
Lymphatic Endothelial-to-Myofibroblast Transition: A Potential New Mechanism Underlying Skin Fibrosis in Systemic Sclerosis

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Immunofluorescence on Human Skin Sections
2.2. Culture of Human Dermal Lymphatic Microvascular Endothelial Cells
2.3. Collection of Serum Samples
2.4. Cell Stimulation and Analysis of Cell Morphology
2.5. RNA Isolation, cDNA Synthesis and Quantitative SYBR Green Real-Time PCR
2.6. Western Blotting
2.7. Fluorescence Immunocytochemistry
2.8. Collagen Gel Contraction Assay
2.9. TGFβ1 Enzyme-Linked Immunosorbent Assay
2.10. Statistical Analysis
3. Results
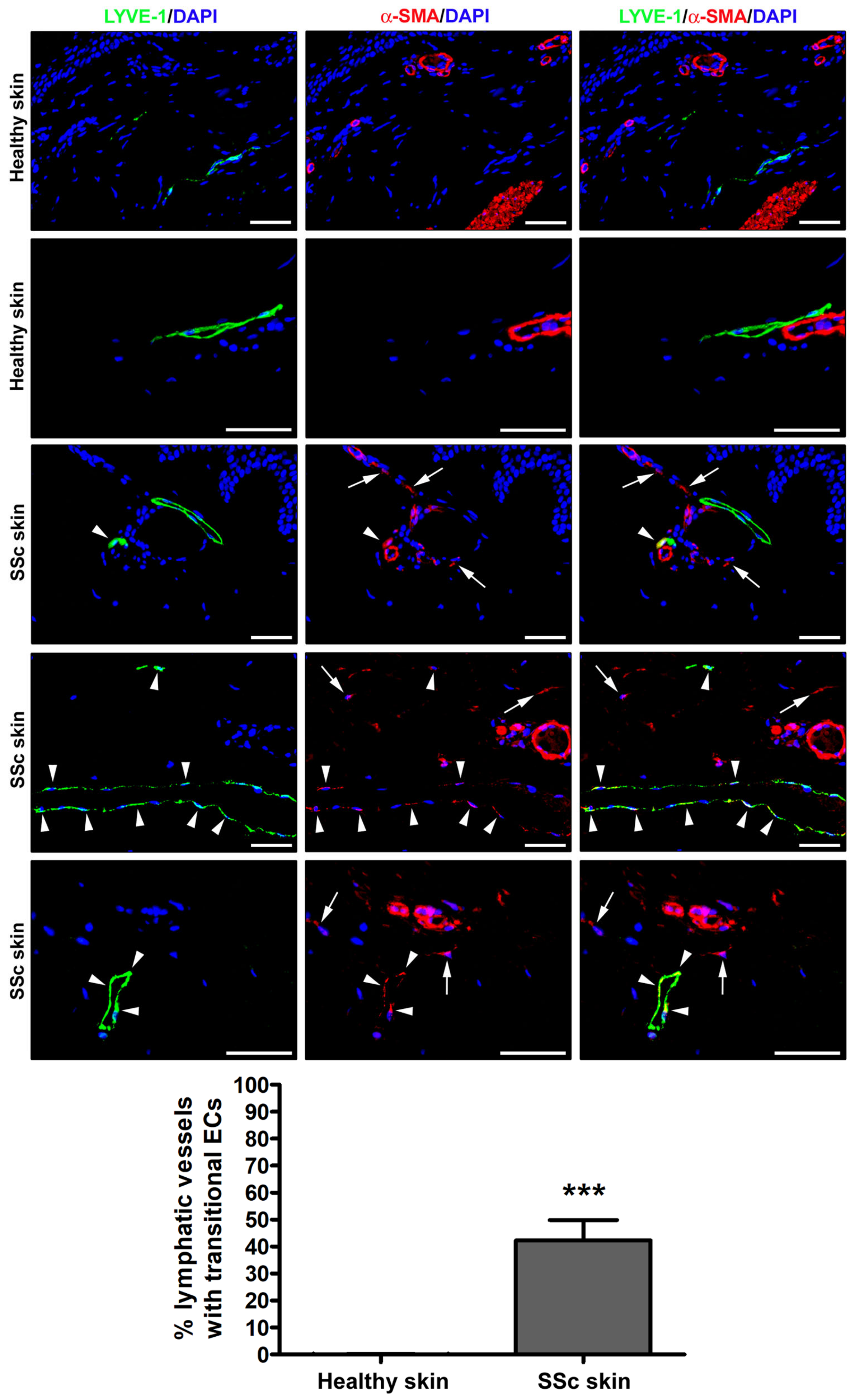
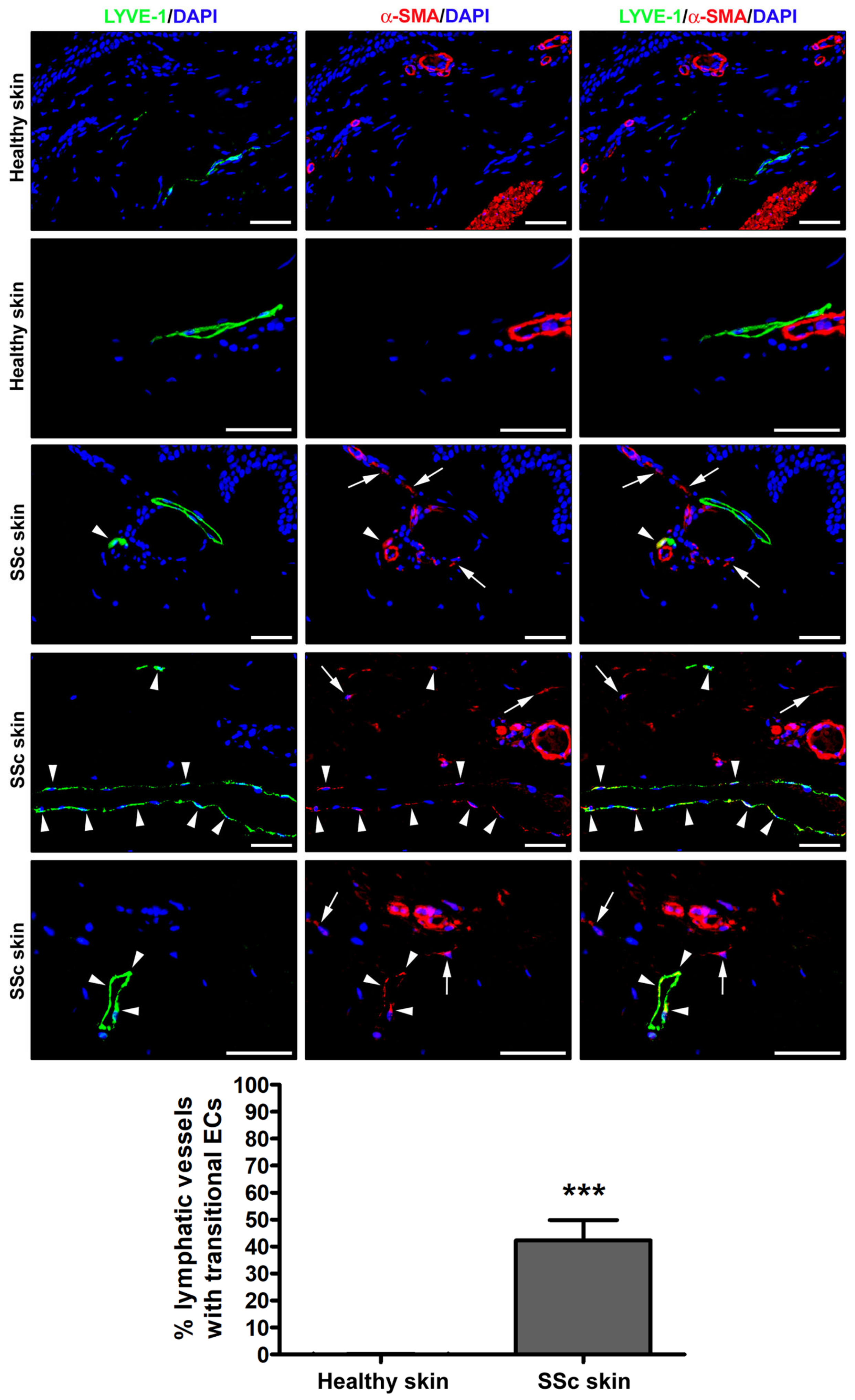
3.1. Detection of Ly-EndMT in the Lesional Skin of Patients with SSc
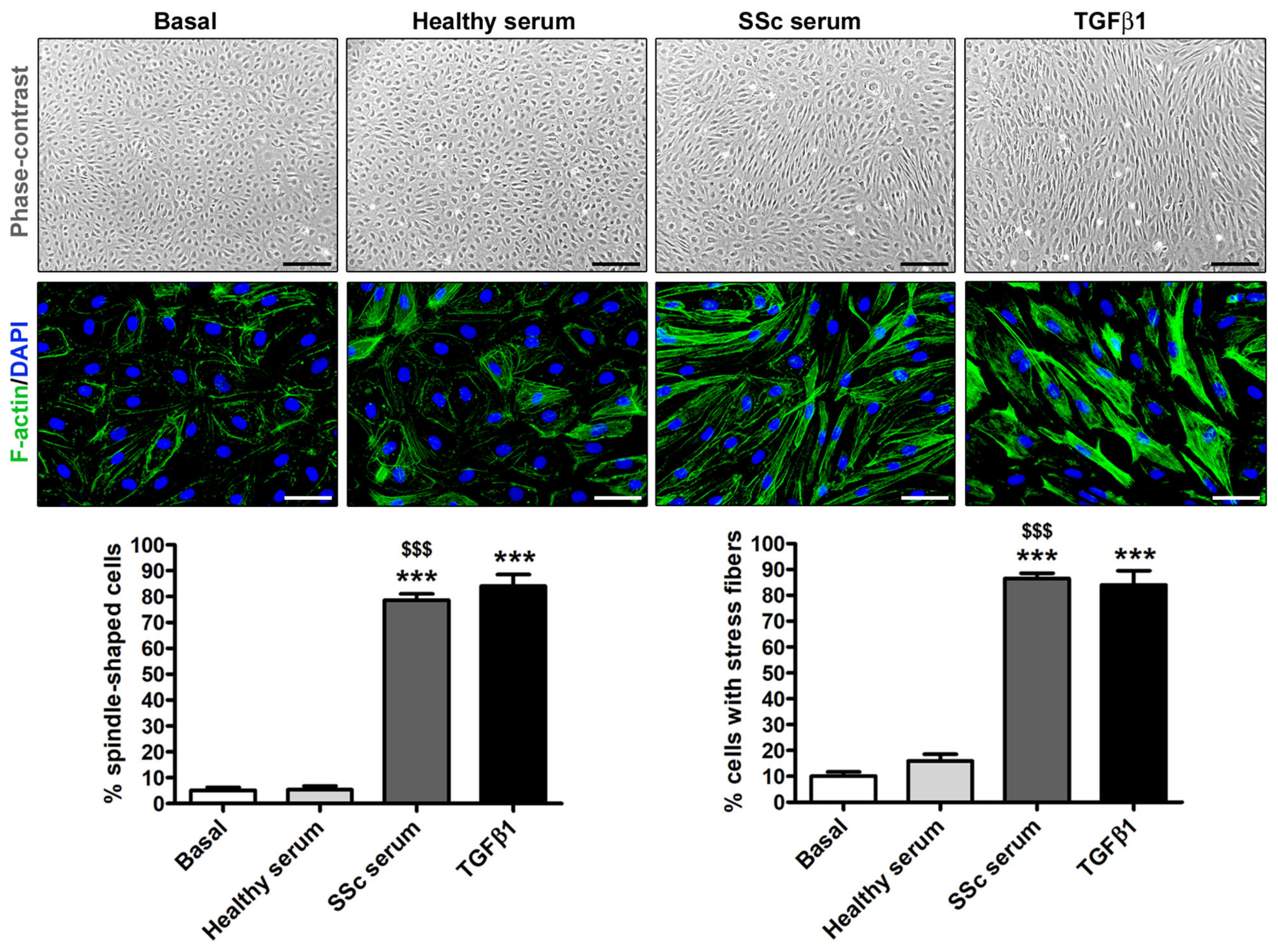
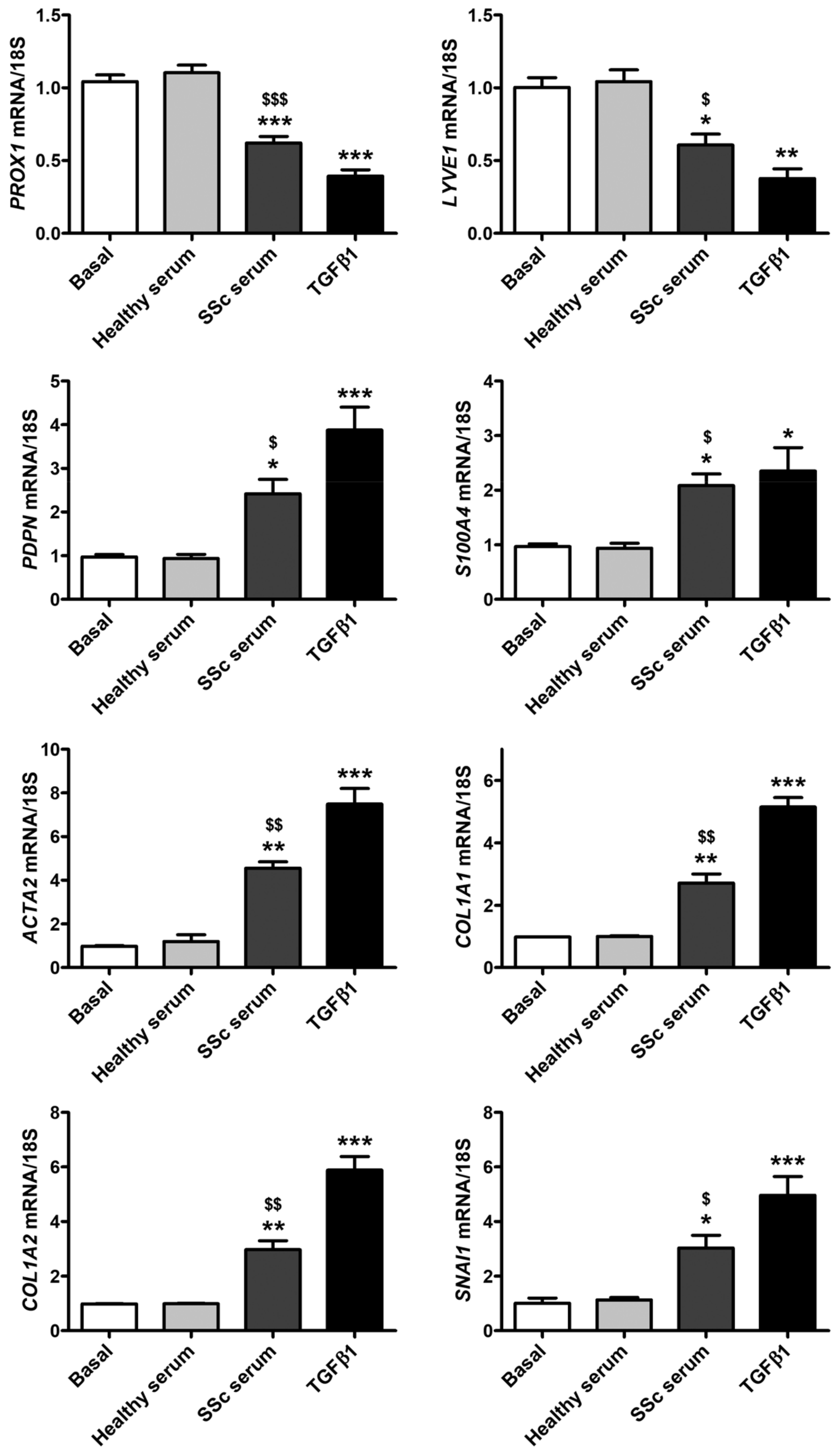
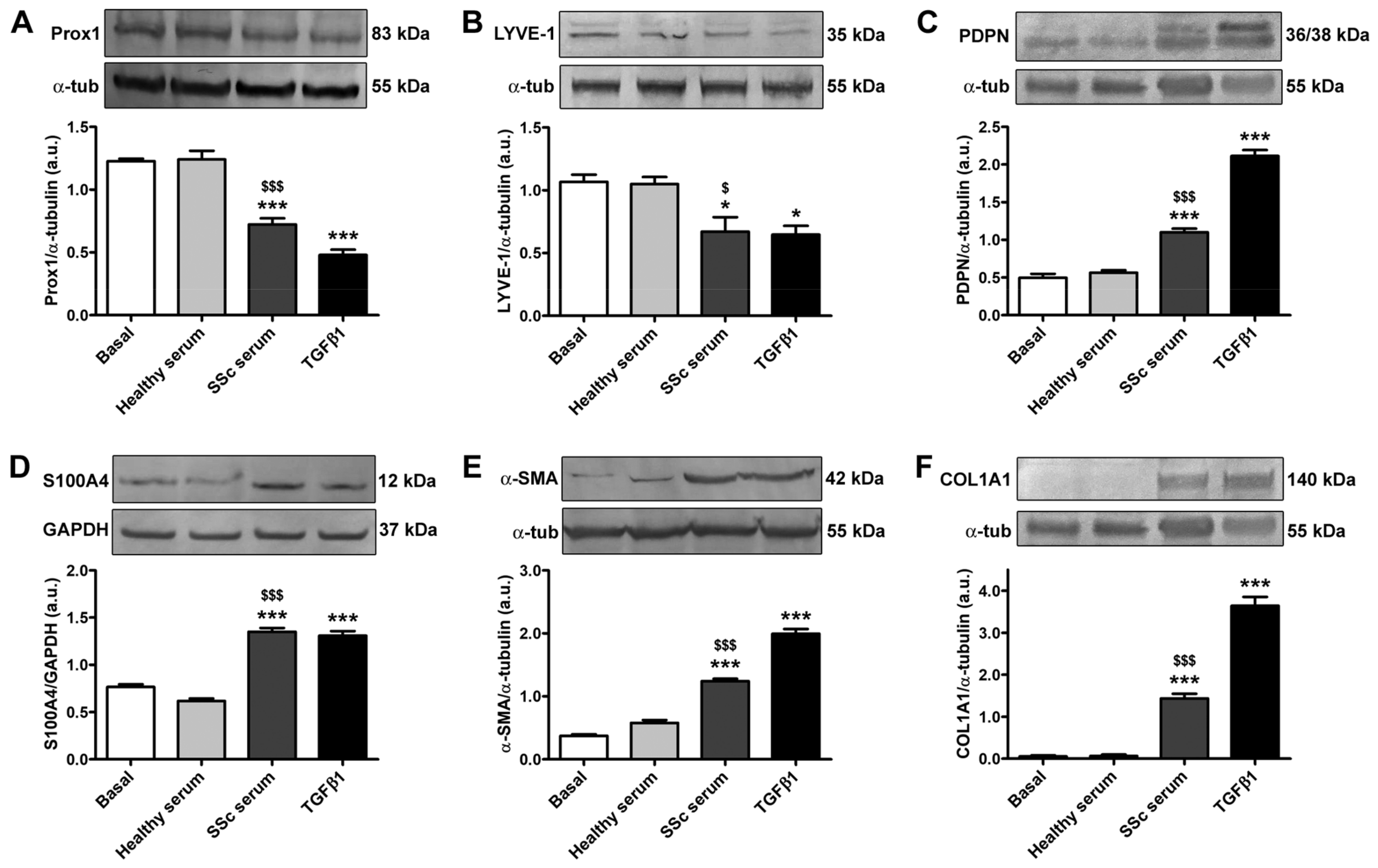
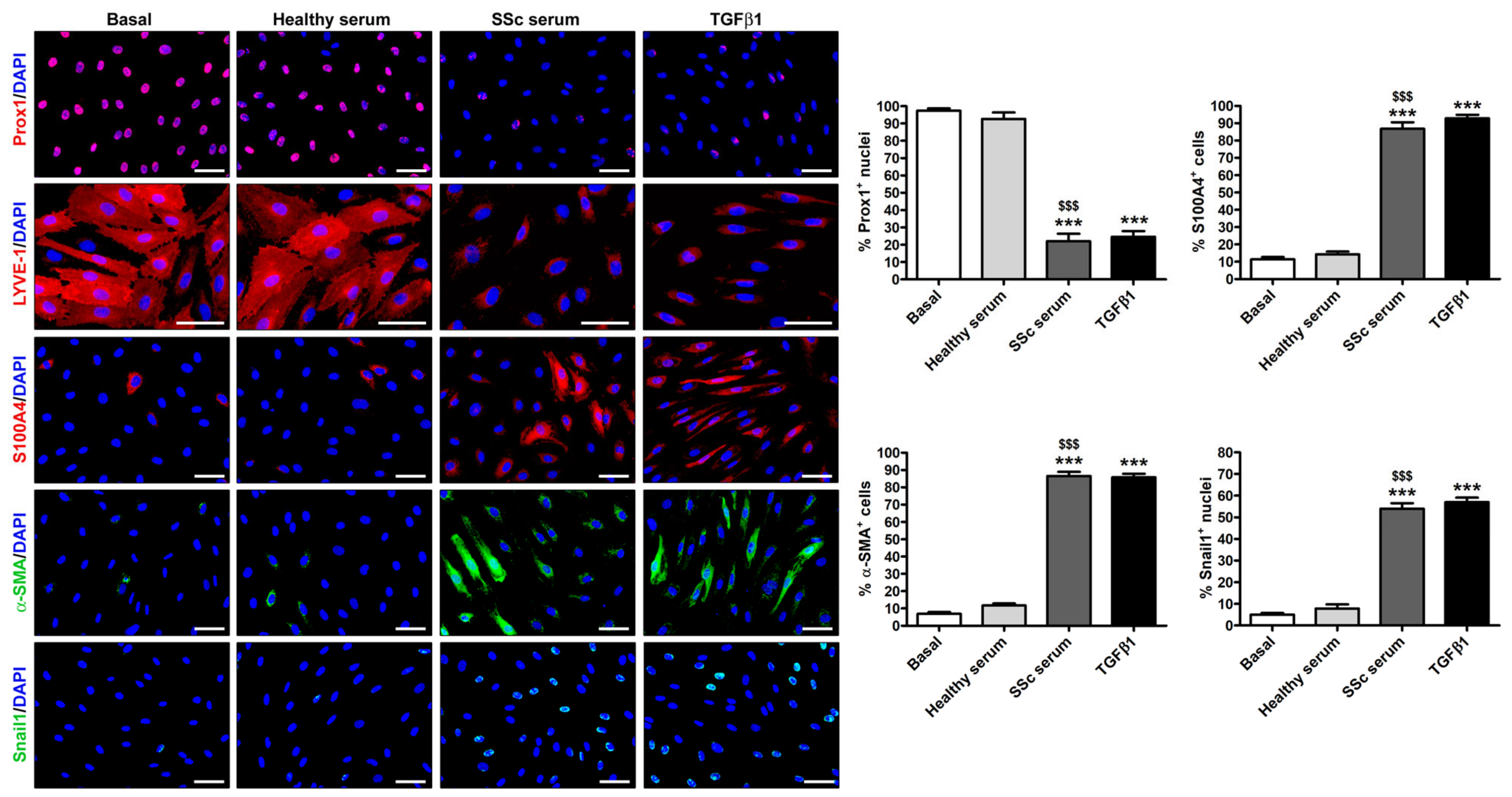
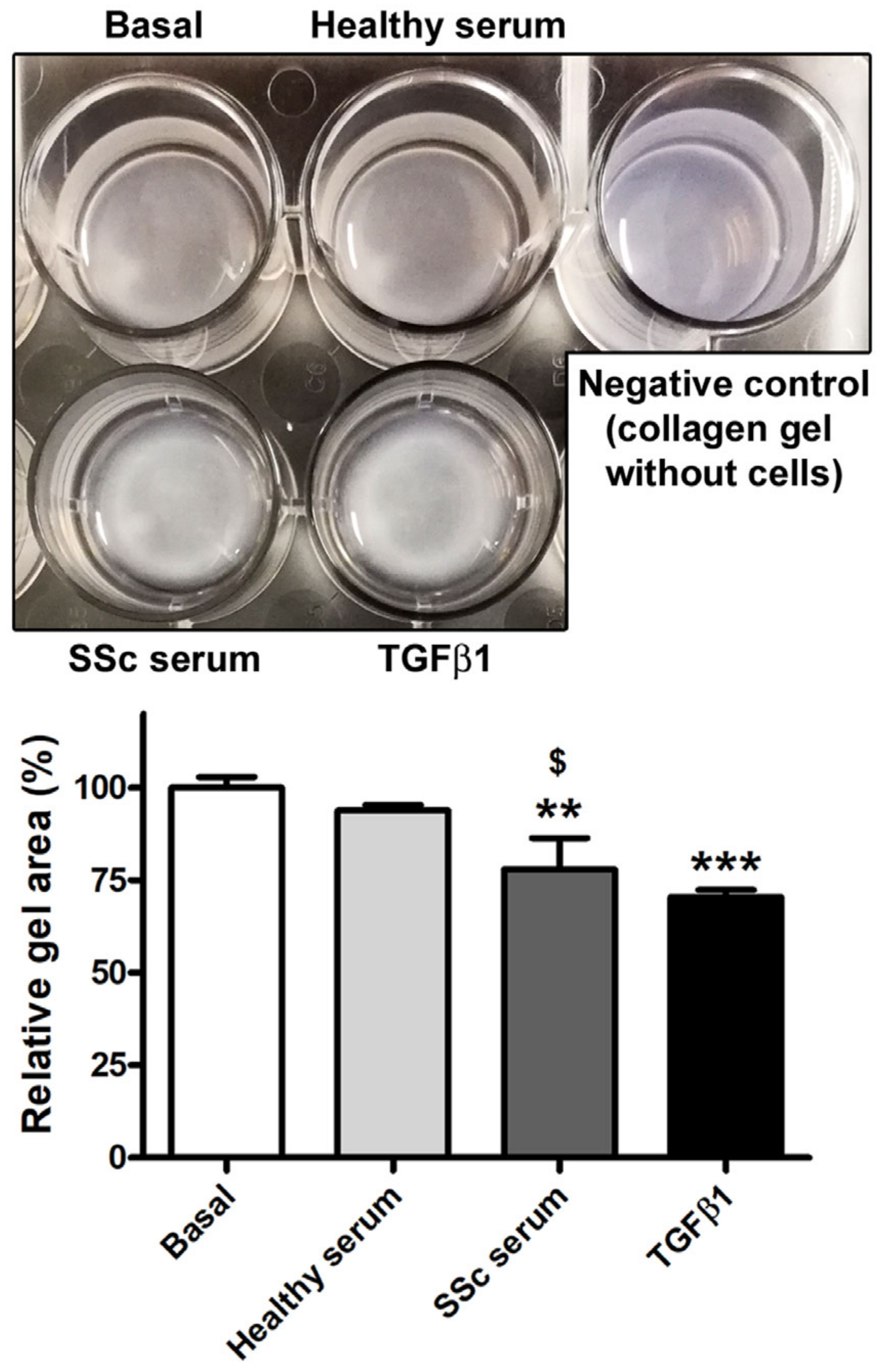
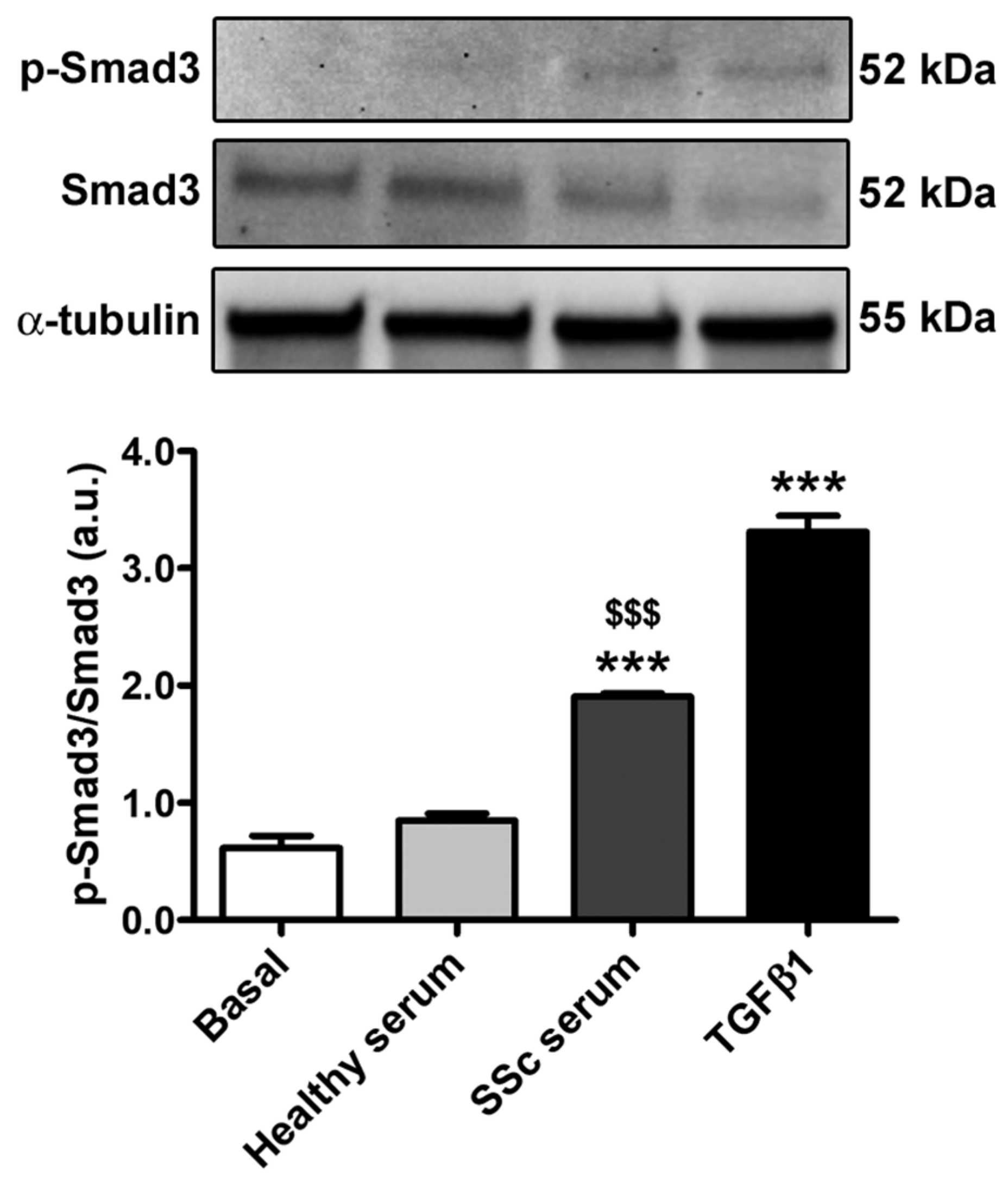
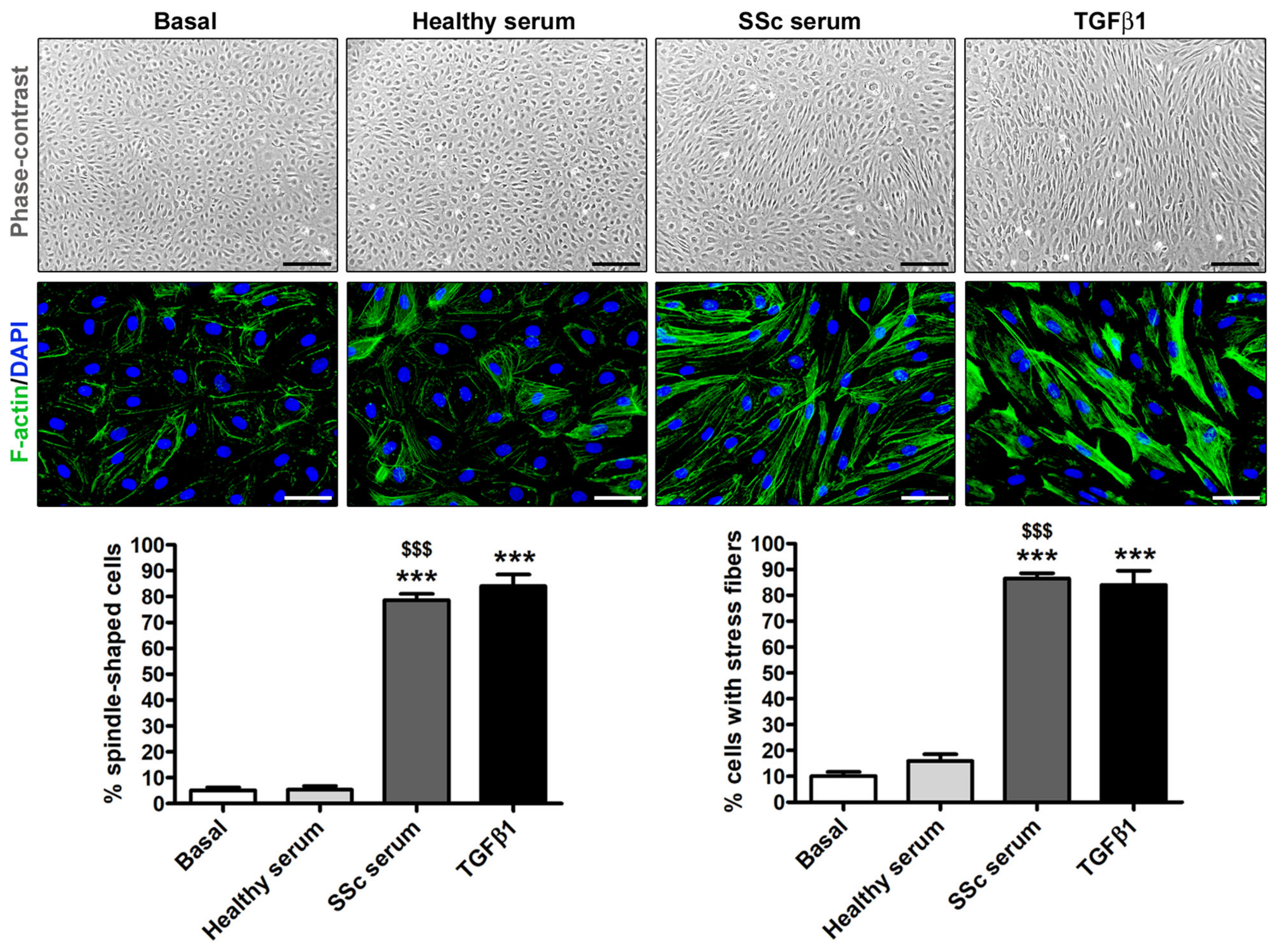
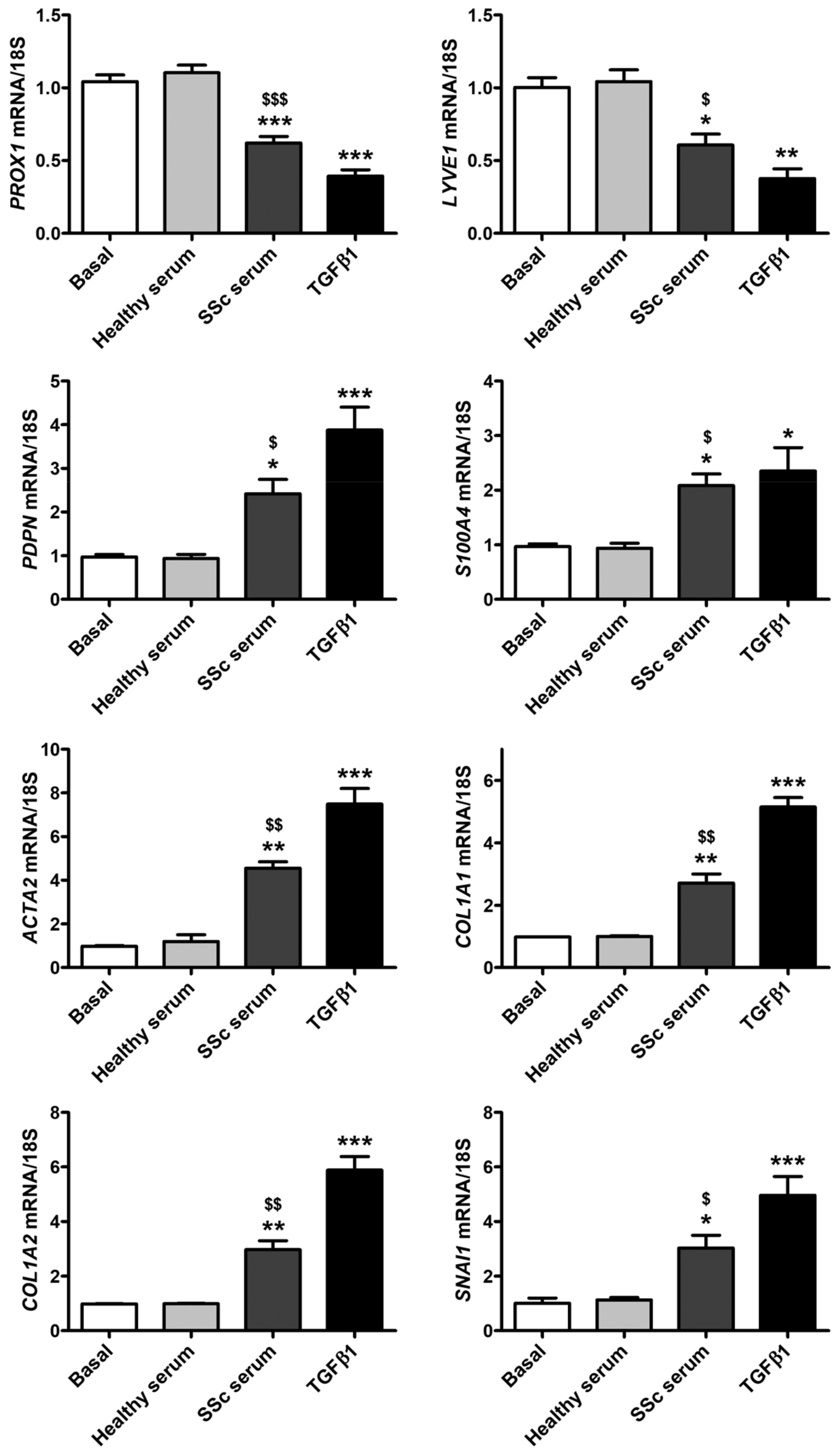
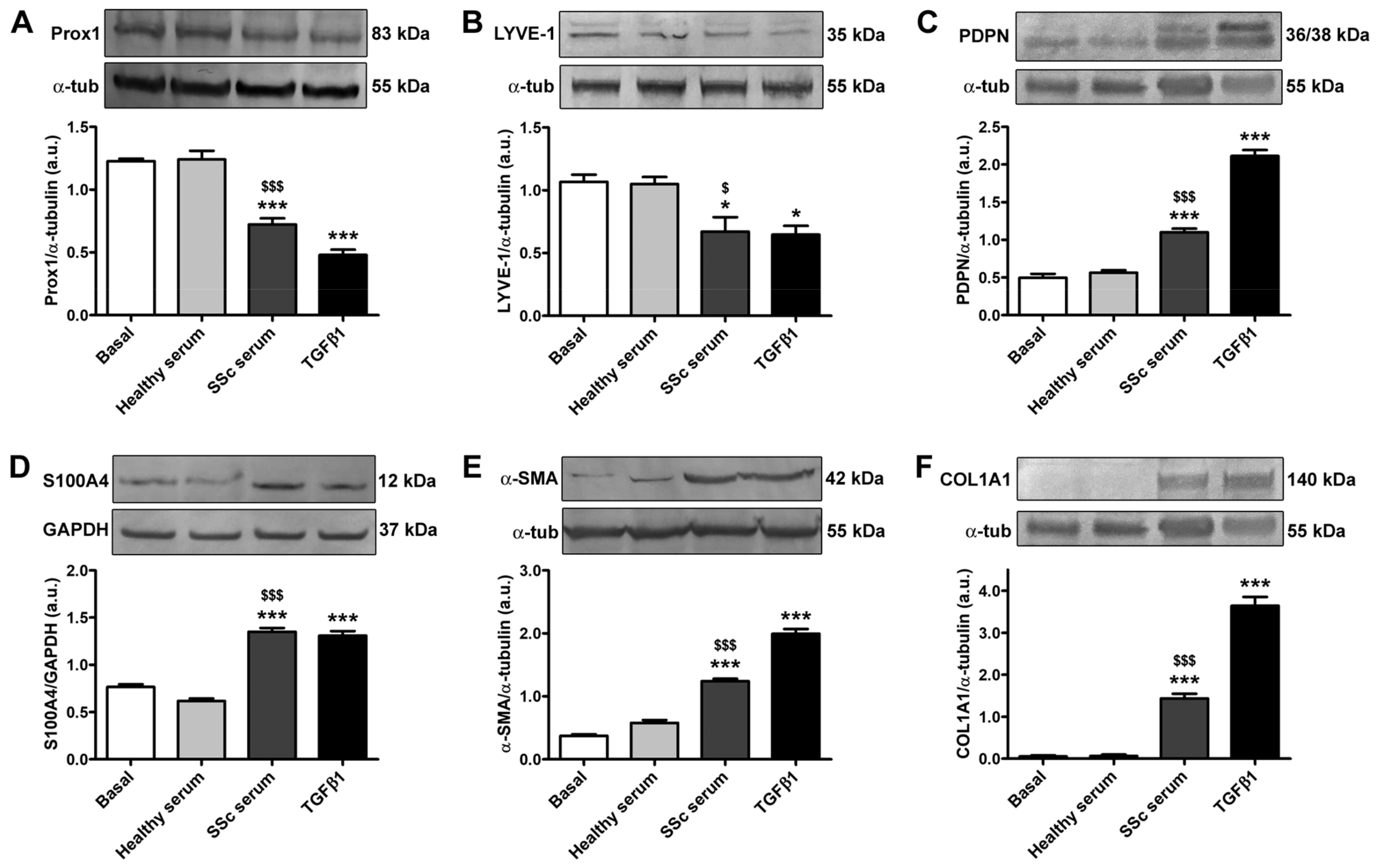
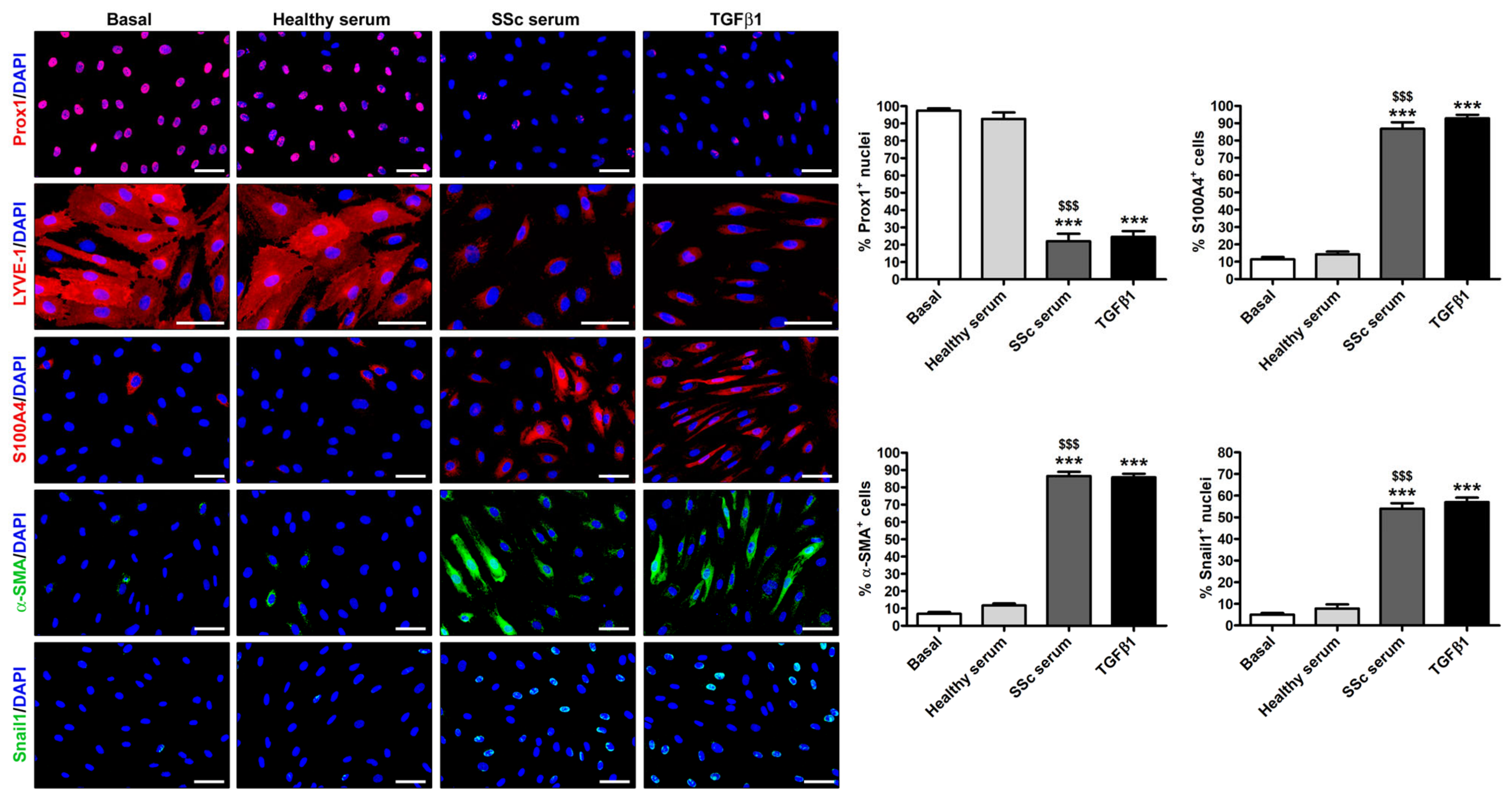
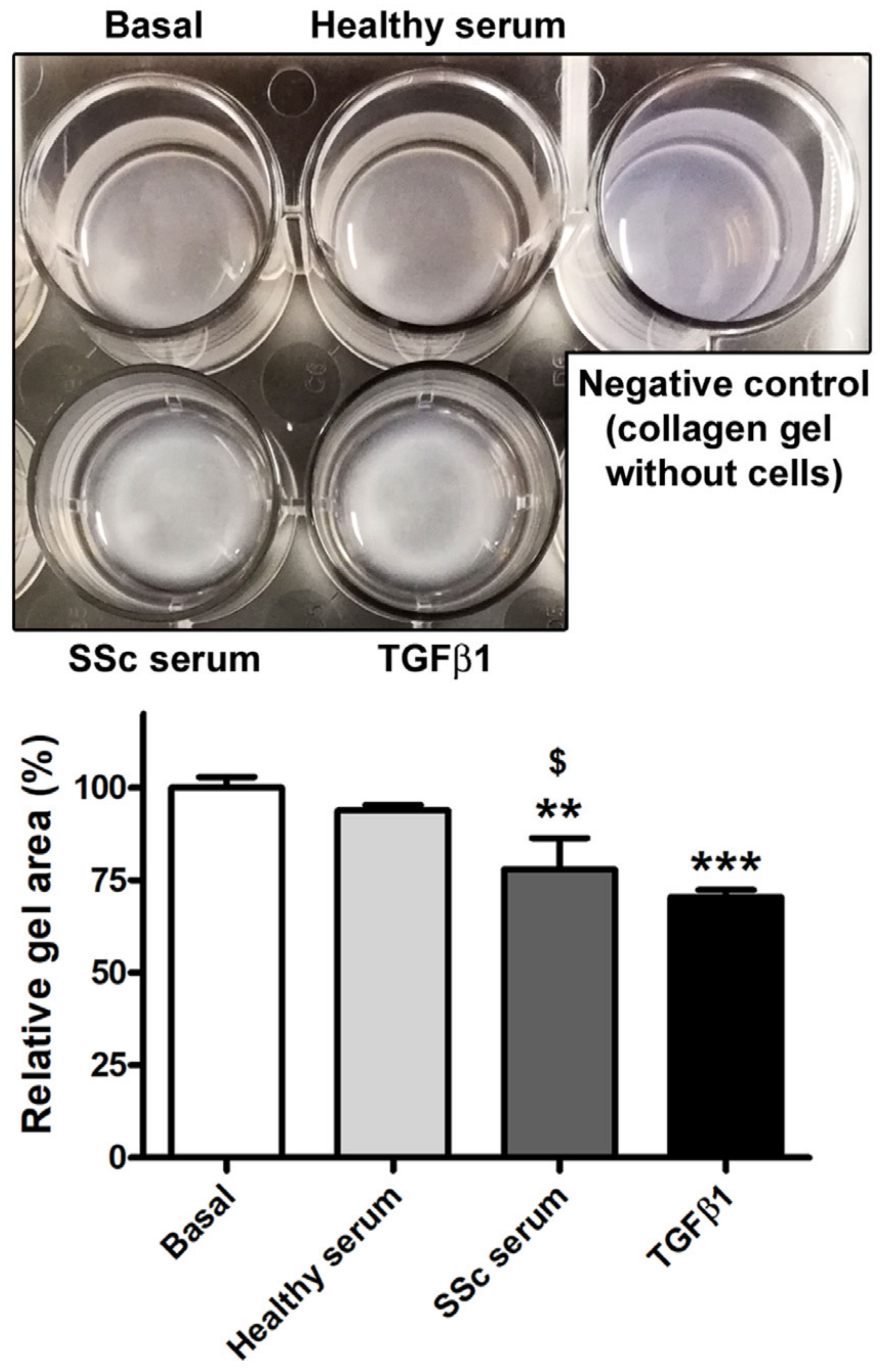
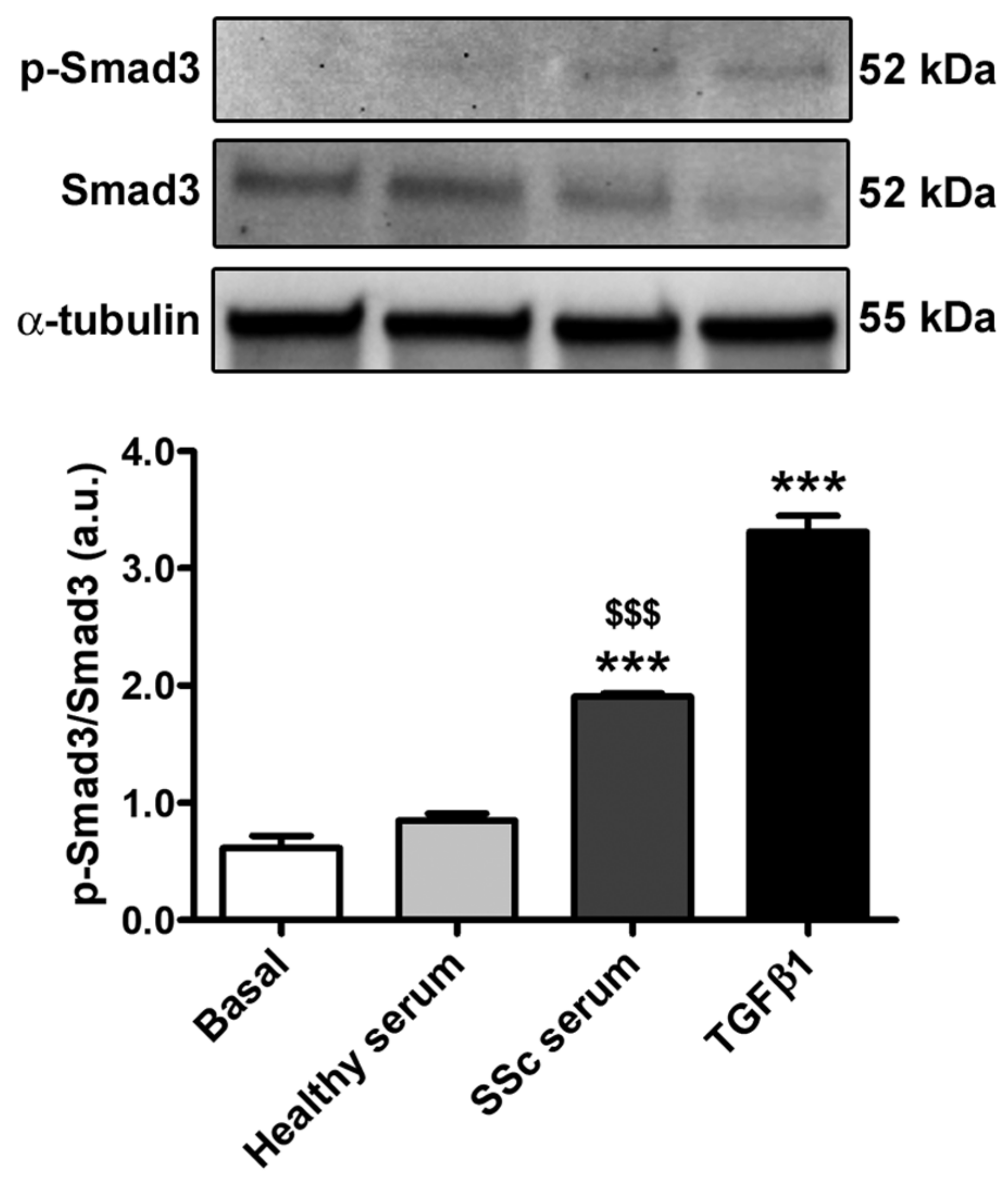
3.2. Treatment with SSc Serum Induces the Acquisition of a Myofibroblast-Like Profibrotic Phenotype in HdLy-MVECs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khanna, D.; Krieger, N.; Sullivan, K.M. Improving Outcomes in Scleroderma: Recent Progress of Cell-Based Therapies. Rheumatology 2023, 62, 2060–2069. [Google Scholar] [CrossRef] [PubMed]
- Asano, Y. The Pathogenesis of Systemic Sclerosis: An Understanding Based on a Common Pathologic Cascade across Multiple Organs and Additional Organ-Specific Pathologies. J. Clin. Med. 2020, 9, 2687. [Google Scholar] [CrossRef] [PubMed]
- Volkmann, E.R.; Andréasson, K.; Smith, V. Systemic Sclerosis. Lancet 2023, 401, 304–318. [Google Scholar] [CrossRef] [PubMed]
- Varga, J.; Trojanowska, M.; Kuwana, M. Pathogenesis of Systemic Sclerosis: Recent Insights of Molecular and Cellular Mechanisms and Therapeutic Opportunities. J. Scleroderma Relat. Disord. 2017, 2, 137–152. [Google Scholar] [CrossRef]
