
ODF2 Negatively Regulates CP110 Levels at the Centrioles/Basal Bodies to Control the Biogenesis of Primary Cilia


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmid Constructs
2.2. Immune-Cytology and Protein Quantification
2.3. Transfection of Cells
2.4. Rapamycin-Induced Dimerization
3. Results
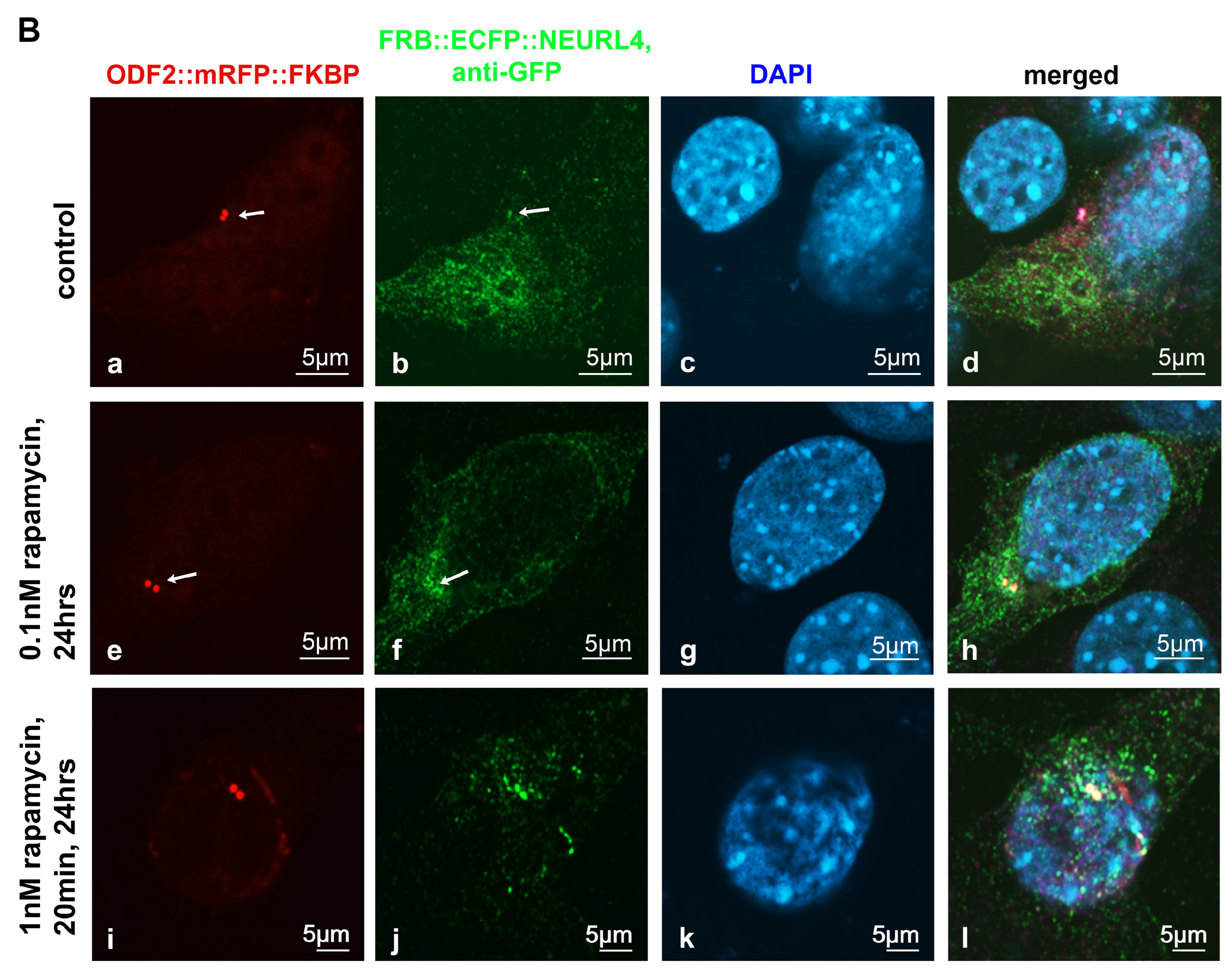
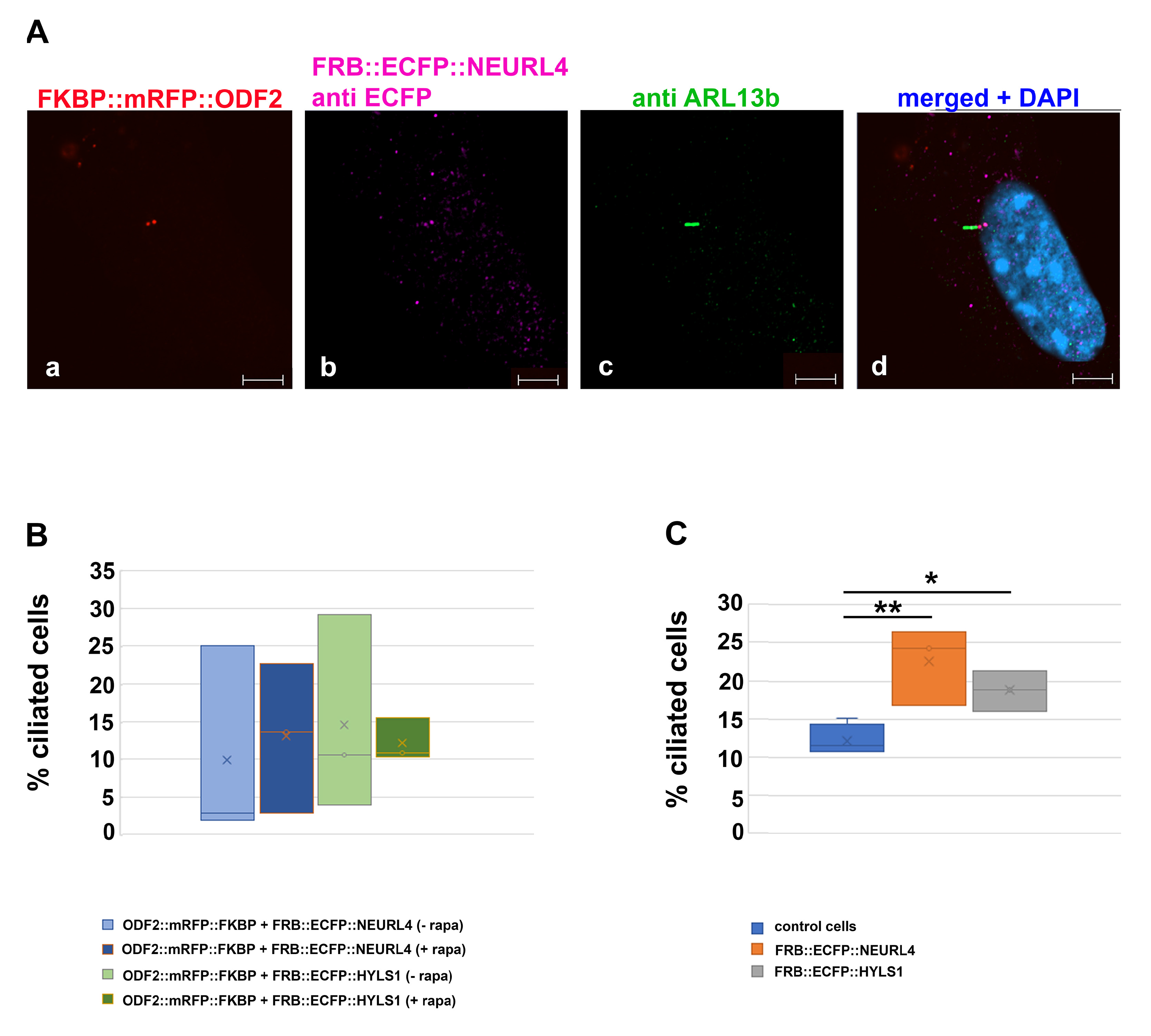
3.1. ODF2-Mediated Recruitment of NEURL4 to the Centrosome
3.2. Rapamycin-Induced Recruitment of NEURL4 to the Centrosome Did Not Enhance the Loss of CP110
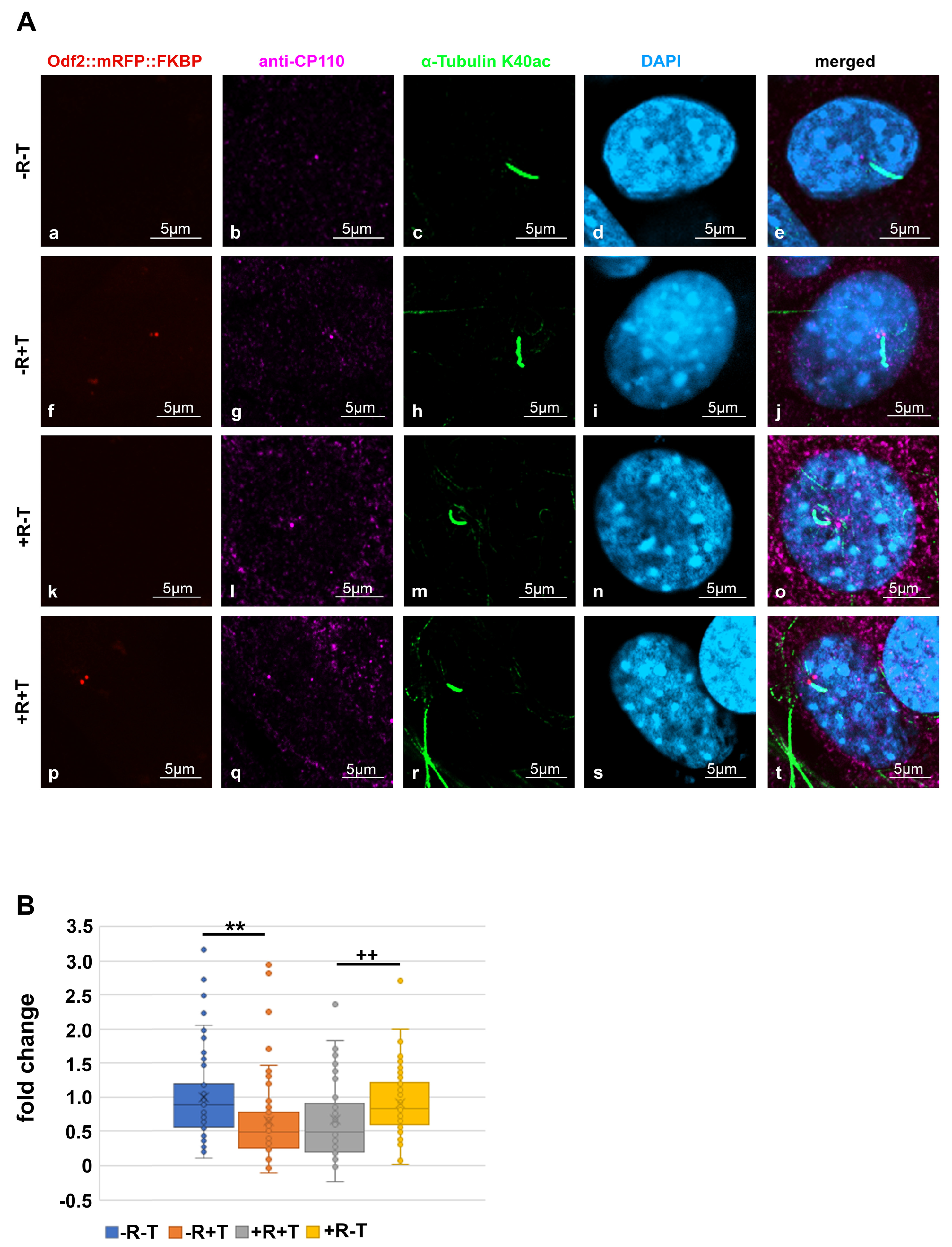
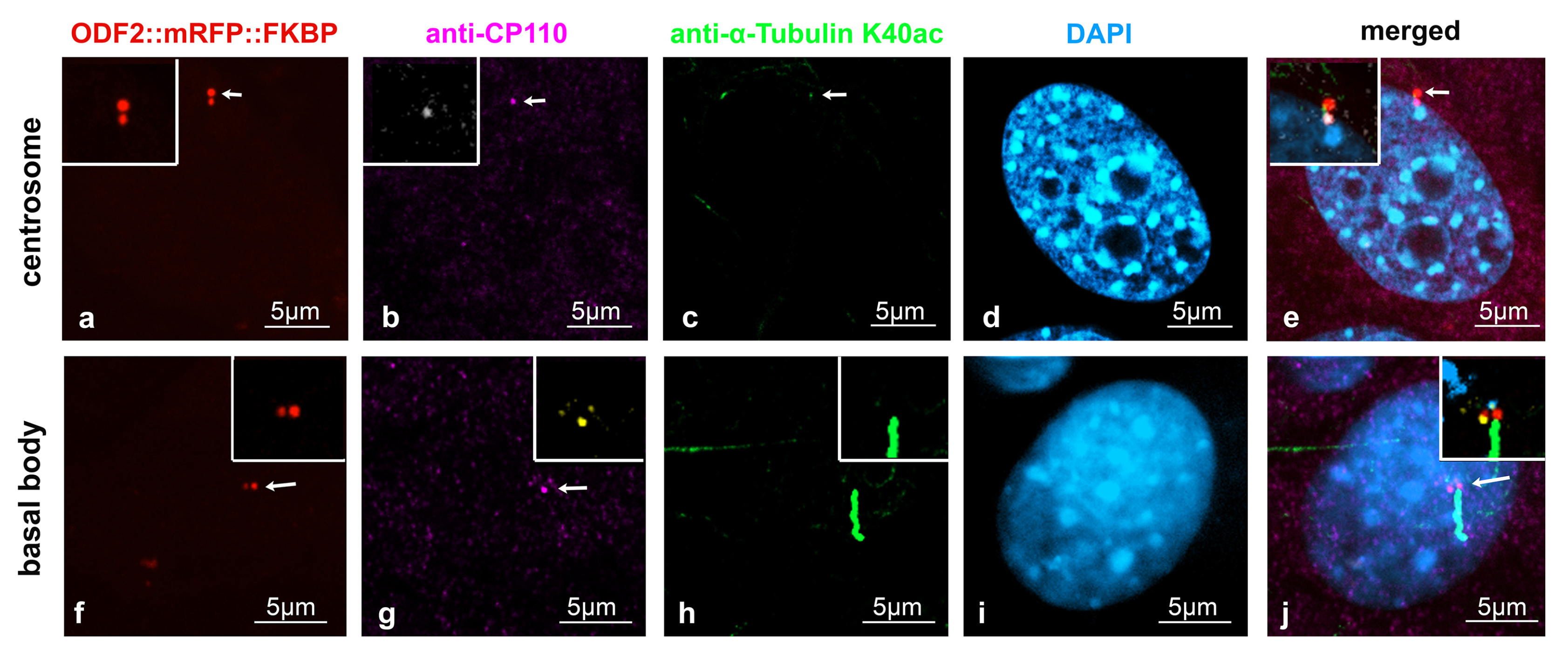
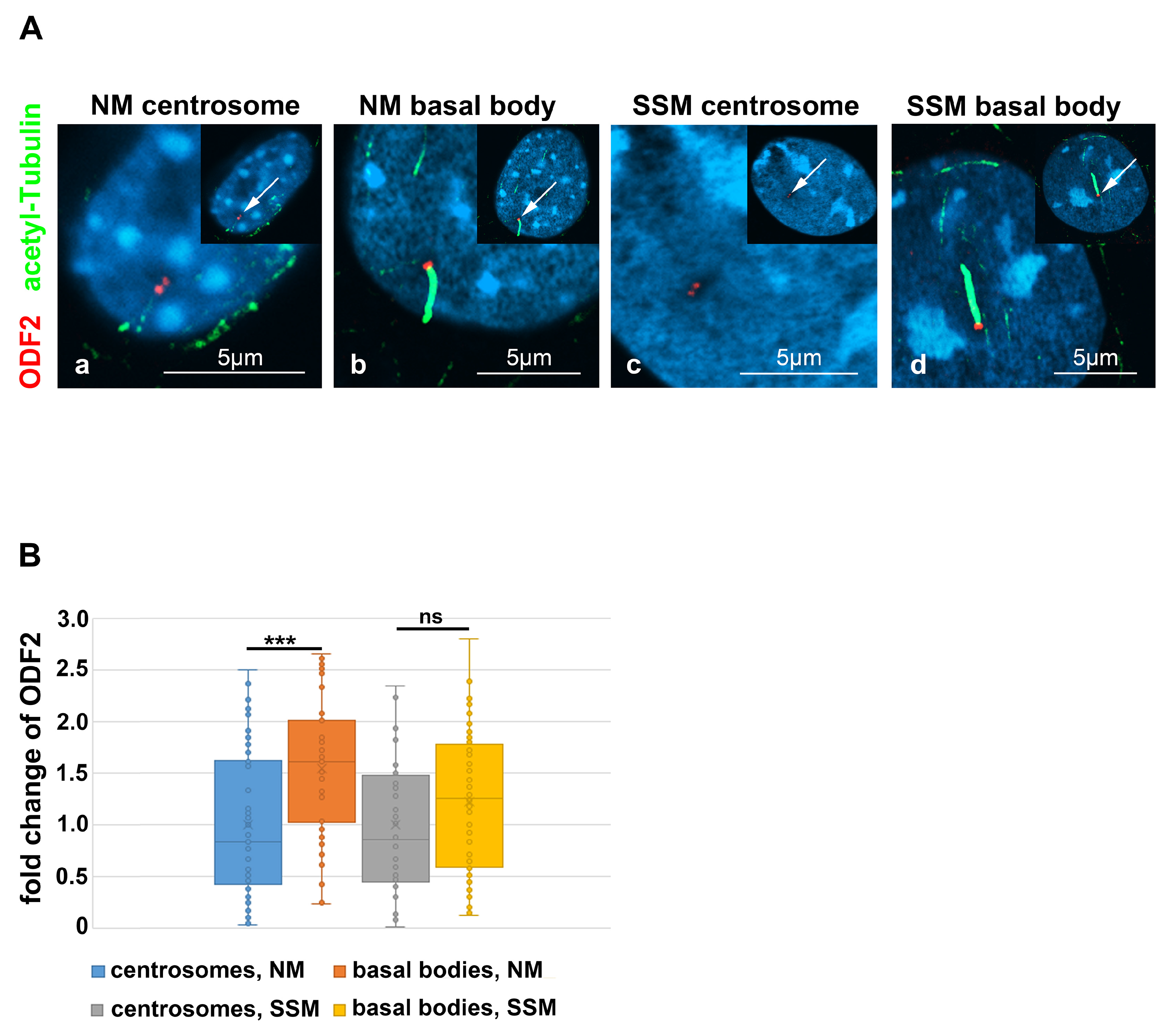
3.3. Distribution of ODF2 and CP110 at the Centrosome and the Basal Body
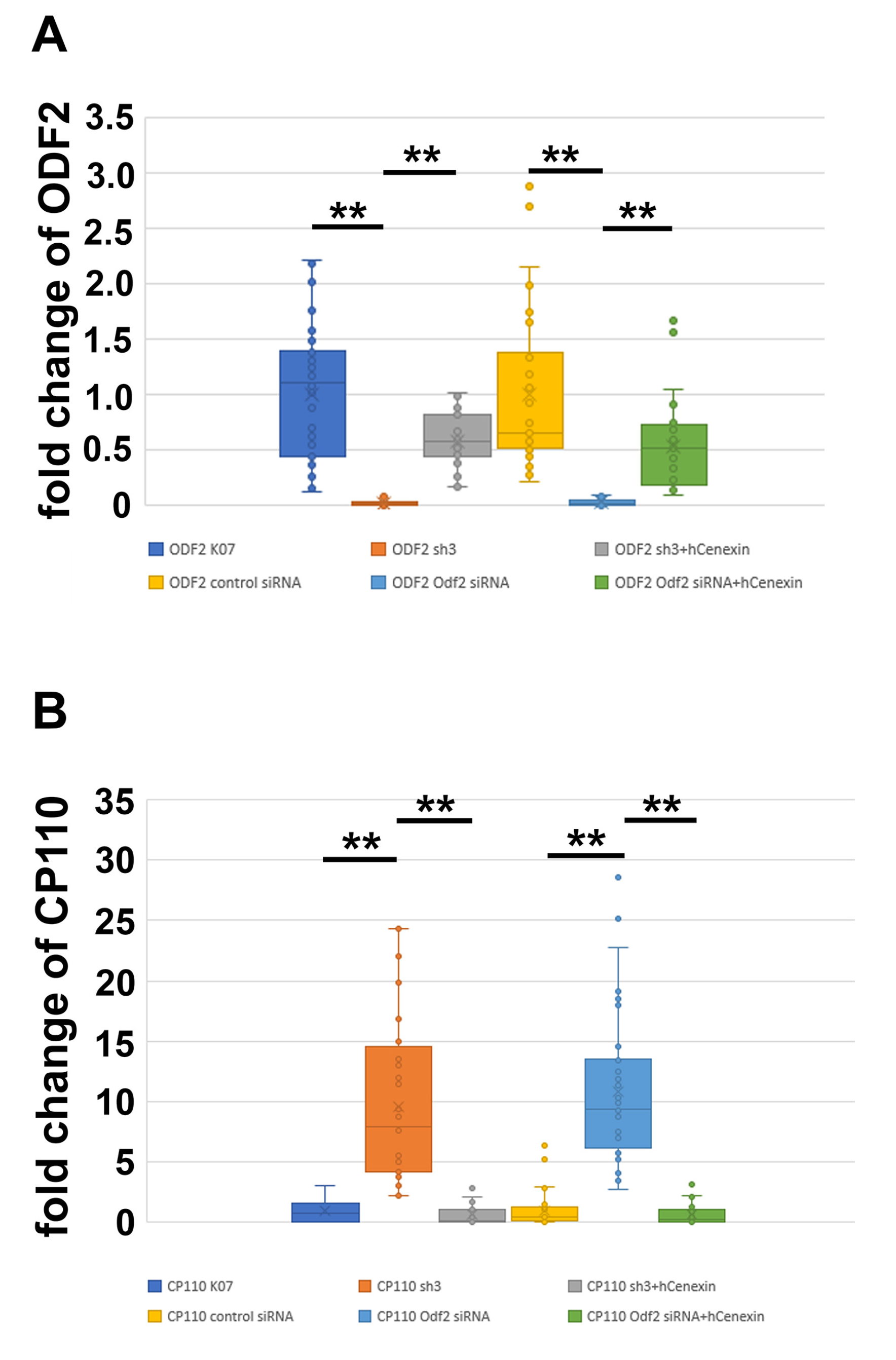
3.4. ODF2 Promotes Depletion of Centrosomal CP110
3.5. Enrichment of ODF2 at the Basal Body
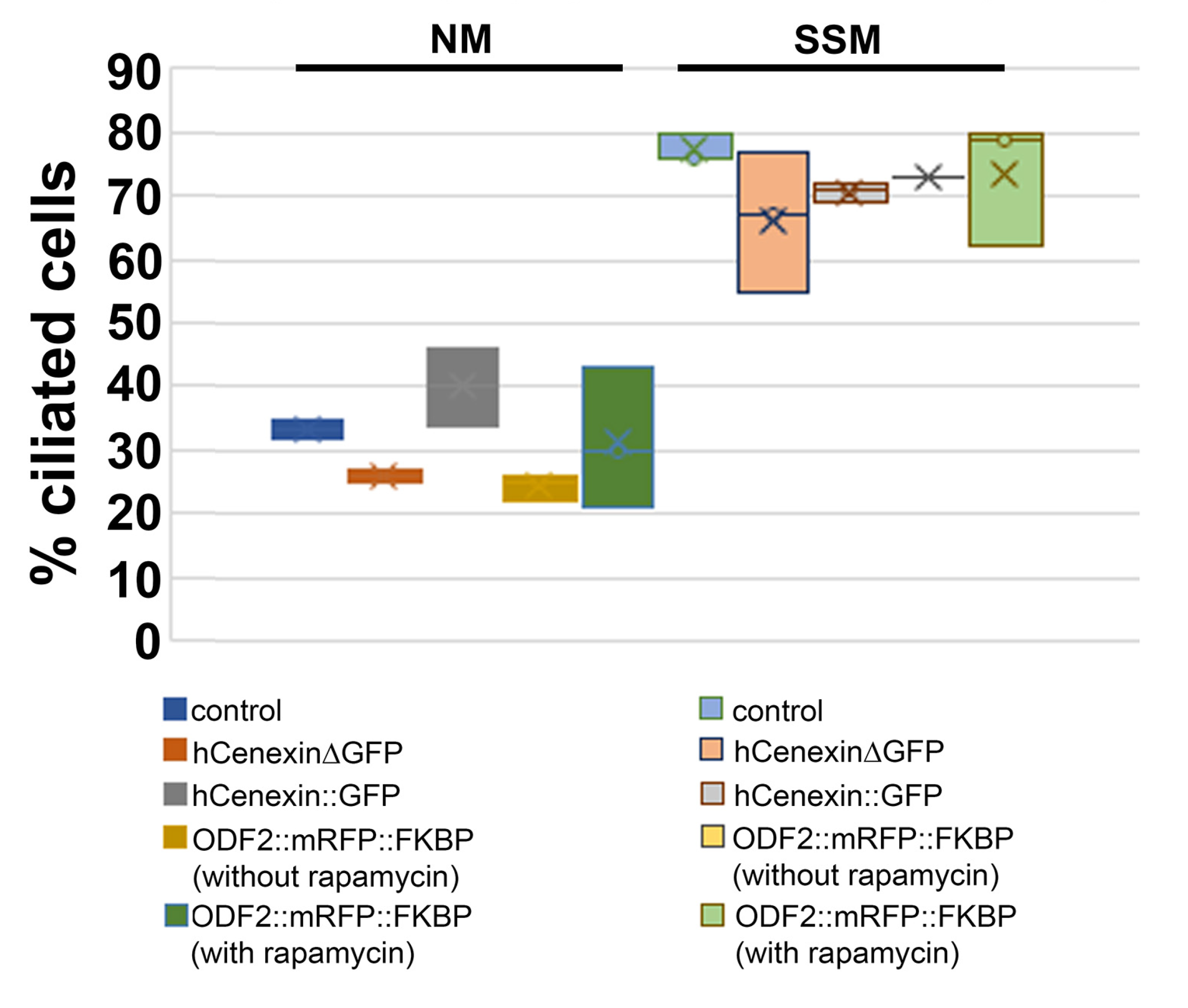
3.6. The overexpression of ODF2 Did Not Promote Cilia Formation
3.7. Ciliogenesis and Neurl4 Expression
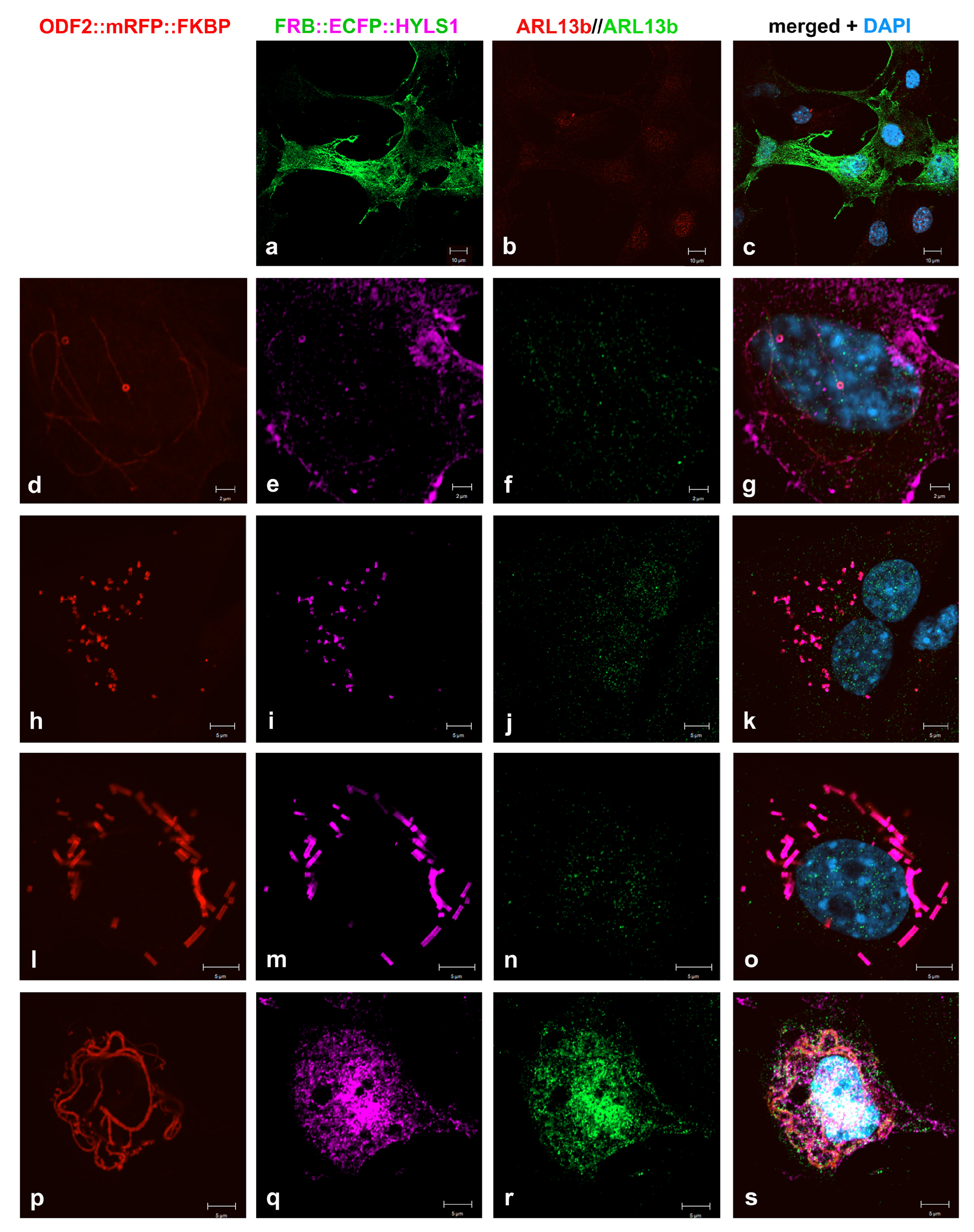
3.8. Formation of Higher Order Structures by Interaction between HYLS1 and ODF2
4. Discussion
4.1. Rapamycin-Induced Targeting to the Centriole
4.2. ODF2 Controls CP110 Levels
4.3. ODF2 Is Not Sufficient to Promote Cilia Formation
4.4. Co-Localization of ODF2 and HYLS1
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Badano, J.L.; Mitsuma, N.; Beales, P.L.; Katsanis, N. The ciliopathies: An emerging class of human genetic disorders. Annu. Rev. Genom. Hum. Genet. 2006, 7, 125–148. [Google Scholar] [CrossRef] [PubMed]
- Satir, P.; Christensen, T. Overview of structure and function of mammalian cilia. Annu. Rev. Physiol. 2007, 69, 377–400. [Google Scholar] [CrossRef] [PubMed]
- Gerdes, M.; Davis, E.E.; Katsanis, N. The vertebrate primary cilium in development, homeostasis, and disease. Cell 2009, 137, 32–45. [Google Scholar] [CrossRef] [PubMed]
- Goetz, S.C.; Anderson, K.V. The primary cilium: A signalling center during vertebrate development. Nat. Rev. Genet. 2010, 11, 331–344. [Google Scholar] [CrossRef]
- Fisch, C.; Dupuis-Williams, P. Ultrastructure of cilia and flagella—back to the future! Biol. Cell 2011, 103, 249–270. [Google Scholar] [CrossRef]
- Hoyer-Fender, S. Primary and motile cilia: Their Ultrastructure and Ciliogenesis. In Cilia and Nervous System Development and Function; Tucker, K.L., Caspary, T., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 1–53. ISBN 978-94-007-5808-7. [Google Scholar]
- Reiter, J.; Leroux, M.R. Genes and molecular pathways underpinning ciliopathies. Nat. Rev. Mol. Cell Biol. 2017, 18, 533–547. [Google Scholar] [CrossRef]
