
Inhibition of Toll-like Receptor 4 Using Small Molecule, TAK-242, Protects Islets from Innate Immune Responses

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Sample Collection
2.2. Islet Isolation
2.3. Surface Modification of Islets
2.4. In Vitro IBMIR
2.5. Clotting Assay
2.6. One-Way Mixed Lymphocyte Reaction
2.7. CD8+ T Cell Activation Assay
2.8. In Vitro Co-Culture of Allogenic PBMC and Islets
2.9. Cytokine Quantification
2.10. RT-qPCR for Secreted Stress and Damage miRNA
2.11. Flow Cytometry
2.12. Statistical Analysis
3. Results
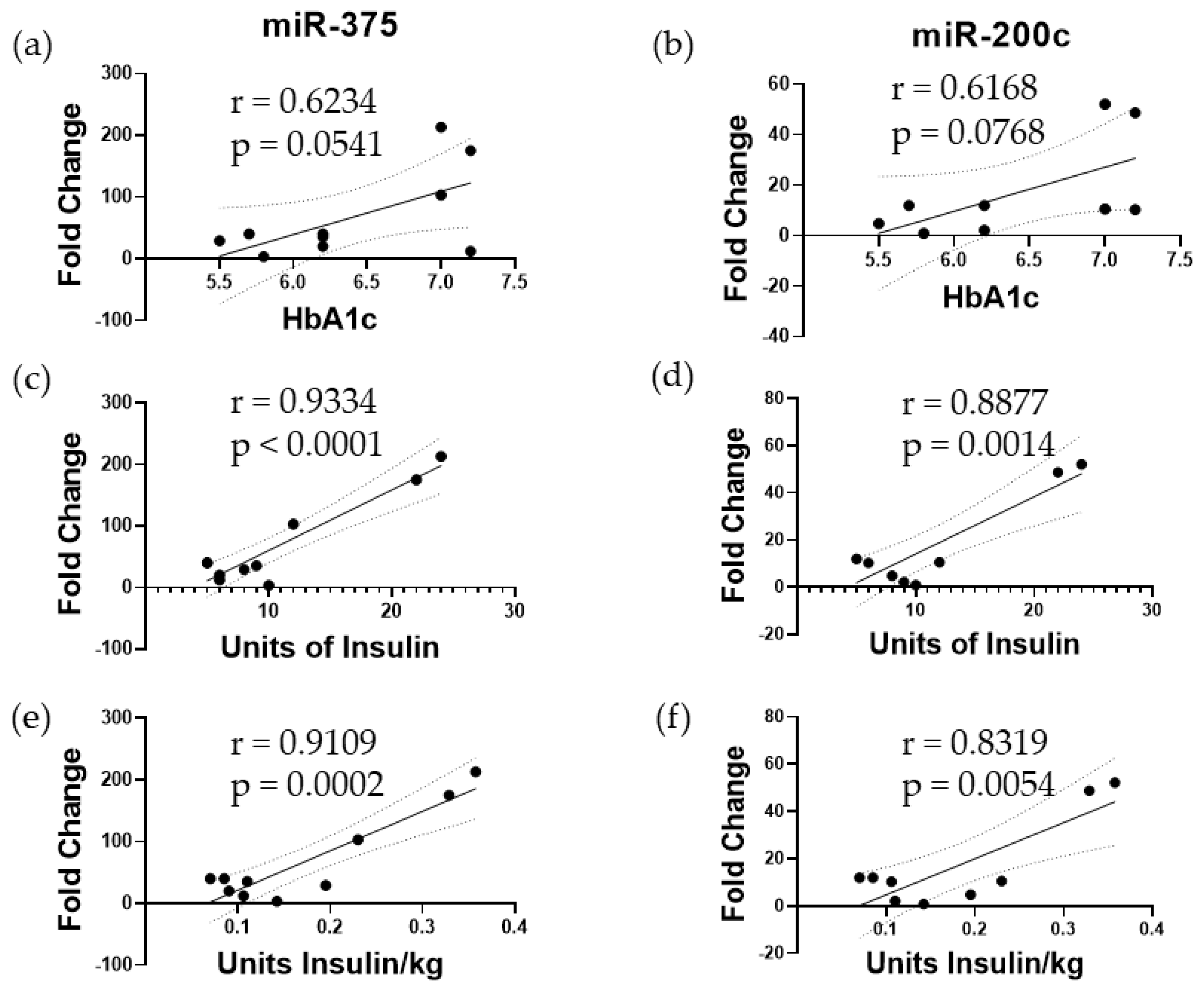
3.1. Elevated miR-375 and miR-200c Correspond to Poor Islet Function following TPIAT
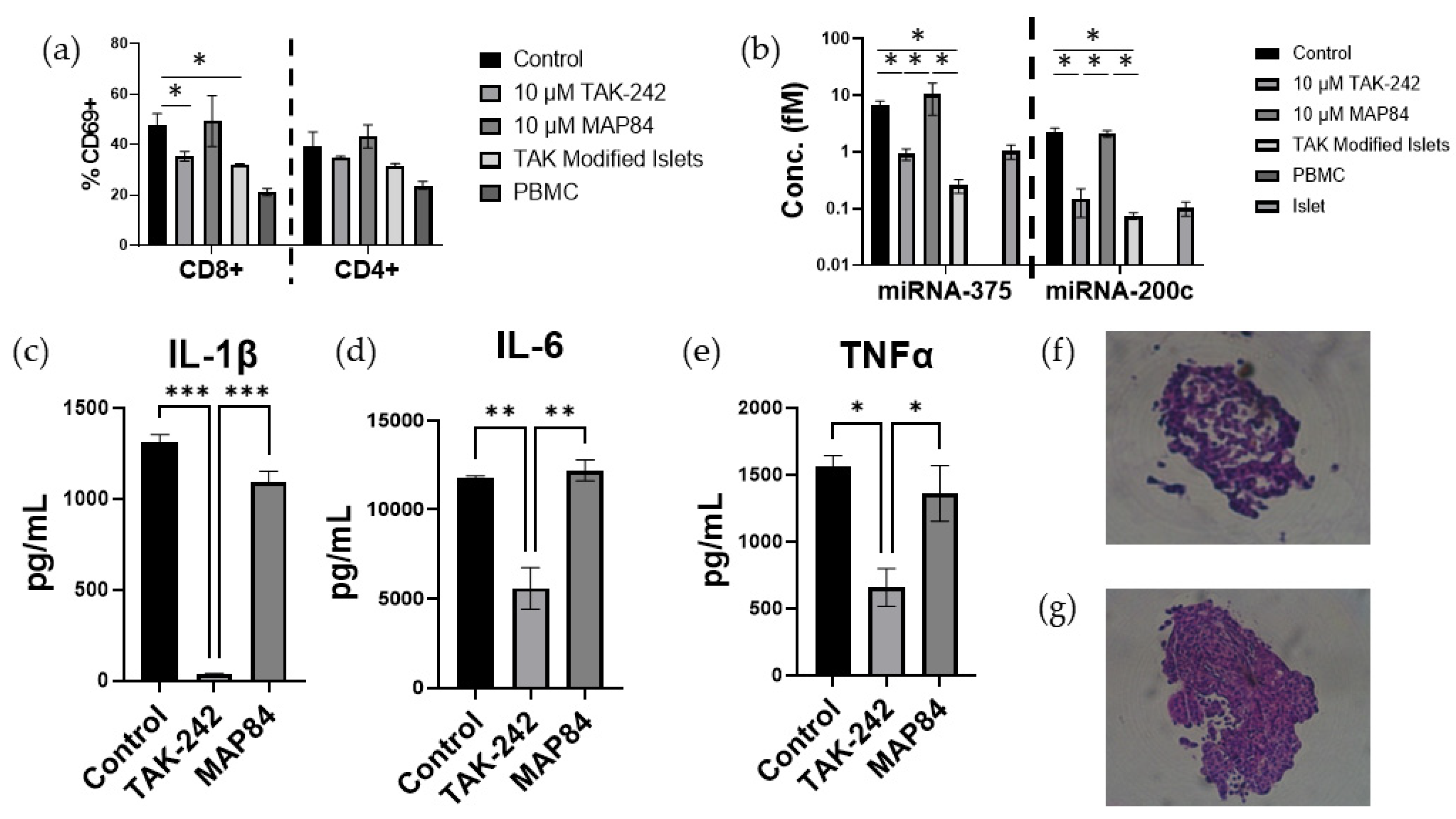
3.2. TAK-242 Reduces Damage to Islets Exposed to IBMIR In Vitro
3.3. TAK-242 Inhibits T Cell Activation and Proliferation in a One-Way Mixed Lymphocyte Reaction with Splenocytes
3.4. TAK-242 Directly Inhibits the Activation of CD8+ T Cells
3.5. T Cell Activation and Islet Damage Is Inhibited by the Presence of TAK-242
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Katsarou, A.; Gudbjörnsdottir, S.; Rawshani, A.; Dabelea, D.; Bonifacio, E.; Anderson, B.J.; Jacobsen, L.M.; Schatz, D.A.; Lernmark, Å. Type 1 Diabetes Mellitus. Nat. Rev. Dis. Primer 2017, 3, 17016. [Google Scholar] [CrossRef]
- Shapiro, A.M.J.; Pokrywczynska, M.; Ricordi, C. Clinical Pancreatic Islet Transplantation. Nat. Rev. Endocrinol. 2017, 13, 268–277. [Google Scholar] [CrossRef]
- Sutherland, D.E.R.; Radosevich, D.M.; Bellin, M.D.; Hering, B.J.; Beilman, G.J.; Dunn, T.B.; Chinnakotla, S.; Vickers, S.M.; Bland, B.; Balamurugan, A.N.; et al. Total Pancreatectomy and Islet Autotransplantation for Chronic Pancreatitis. J. Am. Coll. Surg. 2012, 214, 409–424. [Google Scholar] [CrossRef]
- Shapiro, A.M.J.; Lakey, J.R.T.; Ryan, E.A.; Korbutt, G.S.; Toth, E.; Warnock, G.L.; Kneteman, N.M.; Rajotte, R.V. Islet Transplantation in Seven Patients with Type 1 Diabetes Mellitus Using a Glucocorticoid-Free Immunosuppressive Regimen. N. Engl. J. Med. 2000, 343, 230–238. [Google Scholar] [CrossRef]