- Korman, B. Evolving Insights into the Cellular and Molecular Pathogenesis of Fibrosis in Systemic Sclerosis. Transl. Res. J. Lab. Clin. Med. 2019, 209, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Van Caam, A.; Vonk, M.; van den Hoogen, F.; van Lent, P.; van der Kraan, P. Unraveling SSc Pathophysiology; The Myofibroblast. Front. Immunol. 2018, 9, 2452. [Google Scholar] [CrossRef]
- Romano, E.; Rosa, I.; Fioretto, B.S.; Matucci-Cerinic, M.; Manetti, M. New Insights into Profibrotic Myofibroblast Formation in Systemic Sclerosis: When the Vascular Wall Becomes the Enemy. Life 2021, 11, 610. [Google Scholar] [CrossRef]
- Romano, E.; Rosa, I.; Fioretto, B.S.; Matucci-Cerinic, M.; Manetti, M. The Role of Pro-Fibrotic Myofibroblasts in Systemic Sclerosis: From Origin to Therapeutic Targeting. Curr. Mol. Med. 2022, 22, 209–239. [Google Scholar] [CrossRef]
- Garrett, S.M.; Baker Frost, D.; Feghali-Bostwick, C. The Mighty Fibroblast and Its Utility in Scleroderma Research. J. Scleroderma Relat. Disord. 2017, 2, 69–134. [Google Scholar] [CrossRef]
- Homps-Legrand, M.; Crestani, B.; Mailleux, A.A. Origins of Pathological Myofibroblasts in Lung Fibrosis: Insights from Lineage Tracing Mouse Models in the Single-Cell RNA Sequencing Era. Am. J. Physiol. Lung Cell. Mol. Physiol. 2023, 324, L737–L746. [Google Scholar] [CrossRef]
- Rosa, I.; Romano, E.; Fioretto, B.S.; Manetti, M. The Contribution of Mesenchymal Transitions to the Pathogenesis of Systemic Sclerosis. Eur. J. Rheumatol. 2020, 7, S157–S164. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Wang, L. Endothelial-to-Mesenchymal Transition in Tumour Progression and Its Potential Roles in Tumour Therapy. Ann. Med. 2023, 55, 1058–1069. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Nkennor, B.; Mastikhina, O.; Soon, K.; Nunes, S.S. Endothelium-Mediated Contributions to Fibrosis. Semin. Cell Dev. Biol. 2020, 101, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Piera-Velazquez, S.; Li, Z.; Jimenez, S.A. Role of Endothelial-Mesenchymal Transition (EndoMT) in the Pathogenesis of Fibrotic Disorders. Am. J. Pathol. 2011, 179, 1074–1080. [Google Scholar] [CrossRef]