- Higgins, M.; Obaidi, I.; McMorrow, T. Primary cilia and their role in cancer. Oncol. Lett. 2019, 17, 3041–3047. [Google Scholar] [CrossRef]
- Wheatley, D.N. Cilia in cell-cultured fibroblasts. III. Relationship between mitotic activity and cilium frequency in mouse 3T6 fibroblasts. J. Anat. 1971, 110, 367–382. [Google Scholar]
- Tucker, R.W.; Pardee, A.B.; Fujiwara, K. Centriole ciliation is related to quiescence and DNA synthesis in 3T3 cells. Cell 1979, 17, 527–535. [Google Scholar] [CrossRef]
- Quarmby, L.M.; Parker, J.D.K. Cilia and the cell cycle. J. Cell Biol. 2005, 169, 707–710. [Google Scholar] [CrossRef]
- Santos, N.; Reiter, J.F. Building it up and taking it down: The regulation of vertebrate ciliogenesis. Dev. Dyn. 2008, 237, 1972–1981. [Google Scholar] [CrossRef] [PubMed]
- Plotnikova, O.V.; Pugacheva, E.N.; Golemis, E.A. Primary cilia and the cell cycle. Methods Cell Biol. 2009, 94, 137–160. [Google Scholar] [PubMed]
- Seeley, E.S.; Nachury, M.V. The perennial organelle: Assembly and disassembly of the primary cilium. J. Cell Sci. 2010, 123, 511–518. [Google Scholar] [CrossRef]
- Kim, S.K.; Tsiokas, L. Cilia and cell cycle re-entry. Cell Cycle 2011, 10, 2683–2690. [Google Scholar] [CrossRef] [PubMed]
- Avidor-Reiss, T.; Gopalakrishnan, J. Cell cycle regulation of the centrosome and cilium. Drug Discov. Today Dis. Mech. 2013, 10, e119–e124. [Google Scholar] [CrossRef]
- Ishikawa, H.; Marshall, W.F. Ciliogenesis: Building the cell’s antenna. Nat. Rev. Mol. Cell Biol. 2011, 12, 222–234. [Google Scholar] [CrossRef]
- Kumar, D.; Reiter, J. How the centriole builds its cilium: Of mothers, daughters, and the acquisition of appendages. Curr. Opin. Struct. Biol. 2021, 66, 41–48. [Google Scholar] [CrossRef]
- Bornens, M. Centrosome composition and microtubule anchoring mechanisms. Curr. Opin. Cell Biol. 2002, 14, 25–34. [Google Scholar] [CrossRef]
- Fliegauf, M.; Benzing, T.; Omran, H. When cilia go bad: Cilia defects and ciliopathies. Nat. Rev. Mol. Cell Biol. 2007, 8, 880–893. [Google Scholar] [CrossRef]
- Pedersen, L.B.; Veland, I.R.; Schroder, J.M.; Christensen, S.T. Assembly of primary cilia. Dev. Dyn. 2008, 237, 1993–2006. [Google Scholar] [CrossRef]
- Van Dam, T.J.; Kennedy, J.; van der Lee, R.; De Vrieze, E.; Wunderlich, K.A.; Rix, S.; Dougherty, G.W.; Lambacher, N.J.; Li, C.; Jensen, V.L.; et al. CiliaCarta: An integrated and validated compendium of ciliary genes. PLoS ONE 2019, 14, e0216705. [Google Scholar] [CrossRef]