- Toso, C.; Shapiro, A.M.J.; Bowker, S.; Dinyari, P.; Paty, B.; Ryan, E.A.; Senior, P.; Johnson, J.A. Quality of Life After Islet Transplant: Impact of the Number of Islet Infusions and Metabolic Outcome. Transplantation 2007, 84, 664–666. [Google Scholar] [CrossRef]
- Foster, E.D.; Bridges, N.D.; Feurer, I.D.; Eggerman, T.L.; Hunsicker, L.G.; Alejandro, R. Clinical Islet Transplantation Consortium Improved Health-Related Quality of Life in a Phase 3 Islet Transplantation Trial in Type 1 Diabetes Complicated by Severe Hypoglycemia. Diabetes Care 2018, 41, 1001–1008. [Google Scholar] [CrossRef]
- Delaune, V.; Berney, T.; Lacotte, S.; Toso, C. Intraportal Islet Transplantation: The Impact of the Liver Microenvironment. Transpl. Int. 2017, 30, 227–238. [Google Scholar] [CrossRef]
- Bennet, W.; Groth, C.-G.; Larsson, R.; Nilsson, B.; Korsgren, O. Isolated Human Islets Trigger an Instant Blood Mediated Inflammatory Reaction: Implications for Intraportal Islet Transplantation as a Treatment for Patients with Type 1 Diabetes. Upsala J. Med Sci. 2000, 105, 125–133. [Google Scholar] [CrossRef]
- Lu, Y.-C.; Yeh, W.-C.; Ohashi, P.S. LPS/TLR4 Signal Transduction Pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef]
- Wang, N.; Liang, H.; Zen, K. Molecular Mechanisms That Influence the Macrophage M1–M2 Polarization Balance. Front. Immunol. 2014, 5, 614. [Google Scholar] [CrossRef]
- Mok, D.; Black, M.; Gupta, N.; Arefanian, H.; Tredget, E.; Rayat, G.R. Early Immune Mechanisms of Neonatal Porcine Islet Xenograft Rejection. Xenotransplantation 2019, 26, e12546. [Google Scholar] [CrossRef]
- Wu, T.-T.; Chen, T.-L.; Chen, R.-M. Lipopolysaccharide Triggers Macrophage Activation of Inflammatory Cytokine Expression, Chemotaxis, Phagocytosis, and Oxidative Ability via a Toll-like Receptor 4-Dependent Pathway: Validated by RNA Interference. Toxicol. Lett. 2009, 191, 195–202. [Google Scholar] [CrossRef]
- Dong, H.; Zhang, Y.; Song, L.; Kim, D.-S.; Wu, H.; Yang, L.; Li, S.; Morgan, K.A.; Adams, D.B.; Wang, H. Cell-Permeable Peptide Blocks TLR4 Signaling and Improves Islet Allograft Survival. Cell Transplant. 2016, 25, 1319–1329. [Google Scholar] [CrossRef]