- Jimenez, S.A.; Piera-Velazquez, S. Endothelial to Mesenchymal Transition (EndoMT) in the Pathogenesis of Systemic Sclerosis-Associated Pulmonary Fibrosis and Pulmonary Arterial Hypertension. Myth or Reality? Matrix Biol. J. Int. Soc. Matrix Biol. 2016, 51, 26–36. [Google Scholar] [CrossRef]
- Manetti, M.; Romano, E.; Rosa, I.; Guiducci, S.; Bellando-Randone, S.; De Paulis, A.; Ibba-Manneschi, L.; Matucci-Cerinic, M. Endothelial-to-Mesenchymal Transition Contributes to Endothelial Dysfunction and Dermal Fibrosis in Systemic Sclerosis. Ann. Rheum. Dis. 2017, 76, 924–934. [Google Scholar] [CrossRef]
- Good, R.B.; Gilbane, A.J.; Trinder, S.L.; Denton, C.P.; Coghlan, G.; Abraham, D.J.; Holmes, A.M. Endothelial to Mesenchymal Transition Contributes to Endothelial Dysfunction in Pulmonary Arterial Hypertension. Am. J. Pathol. 2015, 185, 1850–1858. [Google Scholar] [CrossRef]
- Mendoza, F.A.; Piera-Velazquez, S.; Farber, J.L.; Feghali-Bostwick, C.; Jiménez, S.A. Endothelial Cells Expressing Endothelial and Mesenchymal Cell Gene Products in Lung Tissue from Patients With Systemic Sclerosis-Associated Interstitial Lung Disease. Arthritis Rheumatol. 2016, 68, 210–217. [Google Scholar] [CrossRef]
- Akhmetshina, A.; Beer, J.; Zwerina, K.; Englbrecht, M.; Palumbo, K.; Dees, C.; Reich, N.; Zwerina, J.; Szucs, G.; Gusinde, J.; et al. Decreased Lymphatic Vessel Counts in Patients with Systemic Sclerosis: Association with Fingertip Ulcers. Arthritis Rheum. 2010, 62, 1513–1522. [Google Scholar] [CrossRef]
- Manetti, M.; Milia, A.F.; Guiducci, S.; Romano, E.; Matucci-Cerinic, M.; Ibba-Manneschi, L. Progressive Loss of Lymphatic Vessels in Skin of Patients with Systemic Sclerosis. J. Rheumatol. 2011, 38, 297–301. [Google Scholar] [CrossRef]
- Chen, J.-M.; Luo, B.; Ma, R.; Luo, X.-X.; Chen, Y.-S.; Li, Y. Lymphatic Endothelial Markers and Tumor Lymphangiogenesis Assessment in Human Breast Cancer. Diagnostics 2021, 12, 4. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.-L.; Yang, N.-Z.; Shi, L.-H.; Zhao, G.-H.; Zhou, W.; Ding, Q.; Wang, M.-H.; Zhang, Y.-S. The Optimum Marker for the Detection of Lymphatic Vessels. Mol. Clin. Oncol. 2017, 7, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J. Cell Fate Determination of Lymphatic Endothelial Cells. Int. J. Mol. Sci. 2020, 21, 4790. [Google Scholar] [CrossRef] [PubMed]
