

Intensive Care Unit Acquired Weakness Is Associated with Rapid Changes to Skeletal Muscle Proteostasis

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
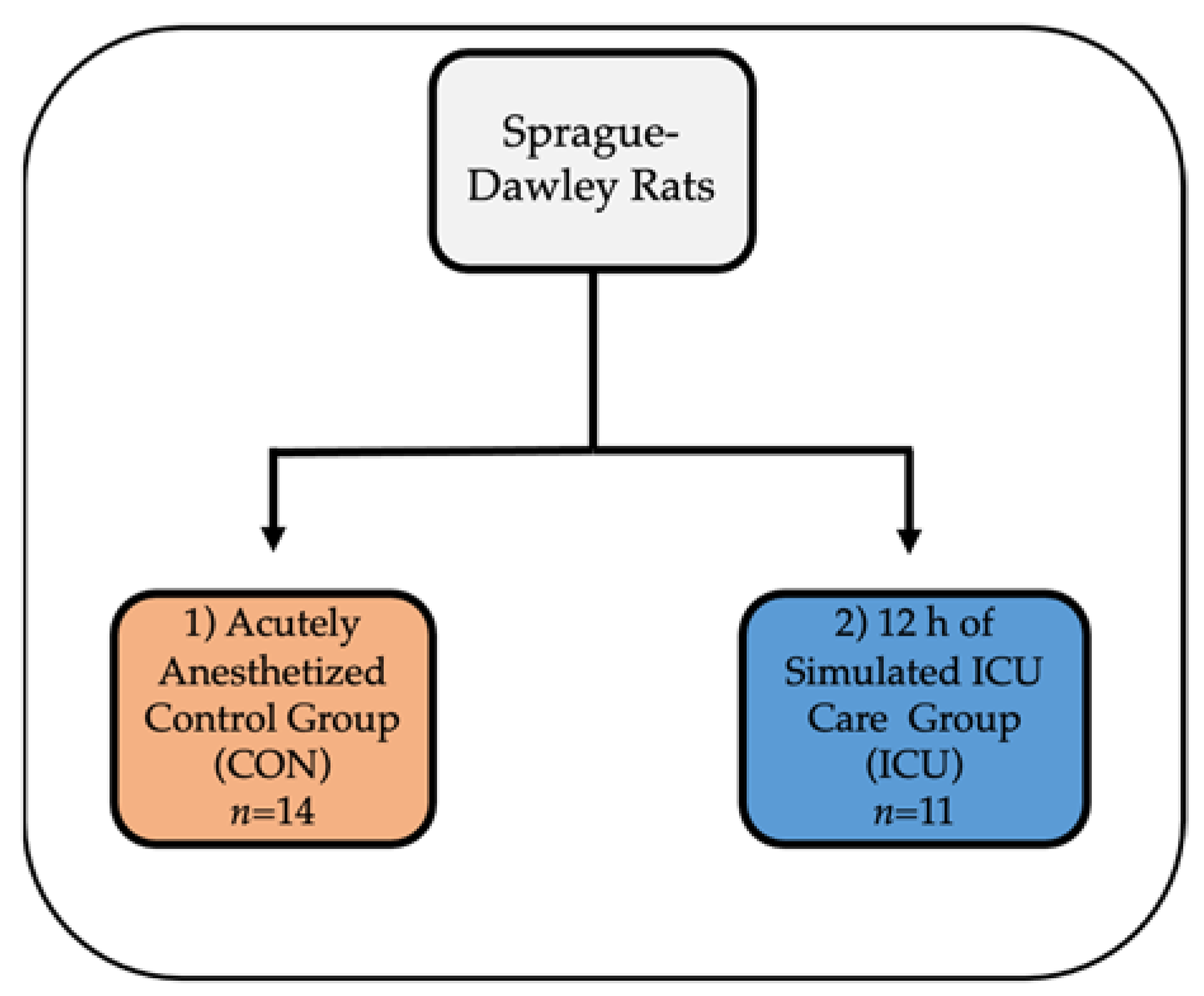