- Zou, C.; Li, J.; Bai, Y.; Gunning, W.T.; Wazer, D.E.; Band, V.; Gao, Q. Centrobin: A novel daughter centriole-associated protein that is required for centriole duplication. J. Cell Biol. 2005, 171, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Mahjoub, M.R.; Xie, Z.; Stearns, T. Cep120 is asymmetrically localized to the daughter centriole and is essential for centriole assembly. J. Cell Biol. 2010, 191, 331–346. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Kim, S.; Kobayashi, T.; Liang, F.X.; Korzeniewski, N.; Duensing, S.; Dynlacht, B.D. Neurl4, a novel daughter centriole protein, prevents formation of ectopic microtubule organizing centres. EMBO Rep. 2012, 13, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.T.; Chong, W.M.; Wang, W.J.; Mazo, G.; Tanos, B.; Chen, Z.; Tran, T.M.N.; Chen, Y.D.; Weng, R.R.; Huang, C.E.; et al. Super-resolution architecture of mammalian centriole distal appendages reveals distinct blade and matrix functional components. Nat. Commun. 2018, 9, 2023. [Google Scholar] [CrossRef]
- Chong, W.M.; Wang, W.-J.; Lo, C.-H.; Chiu, T.-Y.; Chang, T.-J.; Liu, Y.-P.; Tanos, B.; Mazo, G.; Tsou, M.-F.B.; Jane, W.-N.; et al. Super-Resolution Microscopy Reveals Coupling between Mammalian Centriole Subdistal Appendages and Distal Appendages. eLife 2020, 9, e53580. [Google Scholar] [CrossRef]
- Graser, S.; Stierhof, Y.D.; Lavoie, S.B.; Gassner, O.S.; Lamla, S.; Le Clech, M.; Nigg, E.A. Cep164, a novel centriole appendage protein required for primary cilium formation. J. Cell Biol. 2007, 179, 321–330. [Google Scholar] [CrossRef]
- Schmidt, K.N.; Kuhns, S.; Neuner, A.; Hub, B.; Zentgraf, H.; Pereira, G. Cep164 mediates vesicular docking to the mother centriole during early steps of ciliogenesis. J. Cell Biol. 2012, 199, 1083–1101. [Google Scholar] [CrossRef]
- Sullenberger, C.; Vasquez-Limeta, A.; Kong, D.; Loncarek, J. With age comes maturity: Biochemical and structural transformation of a human centriole in the making. Cells 2020, 9, 1429. [Google Scholar] [CrossRef]
- Sánchez, I.; Dynlacht, B.D. Cilium assembly and disassembly. Nat. Cell Biol. 2016, 18, 711–717. [Google Scholar] [CrossRef]
- Tsang, W.Y.; Dynlacht, B.D. CP110 and its network of partners coordinately regulate cilia assembly. Cilia 2013, 2, 9. [Google Scholar] [CrossRef] [PubMed]
- Nagai, T.; Mukoyama, S.; Kagiwada, H.; Goshima, N.; Mizuno, K. Cullin-3-KCTD10-mediated CEP97 degradation prootes primary cilium formation. J. Cell Sci. 2018, 131, jcs219527. [Google Scholar] [CrossRef] [PubMed]
- Spektor, A.; Tsang, W.Y.; Khoo, D.; Dynlacht, B.D. Cep97 and CP110 suppress a cilia assembly program. Cell 2007, 130, 678–690. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.I.; Kleylein-Sohn, J.; Westendorf, J.; Le Clech, M.; Lavoie, S.B.; Stierhof, Y.D.; Nigg, E.A. Control of centriole length by CPAP and CP110. Curr. Biol. 2009, 19, 1005–1011. [Google Scholar] [CrossRef]