- Povýšil, C.; Kaňa, M.; Zámečník, L.; Vaľová, Z.; Hanuš, T. Podoplanin (D2-40) Is a Reliable Marker of Urinary Bladder Myofibroblasts (Telocytes). Folia Biol. 2014, 60, 286–289. [Google Scholar]
- Nazari, B.; Rice, L.M.; Stifano, G.; Barron, A.M.S.; Wang, Y.M.; Korndorf, T.; Lee, J.; Bhawan, J.; Lafyatis, R.; Browning, J.L. Altered Dermal Fibroblasts in Systemic Sclerosis Display Podoplanin and CD90. Am. J. Pathol. 2016, 186, 2650–2664. [Google Scholar] [CrossRef]
- Suchanski, J.; Tejchman, A.; Zacharski, M.; Piotrowska, A.; Grzegrzolka, J.; Chodaczek, G.; Nowinska, K.; Rys, J.; Dziegiel, P.; Kieda, C.; et al. Podoplanin Increases the Migration of Human Fibroblasts and Affects the Endothelial Cell Network Formation: A Possible Role for Cancer-Associated Fibroblasts in Breast Cancer Progression. PLoS ONE 2017, 12, e0184970. [Google Scholar] [CrossRef]
- Cimini, M.; Kishore, R. Role of Podoplanin-Positive Cells in Cardiac Fibrosis and Angiogenesis After Ischemia. Front. Physiol. 2021, 12, 667278. [Google Scholar] [CrossRef]
- Kondo, R.; Sakamoto, N.; Harada, K.; Hashimoto, H.; Morisue, R.; Yanagihara, K.; Kinoshita, T.; Kojima, M.; Ishii, G. Cancer-Associated Fibroblast-Dependent and -Independent Invasion of Gastric Cancer Cells. J. Cancer Res. Clin. Oncol. 2022, 149, 5309–5319. [Google Scholar] [CrossRef]
- Tian, Y.; Chen, X.; Wang, X.; Song, Y. Podoplanin Promotes the Carcinogenicity of Gastric Cancer by Activating Ezrin and Mediating the Crosstalk between Tumour Cells and Cancer-Associated Fibroblasts. Exp. Physiol. 2023, 108, 740–751. [Google Scholar] [CrossRef]
- Ichise, T.; Yoshida, N.; Ichise, H. FGF2-Induced Ras-MAPK Signalling Maintains Lymphatic Endothelial Cell Identity by Upregulating Endothelial-Cell-Specific Gene Expression and Suppressing TGFβ Signalling through Smad2. J. Cell Sci. 2014, 127, 845–857. [Google Scholar] [CrossRef]