- Shen, H.; Tesar, B.M.; Walker, W.E.; Goldstein, D.R. Dual Signaling of MyD88 and TRIF Is Critical for Maximal TLR4-Induced Dendritic Cell Maturation1. J. Immunol. 2008, 181, 1849–1858. [Google Scholar] [CrossRef]
- Alloatti, A.; Kotsias, F.; Pauwels, A.-M.; Carpier, J.-M.; Jouve, M.; Timmerman, E.; Pace, L.; Vargas, P.; Maurin, M.; Gehrmann, U.; et al. Toll-like Receptor 4 Engagement on Dendritic Cells Restrains Phago-Lysosome Fusion and Promotes Cross-Presentation of Antigens. Immunity 2015, 43, 1087–1100. [Google Scholar] [CrossRef]
- Weimershaus, M.; Mauvais, F.-X.; Saveanu, L.; Adiko, C.; Babdor, J.; Abramova, A.; Montealegre, S.; Lawand, M.; Evnouchidou, I.; Huber, K.J.; et al. Innate Immune Signals Induce Anterograde Endosome Transport Promoting MHC Class I Cross-Presentation. Cell Rep. 2018, 24, 3568–3581. [Google Scholar] [CrossRef]
- Goldberg, A.; Parolini, M.; Chin, B.Y.; Czismadia, E.; Otterbein, L.E.; Bach, F.H.; Wang, H. Toll-like Receptor 4 Suppression Leads to Islet Allograft Survival. FASEB J. 2007, 21, 2840–2848. [Google Scholar] [CrossRef]
- Giovannoni, L.; Muller, Y.D.; Lacotte, S.; Parnaud, G.; Borot, S.; Meier, R.P.H.; Lavallard, V.; Bédat, B.; Toso, C.; Daubeuf, B.; et al. Enhancement of Islet Engraftment and Achievement of Long-Term Islet Allograft Survival by Toll-like Receptor 4 Blockade. Transplantation 2015, 99, 29–35. [Google Scholar] [CrossRef]
- Krüger, B.; Yin, N.; Zhang, N.; Yadav, A.; Coward, W.; Lal, G.; Zang, W.; Heeger, P.S.; Bromberg, J.S.; Murphy, B.; et al. Islet-Expressed TLR2 and TLR4 Sense Injury and Mediate Early Graft Failure after Transplantation. Eur. J. Immunol. 2010, 40, 2914–2924. [Google Scholar] [CrossRef]
- Matsunaga, N.; Tsuchimori, N.; Matsumoto, T. Masayuki Ii TAK-242 (Resatorvid), a Small-Molecule Inhibitor of Toll-like Receptor (TLR) 4 Signaling, Binds Selectively to TLR4 and Interferes with Interactions between TLR4 and Its Adaptor Molecules. Mol. Pharmacol. 2011, 79, 34–41. [Google Scholar] [CrossRef]