- Song, R.; Walentek, P.; Sponer, N.; Klimke, A.; Lee, J.S.; Dixon, G.; Harland, R.; Wan, Y.; Lishko, P.; Lize, M.; et al. miR-34/449 miRNAs are required for motile ciliogenesis by repressing cp110. Nature 2014, 510, 115–120. [Google Scholar] [CrossRef]
- Yadav, S.P.; Sharma, N.K.; Liu, C.; Dong, L.; Li, T.; Swaroop, A. Centrosomal protein CP110 controls maturation of the mother centriole during cilia biogenesis. Development 2016, 143, 1491–1501. [Google Scholar] [CrossRef]
- Walentek, P.; Quigley, I.K.; Sun, D.I.; Sajjan, U.K.; Kintner, C.; Harland, R.M. Ciliary transcription factors and miRNAs precisely regulate Cp110 levels required for ciliary adhesions and ciliogenesis. eLife 2016, 5, e17557. [Google Scholar] [CrossRef]
- D’Angiolella, V.; Donato, V.; Vijayakumar, S.; Saraf, A.; Florens, L.; Washburn, M.P.; Dynlacht, B.; Pagano, M. SCF (Cyclin F) controls centrosome homeostasis and mitotic fidelity through CP110 degradation. Nature 2010, 466, 138–142. [Google Scholar] [CrossRef]
- Li, J.; D’Angiolella, V.; Seeley, E.S.; Kim, S.; Kobayashi, T.; Fu, W.; Campos, E.I.; Pagano, M.; Dynlacht, B.D. USP33 regulates centrosome biogenesis via deubiquitination of the centriolar protein CP110. Nature 2013, 495, 255–259. [Google Scholar] [CrossRef]
- Loukil, A.; Tormanen, K.; Sütterlin, C. The daughter centriole controls ciliogenesis by regulating Neurl-4 localization at the centrosome. J. Cell Biol. 2017, 216, 1287–1300. [Google Scholar] [CrossRef]
- Al-Hakim, A.K.; Bashkurov, M.; Gingras, A.C.; Durocher, D.; Pelletier, L. Interaction proteomics identify NEURL4 and the HECT E3 ligase HERC2 as novel modulators of centrosome architecture. Mol. Cell. Proteom. 2012, 11, 014233. [Google Scholar] [CrossRef] [PubMed]
- Goetz, S.C.; Liem, K.F., Jr.; Anderson, K.C. The spinocerebellar ataxia-associated gene Tau tubulin kinase 2 controls the initiation of ciliogenesis. Cell 2012, 151, 847–858. [Google Scholar] [CrossRef] [PubMed]
- Cajanek, L.; Nigg, E.A. Cep164 triggers ciliogenesis by recruiting Tau tubulin kinase 2 to the mother centriole. Proc. Natl. Acad. Sci. USA 2014, 111, E2841–E2850. [Google Scholar] [CrossRef]
- Oda, T.; Chiba, S.; Nagai, T.; Mizuno, K. Binding to Cep164, but not EB1, is essential for centriolar localization of TTBK2 and its function in ciliogenesis. Genes Cells 2014, 19, 927–940. [Google Scholar] [CrossRef]
- Lo, C.H.; Lin, I.H.; Yang, T.T.; Huang, Y.C.; Tanos, B.E.; Chou, P.C.; Chang, C.W.; Tsay, Y.G.; Liao, J.C.; Wang, W.J. Phosphorylation of CEP83 by TTBK2 is necessary for cilia initiation. J. Cell Biol. 2019, 218, 3489–3505. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Zhang, Y.; Wei, Q.; Huang, Y.; Hu, J.; Ling, K. Phosphatidylinositol phosphate kinase PIPKIgamma and phosphatase INPP5E coordinate initiation of ciliogenesis. Nat. Cell Biol. 2016, 20, 175–185. [Google Scholar]