- Van den Hoogen, F.; Khanna, D.; Fransen, J.; Johnson, S.R.; Baron, M.; Tyndall, A.; Matucci-Cerinic, M.; Naden, R.P.; Medsger, T.A.; Carreira, P.E.; et al. 2013 Classification Criteria for Systemic Sclerosis: An American College of Rheumatology/European League against Rheumatism Collaborative Initiative. Arthritis Rheum. 2013, 65, 2737–2747. [Google Scholar] [CrossRef] [PubMed]
- Avraham, T.; Yan, A.; Zampell, J.C.; Daluvoy, S.V.; Haimovitz-Friedman, A.; Cordeiro, A.P.; Mehrara, B.J. Radiation Therapy Causes Loss of Dermal Lymphatic Vessels and Interferes with Lymphatic Function by TGF-Β1-Mediated Tissue Fibrosis. Am. J. Physiol. Cell Physiol. 2010, 299, C589–C605. [Google Scholar] [CrossRef] [PubMed]
- Itoh, F.; Watabe, T. TGF-β Signaling in Lymphatic Vascular Vessel Formation and Maintenance. Front. Physiol. 2022, 13, 1081376. [Google Scholar] [CrossRef] [PubMed]
- Yoshimatsu, Y.; Watabe, T. Emerging Roles of Inflammation-Mediated Endothelial–Mesenchymal Transition in Health and Disease. Inflamm. Regen. 2022, 42, 9. [Google Scholar] [CrossRef]
- Frommer, M.L.; Langridge, B.J.; Awad, L.; Jasionowska, S.; Denton, C.P.; Abraham, D.J.; Abu-Hanna, J.; Butler, P.E.M. Single-Cell Analysis of ADSC Interactions with Fibroblasts and Endothelial Cells in Scleroderma Skin. Cells 2023, 12, 1784. [Google Scholar] [CrossRef]
- Bruni, C.; Frech, T.; Manetti, M.; Rossi, F.W.; Furst, D.E.; De Paulis, A.; Rivellese, F.; Guiducci, S.; Matucci-Cerinic, M.; Bellando-Randone, S. Vascular Leaking, a Pivotal and Early Pathogenetic Event in Systemic Sclerosis: Should the Door Be Closed? Front. Immunol. 2018, 9, 2045. [Google Scholar] [CrossRef]
- Rossi, A.; Sozio, F.; Sestini, P.; Renzoni, E.A.; Khan, K.; Denton, C.P.; Abraham, D.J.; Weber, E. Lymphatic and Blood Vessels in Scleroderma Skin, a Morphometric Analysis. Hum. Pathol. 2010, 41, 366–374. [Google Scholar] [CrossRef]
- Czarnowska, E.; Ratajska, A.; Jankowska-Steifer, E.; Flaht-Zabost, A.; Niderla-Bielińska, J. Extracellular Matrix Molecules Associated with Lymphatic Vessels in Health and Disease. Histol. Histopathol. 2023. Epub ahead of print. [Google Scholar] [CrossRef]