- Rice, T.W.; Wheeler, A.P.; Bernard, G.R.; Vincent, J.-L.; Angus, D.C.; Aikawa, N.; Demeyer, I.; Sainati, S.; Amlot, N.; Cao, C.; et al. A Randomized, Double-Blind, Placebo-Controlled Trial of TAK-242 for the Treatment of Severe Sepsis*. Crit. Care Med. 2010, 38, 1685–1694. [Google Scholar] [CrossRef]
- Gao, Q.; Ma, L.L.; Gao, X.; Yan, W.; Williams, P.; Yin, D.P. TLR4 Mediates Early Graft Failure After Intraportal Islet Transplantation. Am. J. Transplant. 2010, 10, 1588–1596. [Google Scholar] [CrossRef]
- Chang, C.A.; Akinbobuyi, B.; Quintana, J.M.; Yoshimatsu, G.; Naziruddin, B.; Kane, R.R. Ex-Vivo Generation of Drug-Eluting Islets Improves Transplant Outcomes by Inhibiting TLR4-Mediated NFkB Upregulation. Biomaterials 2018, 159, 13–24. [Google Scholar] [CrossRef]
- Chinnakotla, S.; Beilman, G.J.; Dunn, T.B.; Bellin, M.D.; Freeman, M.L.; Radosevich, D.M.; Arain, M.; Amateau, S.K.; Mallery, J.S.; Schwarzenberg, S.J.; et al. Factors Predicting Outcomes after a Total Pancreatectomy and Islet Autotransplantation Lessons Learned From over 500 Cases. Ann. Surg. 2015, 262, 610–622. [Google Scholar] [CrossRef]
- Kin, T. Islet Isolation for Clinical Transplantation. In The Islets of Langerhans; Islam, M.S., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 683–710. ISBN 978-90-481-3271-3. [Google Scholar]
- Saravanan, P.B.; Vasu, S.; Yoshimatsu, G.; Darden, C.M.; Wang, X.; Gu, J.; Lawrence, M.C.; Naziruddin, B. Differential Expression and Release of Exosomal miRNAs by Human Islets under Inflammatory and Hypoxic Stress. Diabetologia 2019, 62, 1901–1914. [Google Scholar] [CrossRef]
- Kanak, M.A.; Takita, M.; Itoh, T.; SoRelle, J.A.; Murali, S.; Kunnathodi, F.; Shahbazov, R.; Lawrence, M.C.; Levy, M.F.; Naziruddin, B. Alleviation of Instant Blood-Mediated Inflammatory Reaction in Autologous Conditions through Treatment of Human Islets with NF-κB Inhibitors. Transplantation 2014, 98, 578. [Google Scholar] [CrossRef]
- Yoshimatsu, G.; Kunnathodi, F.; Saravanan, P.B.; Shahbazov, R.; Chang, C.; Darden, C.M.; Zurawski, S.; Boyuk, G.; Kanak, M.A.; Levy, M.F.; et al. Pancreatic β-Cell–Derived IP-10/CXCL10 Isletokine Mediates Early Loss of Graft Function in Islet Cell Transplantation. Diabetes 2017, 66, 2857–2867. [Google Scholar] [CrossRef]