- Wang, L.; Dynlacht, B.D. The regulation of cilium assembly and disassembly in development and disease. Development 2018, 145, dev151407. [Google Scholar] [CrossRef]
- Dammermann, A.; Pemble, H.; Mitchell, B.J.; McLeod, I.; Yates, J.R., III; Kintner, C.; Desai, A.B.; Oegema, K. The hydrolethalus syndrome protein HYLS-1 links core centriole structure to cilia formation. Genes Dev. 2009, 23, 2046–2059. [Google Scholar] [CrossRef]
- Chen, C.; Xu, Q.; Zhang, Y.; Davies, B.A.; Huang, Y.; Katzmann, D.J.; Harris, P.C.; Hu, J.; Ling, K. Ciliopathy protein HYLS1 coordinates the biogenesis and signalling of primary cilia by activating the ciliary lipid kinase PIPKIɣ. Sci. Adv. 2021, 7, eabe3401. [Google Scholar] [CrossRef]
- Kuhns, S.; Schmidt, K.N.; Reymann, J.; Gilbert, D.F.; Neuner, A.; Hub, B.; Carvalho, R.; Wiedemann, P.; Zentgraf, H.; Erfle, H.; et al. The microtubule affinity regulating kinase MARK4 promotes axoneme extension during early ciliogenesis. J. Cell Biol. 2013, 200, 505–522. [Google Scholar] [CrossRef]
- Brohmann, H.; Pinnecke, S.; Hoyer-Fender, S. Identification and characterization of new cDNAs encoding outer dense fiber proteins of rat sperm. J. Biol. Chem. 1997, 272, 10327–10332. [Google Scholar] [CrossRef] [PubMed]
- Salmon, N.A.; Reijo Pera, R.A.; Xu, E.Y. A gene trap knockout of the abundant sperm tail protein, outer dense fiber 2, results in preimplantation lethality. Genesis 2006, 44, 515–522. [Google Scholar] [CrossRef]
- Ishikawa, H.; Kubo, A.; Tsukita, S.; Tsukita, S. Odf2-deficient mother centrioles lack distal/subdistal appendages and the ability to generate primary cilia. Nat. Cell Biol. 2005, 7, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.T.; Stearns, T. Centriole age underlies asynchronous primary cilium growth in mammalian cells. Curr. Biol. 2009, 19, 1498–1502. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, S.; Egerer, J.; Fuchs, E.; Haas, A.K.; Barr, F.A. Functional dissection of Rab GTPases involved in primary cilium formation. J. Cell Biol. 2007, 178, 363–369. [Google Scholar] [CrossRef]
- Wang, L.; Failler, M.; Fu, W.; Dynlacht, B.D. A distal centriolar protein network controls organelle maturation and asymmetry. Nat. Commun. 2018, 9, 3938. [Google Scholar] [CrossRef]
- Putyrski, M.; Schultz, C. Protein translocation as a tool: The current rapamycin story. FEBS Lett. 2012, 586, 2097–2105. [Google Scholar] [CrossRef]