- Peluzzo, A.M.; Bkhache, M.; Do, L.N.H.; Autieri, M.V.; Liu, X. Differential Regulation of Lymphatic Junctional Morphology and the Potential Effects on Cardiovascular Diseases. Front. Physiol. 2023, 14, 1198052. [Google Scholar] [CrossRef]
- Manetti, M.; Romano, E.; Rosa, I.; Fioretto, B.S.; Guiducci, S.; Bellando-Randone, S.; Pigatto, E.; Cozzi, F.; Ibba-Manneschi, L.; Matucci-Cerinic, M. Systemic Sclerosis Serum Significantly Impairs the Multi-Step Lymphangiogenic Process: In Vitro Evidence. Int. J. Mol. Sci. 2019, 20, 6189. [Google Scholar] [CrossRef]
- Shiiya, T.; Hirashima, M. From Lymphatic Endothelial Cell Migration to Formation of Tubular Lymphatic Vascular Network. Front. Physiol. 2023, 14, 1124696. [Google Scholar] [CrossRef] [PubMed]
- Manetti, M.; Pratesi, S.; Romano, E.; Rosa, I.; Bruni, C.; Bellando-Randone, S.; Guiducci, S.; Maggi, E.; Ibba-Manneschi, L.; Matucci-Cerinic, M. Decreased Circulating Lymphatic Endothelial Progenitor Cells in Digital Ulcer-Complicated Systemic Sclerosis. Ann. Rheum. Dis. 2019, 78, 575–577. [Google Scholar] [CrossRef] [PubMed]
- Kajiya, K.; Kunstfeld, R.; Detmar, M.; Chung, J.H. Reduction of Lymphatic Vessels in Photodamaged Human Skin. J. Dermatol. Sci. 2007, 47, 241–243. [Google Scholar] [CrossRef] [PubMed]
- Karaman, S.; Buschle, D.; Luciani, P.; Leroux, J.-C.; Detmar, M.; Proulx, S.T. Decline of Lymphatic Vessel Density and Function in Murine Skin during Aging. Angiogenesis 2015, 18, 489–498. [Google Scholar] [CrossRef]
- Cheng, F.; Pekkonen, P.; Laurinavicius, S.; Sugiyama, N.; Henderson, S.; Günther, T.; Rantanen, V.; Kaivanto, E.; Aavikko, M.; Sarek, G.; et al. KSHV-Initiated Notch Activation Leads to Membrane-Type-1 Matrix Metalloproteinase-Dependent Lymphatic Endothelial-to-Mesenchymal Transition. Cell Host Microbe 2011, 10, 577–590. [Google Scholar] [CrossRef]
- Cai, X.; Zhang, W.; Chen, G.; Li, R.-F.; Sun, Y.-F.; Zhao, Y.-F. Mesenchymal Status of Lymphatic Endothelial Cell: Enlightening Treatment of Lymphatic Malformation. Int. J. Clin. Exp. Med. 2015, 8, 12239–12251. [Google Scholar]
