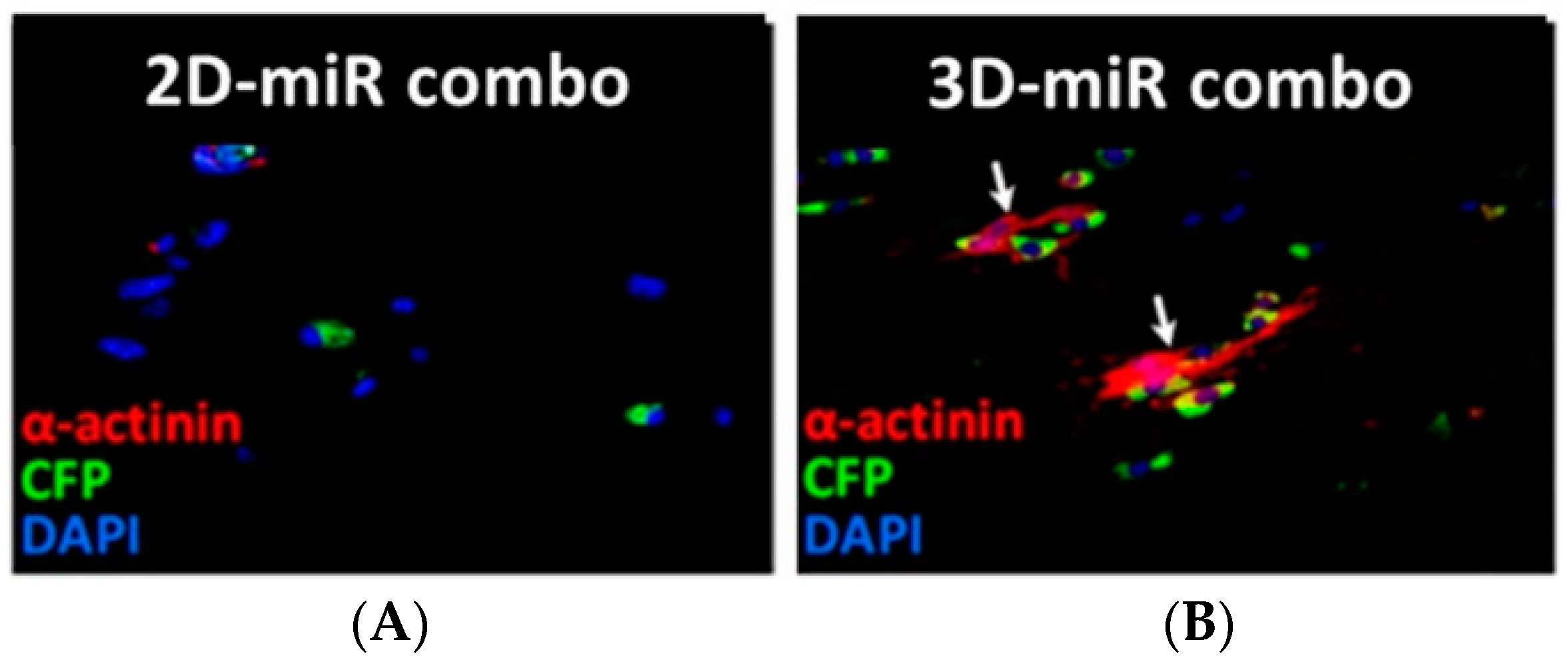
| MEFs, Tail tip fibroblasts (TTFs) | miRNA mediated-direct reprogramming using fibrin based-3D hydrogels vs. 2D culture | Fibrin based-3D hydrogels supported MEF reprogramming enhancing α-MHC, cTnI, α-sarcomeric actinin and Kcnj2 expression; increased Mef2c, Tbx5 and Hand2 expression in TTFs. | Li et al., 2016 [110] |
| Cells | Biomaterials for CM Generation |
|---|---|
| Stem cell differentiation | |
| Pluripotent stem cells: | |
| ESCs | Decellularized cardiac ECM [93], fibrin /Matrigel hydrogel [92], collagen Type I [94]—fibronectin/laminin 70/30 hydrogel [95] |
| iPSCs | Decellularized cardiac ECM [97], gelatin [96]—and collagen Type I [98]-coated scaffolds/hydrogels |
| Multipotent stem cells: | |
| MSCs | Cardiogel [101], collagen Type V substrates [99] |
| CPCs | Decellularized cardiac ECM [105], biomatrix [106], laminin-1 coated scaffolds [91] |
| Indirect Reprogramming | |
| Fibroblasts | Collagen Type I/Fibrin and Fibrin hydrogels [108] |
| Direct Reprogramming | |
| Fibroblasts | Fibrin/Matrigel hydrogel [110], PEG hydrogel containing RGD and laminin [109] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paoletti, C.; Divieto, C.; Chiono, V. Impact of Biomaterials on Differentiation and Reprogramming Approaches for the Generation of Functional Cardiomyocytes. Cells 2018, 7, 114. https://doi.org/10.3390/cells7090114
Paoletti C, Divieto C, Chiono V. Impact of Biomaterials on Differentiation and Reprogramming Approaches for the Generation of Functional Cardiomyocytes. Cells. 2018; 7(9):114. https://doi.org/10.3390/cells7090114
Chicago/Turabian StylePaoletti, Camilla, Carla Divieto, and Valeria Chiono. 2018. "Impact of Biomaterials on Differentiation and Reprogramming Approaches for the Generation of Functional Cardiomyocytes" Cells 7, no. 9: 114. https://doi.org/10.3390/cells7090114
APA StylePaoletti, C., Divieto, C., & Chiono, V. (2018). Impact of Biomaterials on Differentiation and Reprogramming Approaches for the Generation of Functional Cardiomyocytes. Cells, 7(9), 114. https://doi.org/10.3390/cells7090114