- Hüber, D.; Geisler, S.; Monecke, S.; Hoyer-Fender, S. Molecular dissection of ODF2/Cenexin revealed a short stretch of amino acids necessary for targeting to the centrosome and the primary cilium. Eur. J. Cell Biol. 2008, 87, 137–146. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Tylkowski, M.A.; Yang, K.; Hoyer-Fender, S.; Stoykova, A. Pax6 controls centriole maturation in cortical progenitors through Odf2. Cell. Mol. Life Sci. 2014, 72, 1795–1809. [Google Scholar] [CrossRef]
- Soung, N.K.; Kang, Y.H.; Kim, K.; Kamijo, K.; Yoon, H.; Seong, Y.S.; Kuo, Y.L.; Miki, T.; Kim, S.R.; Kuriyama, R.; et al. Requirement of hCenexin for proper mitotic functions of polo-like kinase 1 at the centrosomes. Mol. Cell. Biol. 2006, 26, 8316–8335. [Google Scholar] [CrossRef] [PubMed]
- Tanos, B.E.; Yang, H.J.; Soni, R.; Wang, W.J.; Macaluso, F.P.; Asara, J.M.; Tsou, M.F. Centriole distal appendages promote membrane docking, leading to cilia initiation. Genes Dev. 2013, 27, 163–168. [Google Scholar] [CrossRef]
- Joo, K.; Kim, C.G.; Lee, M.S.; Moon, H.Y.; Lee, S.H.; Kim, M.J.; Kweon, H.S.; Park, W.Y.; Kim, C.H.; Gleeson, J.G.; et al. CCDC41 is required for ciliary vesicle docking to the mother centriole. Proc. Natl. Acad. Sci. USA 2013, 110, 5987–5992. [Google Scholar] [CrossRef] [PubMed]
- Sillibourne, J.E.; Hurbain, I.; Grand-Perret, T.; Goud, B.; Tran, P.; Bornens, M. Primary ciliogenesis requires the distal appendage component Cep123. Biol. Open 2013, 2, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Xu, Q.; Zhang, Y.; Li, Y.; Zhang, Q.; Hu, Z.; Harris, P.C.; Torres, V.E.; Ling, K.; Hu, J. Transition fibre protein FBF1 is required for the ciliary entry of assembled intraflagellar transport complexes. Nat. Commun. 2013, 4, 2750. [Google Scholar] [CrossRef]
- Ye, X.; Zeng, H.; Ning, G.; Reiter, J.F.; Liu, A. C2cd3 is critical for centriolar distal appendage assembly and ciliary vesicle docking in mammals. Proc. Natl. Acad. Sci. USA 2014, 111, 2164–2169. [Google Scholar] [CrossRef]
- Kumar, D.; Rains, A.; Herranz-Pérez, V.; Lu, Q.; Shi, X.; Swaney, D.L.; Stevenson, E.; Krogan, N.J.; Huang, B.; Westlake, C.; et al. A ciliopathy complex builds distal appendages to initiate ciliogenesis. J. Cell Biol. 2021, 220, e202011133. [Google Scholar] [CrossRef]
- Lin, Y.; Wu, C.; Lin, Y.; Hsu, W.; Tang, C.C.; Chag, C.; Tang, T.K. CEP120 interacts with CPAP and positively regulates centriole elongation. J. Cell Biol. 2013, 202, 211–219. [Google Scholar] [CrossRef]
- Tsang, W.Y.; Bossard, C.; Khanna, H.; Peranen, J.; Swaroop, A.; Malhotra, V.; Dynlacht, B.D. CP110 suppresses primary cilia formation through its interaction with CEP290, a protein deficient in human ciliary disease. Dev. Cell 2008, 15, 187–197. [Google Scholar] [CrossRef]