- Wienhöfer, L.; Marker, M.; Antoni, A.-C.; Sutter, K.; Sander, A.; Dudda, M.; Flohé, S.B. TLR4 Transactivates CD8+ T Lymphocytes upon Acute Sterile Tissue Injury. ImmunoHorizons 2021, 5, 298–306. [Google Scholar] [CrossRef]
- GILLA, R.G. Antigen Presentation Pathways for Immunity to Islet Transplants: Relevance to Immunoisolation. Ann. N. Y. Acad. Sci. 1999, 875, 255–260. [Google Scholar] [CrossRef]
- Hughes, A.D.; Zhao, D.; Dai, H.; Abou-Daya, K.I.; Tieu, R.; Rammal, R.; Williams, A.L.; Landsittel, D.P.; Shlomchik, W.D.; Morelli, A.E.; et al. Cross-Dressed Dendritic Cells Sustain Effector T Cell Responses in Islet and Kidney Allografts. J. Clin. Investig. 2020, 130, 287–294. [Google Scholar] [CrossRef]
- Tripathy, A.; Khanna, S.; Padhan, P.; Smita, S.; Raghav, S.; Gupta, B. Direct Recognition of LPS Drive TLR4 Expressing CD8+ T Cell Activation in Patients with Rheumatoid Arthritis. Sci. Rep. 2017, 7, 933. [Google Scholar] [CrossRef]
- Sha, T.; Sunamoto, M.; Kitazaki, T.; Sato, J.; Ii, M.; Iizawa, Y. Therapeutic Effects of TAK-242, a Novel Selective Toll-like Receptor 4 Signal Transduction Inhibitor, in Mouse Endotoxin Shock Model. Eur. J. Pharmacol. 2007, 571, 231–239. [Google Scholar] [CrossRef]
- Liu, Y.; Pu, X.; Qin, X.; Gong, J.; Huang, Z.; Luo, Y.; Mou, T.; Zhou, B.; Shen, A.; Wu, Z. Neutrophil Extracellular Traps Regulate HMGB1 Translocation and Kupffer Cell M1 Polarization During Acute Liver Transplantation Rejection. Front. Immunol. 2022, 13, 823511. [Google Scholar] [CrossRef]
- Leopold Wager, C.M.; Wormley, F.L. Classical versus Alternative Macrophage Activation: The Ying and the Yang in Host Defense against Pulmonary Fungal Infections. Mucosal Immunol. 2014, 7, 1023–1035. [Google Scholar] [CrossRef]
- Kunkel, S.L.; Standiford, T.; Kasahara, K.; Strieter, R.M. Interleukin-8 (IL-8): The Major Neutrophil Chemotactic Factor in the Lung. Exp. Lung Res. 1991, 17, 17–23. [Google Scholar] [CrossRef]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte Chemoattractant Protein-1 (MCP-1): An Overview. J. Interferon Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef]