- Yoshimatsu, Y.; Kimuro, S.; Pauty, J.; Takagaki, K.; Nomiyama, S.; Inagawa, A.; Maeda, K.; Podyma-Inoue, K.A.; Kajiya, K.; Matsunaga, Y.T.; et al. TGF-Beta and TNF-Alpha Cooperatively Induce Mesenchymal Transition of Lymphatic Endothelial Cells via Activation of Activin Signals. PLoS ONE 2020, 15, e0232356. [Google Scholar] [CrossRef]
- Kinashi, H.; Ito, Y.; Sun, T.; Katsuno, T.; Takei, Y. Roles of the TGF-β–VEGF-C Pathway in Fibrosis-Related Lymphangiogenesis. Int. J. Mol. Sci. 2018, 19, 2487. [Google Scholar] [CrossRef]
- Evans, R.; Flores-Borja, F.; Nassiri, S.; Miranda, E.; Lawler, K.; Grigoriadis, A.; Monypenny, J.; Gillet, C.; Owen, J.; Gordon, P.; et al. Integrin-Mediated Macrophage Adhesion Promotes Lymphovascular Dissemination in Breast Cancer. Cell Rep. 2019, 27, 1967–1978.e4. [Google Scholar] [CrossRef]
- Scherer, S.D.; Bauer, J.; Schmaus, A.; Neumaier, C.; Herskind, C.; Veldwijk, M.R.; Wenz, F.; Sleeman, J.P. TGF-Β1 Is Present at High Levels in Wound Fluid from Breast Cancer Patients Immediately Post-Surgery, and Is Not Increased by Intraoperative Radiation Therapy (IORT). PLoS ONE 2016, 11, e0162221. [Google Scholar] [CrossRef]
- Chakraborty, S.; Zawieja, D.C.; Davis, M.J.; Muthuchamy, M. MicroRNA Signature of Inflamed Lymphatic Endothelium and Role of MiR-9 in Lymphangiogenesis and Inflammation. Am. J. Physiol. Cell Physiol. 2015, 309, C680–C692. [Google Scholar] [CrossRef] [PubMed]
- Moskalik, A.; Ratajska, A.; Majchrzak, B.; Jankowska-Steifer, E.; Bartkowiak, K.; Bartkowiak, M.; Niderla-Bielińska, J. MiR-31-5p-Modified RAW 264.7 Macrophages Affect Profibrotic Phenotype of Lymphatic Endothelial Cells In Vitro. Int. J. Mol. Sci. 2022, 23, 13193. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Zuo, X.X.; Li, Y.S.; Gao, S.M.; Dai, X.D.; Zhu, H.L.; Luo, H. Integration of MicroRNA and MRNA Expression Profiles in the Skin of Systemic Sclerosis Patients. Sci. Rep. 2017, 7, 42899. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-X.; Li, F.; Liu, D.; Sun, Y.-Y.; Zhao, Q.-H.; Jiang, R.; Wang, L.; Yuan, P.; Liu, J.-M.; Wu, Y.; et al. The Expression Profiling of MicroRNA in Systemic Sclerosis-Associated Pulmonary Arterial Hypertension. Ann. Transl. Med. 2021, 9, 1458. [Google Scholar] [CrossRef]