- Gonçalves, A.B.; Hasselbalch, S.K.; Joensen, B.B.; Patzke, S.; Martens, P.; Ohlsen, S.K.; Quinodoz, M.; Nikopoulos, K.; Suleiman, R.; Damsø Jeppesen, M.P.; et al. CEP78 functions downstream of CEP350 to control biogenesis of primary cilia by negatively regulating CP110 levels. eLife 2021, 10, e63731. [Google Scholar] [CrossRef]
- Massa, F.; Tammaro, R.; Prado, M.A.; Cesana, M.; Lee, B.H.; Finley, D.; Franco, B.; Morleo, M. The deubiquitinating enzyme Usp14 controls ciliogenesis and Hedgehog signaling. Hum. Mol. Genet. 2018, 28, 764–777. [Google Scholar] [CrossRef]
- Shen, X.L.; Yuan, J.F.; Qin, X.H.; Song, G.P.; Hu, H.B.; Tu, H.-Q.; Song, Z.-Q.; Li, P.-Y.; Xu, Y.-L.; Li, S.; et al. LUBAC regulates ciliogenesis by promoting CP110 removal from the mother centriole. J. Cell Biol. 2021, 221, e202105092. [Google Scholar] [CrossRef]
- Lupas, A.; Van Dyke, M.; Stock, J. Predicting coiled coils from protein sequences. Science 1991, 252, 1162–1164. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2021, 50, D439–D444. [Google Scholar] [CrossRef] [PubMed]
- Truebestein, L.; Leonard, T.A. Coiled-coils: The long and the short of it. Bioessays 2016, 38, 903–916. [Google Scholar] [CrossRef]
- Gupta, G.D.; Coyaud, E.; Goncalves, J.; Mojarad, B.A.; Liu, Y.; Wu, Q.; Gheiratmand, L.; Comartin, D.; Tkach, J.M.; Cheung, S.W.; et al. A dynamic protein interaction landscape of the human centrosome-cilium interface. Cell 2015, 163, 1484–1499. [Google Scholar] [CrossRef]
- Stark, C.; Breitkreutz, B.J.; Reguly, T.; Boucher, L.; Breitkreutz, A.; Tyers, M. BioGRID: A General Repository for Interaction Datasets. Nucleic Acids Res. 2006, 34, D535–D539. [Google Scholar] [CrossRef]
- Prosser, S.L.; Morrison, C.G. Centrin2 regulates CP110 removal in primary cilium formation. J. Cell Biol. 2015, 208, 693–701. [Google Scholar] [CrossRef]
- Ogungbenro, Y.A.; Tena, T.C.; Gaboriau, D.; Lalor, P.; Dockery, P.; Philipp, M.; Morrison, C.G. Centrobin controls primary ciliogenesis in vertebrates. J. Cell Biol. 2018, 217, 1205–1215. [Google Scholar] [CrossRef]
- Liao, J.C.; Yang, T.T.; Weng, R.R.; Kuo, C.T.; Chang, C.W. TTBK2: A Tau protein kinase beyond Tau phosphorylation. BioMed. Res. Intern. 2015, 2015, 575170. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.; Zhang, D.; Li, F.; Chai, P.; Wang, S.; Teng, J.; Chen, J. M-phase phosphoprotein 9 regulates ciliogenesis by modulating CP110-CEP97 complex localization at the mother centriole. Nat. Commun. 2018, 9, 4511. [Google Scholar] [CrossRef] [PubMed]
- Winey, M.; O’Toole, E. Centriole structure. Phil. Trans. R. Soc. B 2014, 369, 20130457. [Google Scholar] [CrossRef] [PubMed]