- Montolio, M.; Biarnés, M.; Téllez, N.; Escoriza, J.; Soler, J.; Montanya, E. Interleukin-1β and Inducible Form of Nitric Oxide Synthase Expression in Early Syngeneic Islet Transplantation. J. Endocrinol. 2007, 192, 169–177. [Google Scholar] [CrossRef]
- Naziruddin, B.; Iwahashi, S.; Kanak, M.A.; Takita, M.; Itoh, T.; Levy, M.F. Evidence for Instant Blood-Mediated Inflammatory Reaction in Clinical Autologous Islet Transplantation. Am. J. Transplant. 2014, 14, 428–437. [Google Scholar] [CrossRef]
- Naziruddin, B.; Kanak, M.A.; Chang, C.A.; Takita, M.; Lawrence, M.C.; Dennison, A.R.; Onaca, N.; Levy, M.F. Improved Outcomes of Islet Autotransplant after Total Pancreatectomy by Combined Blockade of IL-1β and TNFα. Am. J. Transplant. 2018, 18, 2322–2329. [Google Scholar] [CrossRef]
- Baker, M.S.; Chen, X.; Rotramel, A.R.; Nelson, J.J.; Lu, B.; Gerard, C.; Kanwar, Y.; Kaufman, D.B. Genetic Deletion of Chemokine Receptor CXCR3 or Antibody Blockade of Its Ligand IP-10 Modulates Posttransplantation Graft-Site Lymphocytic Infiltrates and Prolongs Functional Graft Survival in Pancreatic Islet Allograft Recipients. Surgery 2003, 134, 126–133. [Google Scholar] [CrossRef]
- Piemonti, L.; Leone, B.E.; Nano, R.; Saccani, A.; Monti, P.; Maffi, P.; Bianchi, G.; Sica, A.; Peri, G.; Melzi, R.; et al. Human Pancreatic Islets Produce and Secrete MCP-1/CCL2: Relevance in Human Islet Transplantation. Diabetes 2002, 51, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.R.; Ma, A.C.; Tavener, S.A.; McDonald, B.; Goodarzi, Z.; Kelly, M.M.; Patel, K.D.; Chakrabarti, S.; McAvoy, E.; Sinclair, G.D.; et al. Platelet TLR4 Activates Neutrophil Extracellular Traps to Ensnare Bacteria in Septic Blood. Nat. Med. 2007, 13, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Krüger, B.; Lal, G.; Luan, Y.; Yadav, A.; Zang, W.; Grimm, M.; Waaga-Gasser, A.M.; Murphy, B.; Bromberg, J.S.; et al. Inhibition of TLR4 Signaling Prolongs Treg-Dependent Murine Islet Allograft Survival. Immunol. Lett. 2010, 127, 119–125. [Google Scholar] [CrossRef] [PubMed][Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Value (n = 10) |