- Maciejewska, M.; Sikora, M.; Stec, A.; Zaremba, M.; Maciejewski, C.; Pawlik, K.; Rudnicka, L. Hypoxia-Inducible Factor-1α (HIF-1α) as a Biomarker for Changes in Microcirculation in Individuals with Systemic Sclerosis. Dermatol. Ther. 2023, 13, 1549–1560. [Google Scholar] [CrossRef]
- Becker, J.; Schwoch, S.; Zelent, C.; Sitte, M.; Salinas, G.; Wilting, J. Transcriptome Analysis of Hypoxic Lymphatic Endothelial Cells Indicates Their Potential to Contribute to Extracellular Matrix Rearrangement. Cells 2021, 10, 1008. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Assay ID | Cat# |
|---|---|---|
| PROX1 | Hs_PROX1_1_SG | QT01006670 |
| LYVE1 | Hs_LYVE1_1_SG | QT00034566 |
| PDPN | Hs_PDPN_1_SG | QT01015084 |
| S100A4 | Hs_S100A4_1_SG | QT00014259 |
| ACTA2 | Hs_ACTA2_1_SG | QT00088102 |
| COL1A1 | Hs_COL1A1_1_SG | QT00037793 |
| COL1A2 | Hs_COL1A2_1_SG | QT00072058 |
| SNAI1 | Hs_SNAI1_1_SG | QT00010010 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosa, I.; Romano, E.; Fioretto, B.S.; El Aoufy, K.; Bellando-Randone, S.; Matucci-Cerinic, M.; Manetti, M. Lymphatic Endothelial-to-Myofibroblast Transition: A Potential New Mechanism Underlying Skin Fibrosis in Systemic Sclerosis. Cells 2023, 12, 2195. https://doi.org/10.3390/cells12172195
Rosa I, Romano E, Fioretto BS, El Aoufy K, Bellando-Randone S, Matucci-Cerinic M, Manetti M. Lymphatic Endothelial-to-Myofibroblast Transition: A Potential New Mechanism Underlying Skin Fibrosis in Systemic Sclerosis. Cells. 2023; 12(17):2195. https://doi.org/10.3390/cells12172195
Chicago/Turabian StyleRosa, Irene, Eloisa Romano, Bianca Saveria Fioretto, Khadija El Aoufy, Silvia Bellando-Randone, Marco Matucci-Cerinic, and Mirko Manetti. 2023. "Lymphatic Endothelial-to-Myofibroblast Transition: A Potential New Mechanism Underlying Skin Fibrosis in Systemic Sclerosis" Cells 12, no. 17: 2195. https://doi.org/10.3390/cells12172195
APA StyleRosa, I., Romano, E., Fioretto, B. S., El Aoufy, K., Bellando-Randone, S., Matucci-Cerinic, M., & Manetti, M. (2023). Lymphatic Endothelial-to-Myofibroblast Transition: A Potential New Mechanism Underlying Skin Fibrosis in Systemic Sclerosis. Cells, 12(17), 2195. https://doi.org/10.3390/cells12172195