- Menkveld, R.; Holleboom, C.A.G.; Rhemrev, J.P.T. Measurement and significance of sperm morphology. Asian J. Androl. 2011, 13, 59–68. [Google Scholar] [CrossRef]
- Weber, K.; Waletzky, A.; Fendl, D.; Ordóñez, P.; Takawale, P.; Hein, F.; Riedel, W.; König, A.; Kunze, M.; Leoni, A.L.; et al. New method for sperm evaluation by 3-dimensional laser scanning microscopy in different laboratory animal species. Int. J. Toxicol. 2014, 33, 353–361. [Google Scholar] [CrossRef]
- Hou, Y.; Wu, Z.; Zhang, Y.; Chen, H.; Hu, J.; Guo, Y.; Peng, Y.; Wei, Q. Functional analysis of hydrolethalus syndrome protein HYLS1 in ciliogenesis and spermatogenesis in Drosophila. Front. Cell Dev. Biol. 2020, 8, 301. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivation Conditions | Plasmid Transfected | Ciliated Cells/Total Cells | % Ciliated Cells | Average % |
|---|---|---|---|---|
| Standard medium | not transfected (control) | 177/507 | 35 | 33.5 |
| 182/565 | 32 | |||
| hCenexinΔGFP | 136/514 | 27 | 26 | |
| 135/539 | 25 | |||
| hCenexin::GFP | 231/507 | 46 | 40 | |
| 177/523 | 34 | |||
| 138NC::mRFP::FKBP (without rapamycin) | 130/505 | 26 | 24 | |
| 110/502 | 22 | |||
| 127/503 | 25 | |||
| 138NC::mRFP::FKBP (with rapamycin) | 220/507 | 43 | 31 | |
| 153/505 | 30 | |||
| 102/498 | 21 | |||
| Serum-deprived | not transfected (control) | 390/511 | 76 | 77 |
| 402/503 | 80 | |||
| 392/518 | 76 | |||
| hCenexinΔGFP | 340/505 | 67 | 66 | |
| 394/512 | 77 | |||
| 282/509 | 55 | |||
| hCenexin::GFP | 360/508 | 71 | 71 | |
| 374/521 | 72 | |||
| 138NC::mRFP::FKBP (without rapamycin) | 379/550 | 69 | ||
| 365/501 | 73 | 73 | ||
| 138NC::mRFP::FKBP (with rapamycin) | 400/500 | 80 | 74 | |
| 405/510 | 79 | |||
| 231/370 | 62 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Otto, M.; Hoyer-Fender, S. ODF2 Negatively Regulates CP110 Levels at the Centrioles/Basal Bodies to Control the Biogenesis of Primary Cilia. Cells 2023, 12, 2194. https://doi.org/10.3390/cells12172194
Otto M, Hoyer-Fender S. ODF2 Negatively Regulates CP110 Levels at the Centrioles/Basal Bodies to Control the Biogenesis of Primary Cilia. Cells. 2023; 12(17):2194. https://doi.org/10.3390/cells12172194
Chicago/Turabian StyleOtto, Madeline, and Sigrid Hoyer-Fender. 2023. "ODF2 Negatively Regulates CP110 Levels at the Centrioles/Basal Bodies to Control the Biogenesis of Primary Cilia" Cells 12, no. 17: 2194. https://doi.org/10.3390/cells12172194
APA StyleOtto, M., & Hoyer-Fender, S. (2023). ODF2 Negatively Regulates CP110 Levels at the Centrioles/Basal Bodies to Control the Biogenesis of Primary Cilia. Cells, 12(17), 2194. https://doi.org/10.3390/cells12172194