|---|---|
| Gender (female:male) | 8:2 |
| Age (years) | 40 ± 11.7 |
| Height (cm) | 165 ± 9.88 |
| Weight (kg) | 70.2 ± 11.82 |
| Body mass index (kg/m2) | 25.75 ± 3.71 |
| Disease duration (years) | 9.4 ± 5.9 |
| Fasting blood glucose (mg/dL) | 91.1 ± 12.3 |
| Stimulated blood glucose (mg/dL) | 153.5 ± 54.5 |
| Basal C-peptide (ng/mL) | 1.91 ± 1.39 |
| Stimulated C-peptide (ng/mL) | 7.39 ± 4.48 |
| ∆ C-peptide (ng/mL) | 5.48 ± 3.47 |
| Initial trimmed pancreas weight (g) | 134.6 ± 25.3 |
| Pancreas weight processed (g) | 88.0 ± 23.3 |
| Total islet yield (IEQ) | 560,973 ± 125,830 |
| Islet particle number (IN) | 335,700 ± 96,743 |
| Islet yield (IEQ/g pancreas) | 5986 ± 2012 |
| Dose (IEQ/kg patient) | 8147 ± 2040 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mattke, J.; Darden, C.M.; Vasu, S.; Lawrence, M.C.; Kirkland, J.; Kane, R.R.; Naziruddin, B. Inhibition of Toll-like Receptor 4 Using Small Molecule, TAK-242, Protects Islets from Innate Immune Responses. Cells 2024, 13, 416. https://doi.org/10.3390/cells13050416
Mattke J, Darden CM, Vasu S, Lawrence MC, Kirkland J, Kane RR, Naziruddin B. Inhibition of Toll-like Receptor 4 Using Small Molecule, TAK-242, Protects Islets from Innate Immune Responses. Cells. 2024; 13(5):416. https://doi.org/10.3390/cells13050416
Chicago/Turabian StyleMattke, Jordan, Carly M. Darden, Srividya Vasu, Michael C. Lawrence, Jeffrey Kirkland, Robert R. Kane, and Bashoo Naziruddin. 2024. "Inhibition of Toll-like Receptor 4 Using Small Molecule, TAK-242, Protects Islets from Innate Immune Responses" Cells 13, no. 5: 416. https://doi.org/10.3390/cells13050416
APA StyleMattke, J., Darden, C. M., Vasu, S., Lawrence, M. C., Kirkland, J., Kane, R. R., & Naziruddin, B. (2024). Inhibition of Toll-like Receptor 4 Using Small Molecule, TAK-242, Protects Islets from Innate Immune Responses. Cells, 13(5), 416. https://doi.org/10.3390/cells13050416