2.1. Animals, Institutional Approval and Experimental Design
2.2. Simulated ICU Protocol
2.3. Skeletal Muscle Fiber Cross-Sectional Area
2.4. Western Blot Analysis
2.5. Real-Time Polymerase Chain Reaction
2.6. Statistical Analysis
3. Results
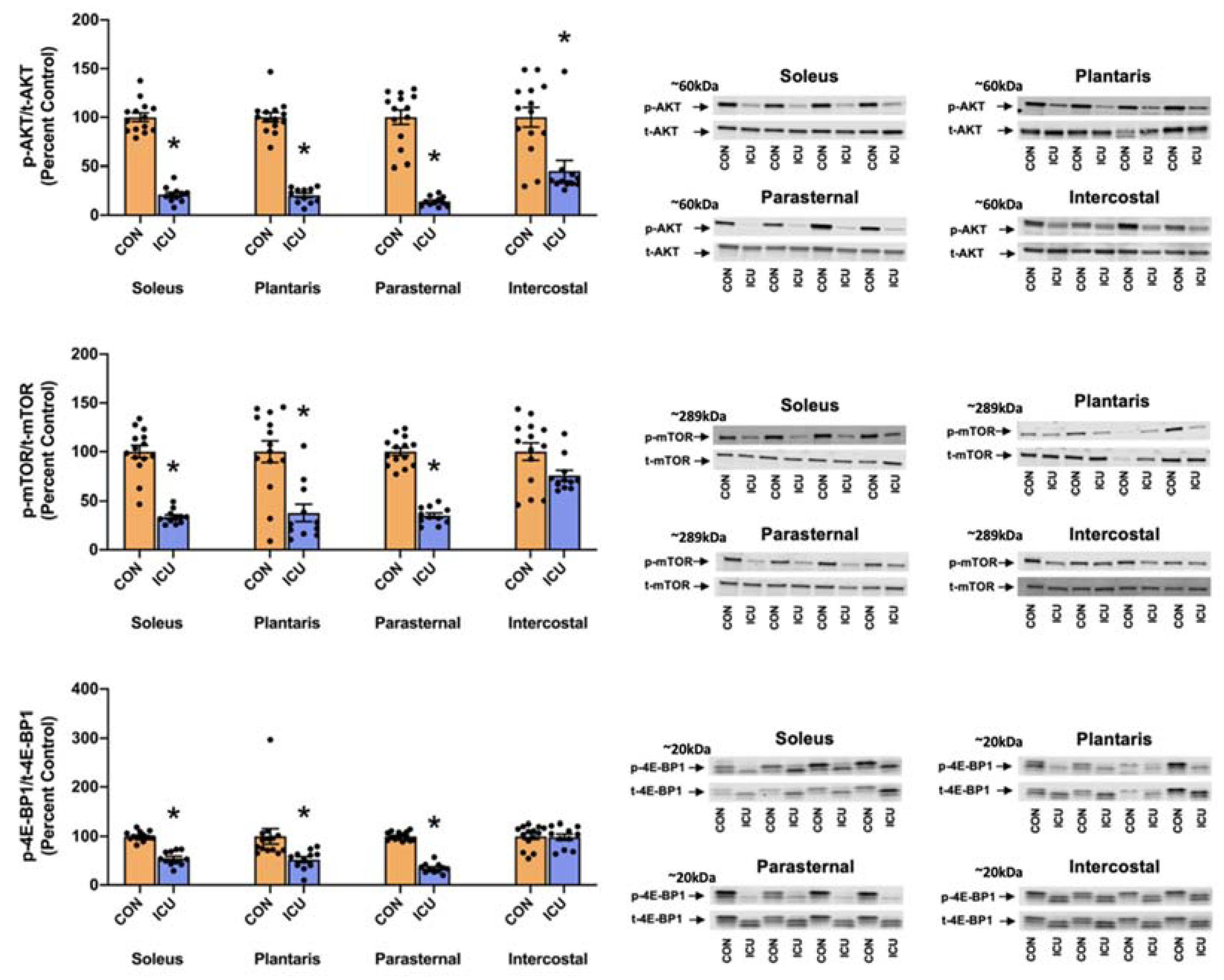
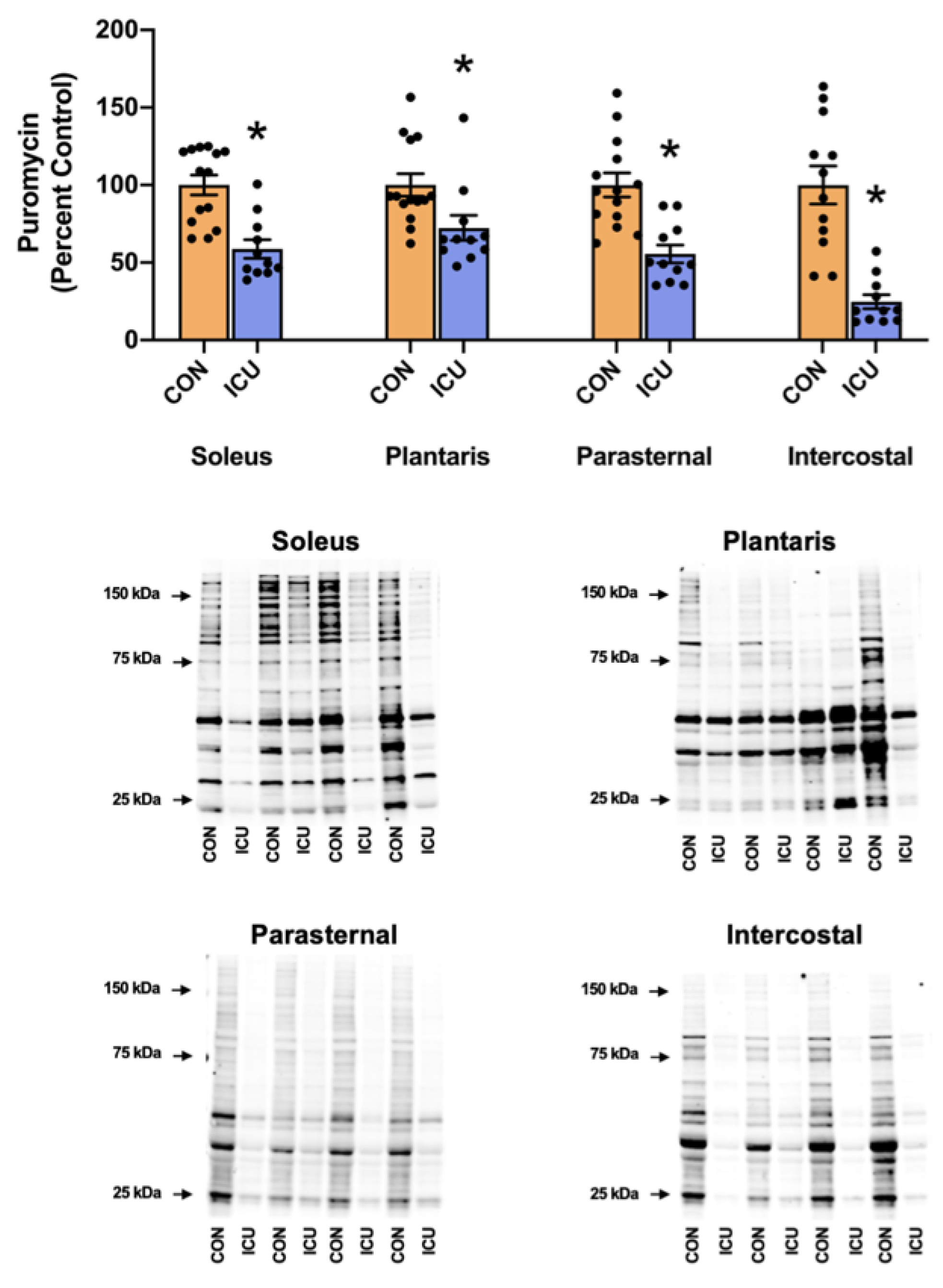
3.1. Simulated ICU Care Depresses Skeletal Muscle Protein Synthesis and Signaling
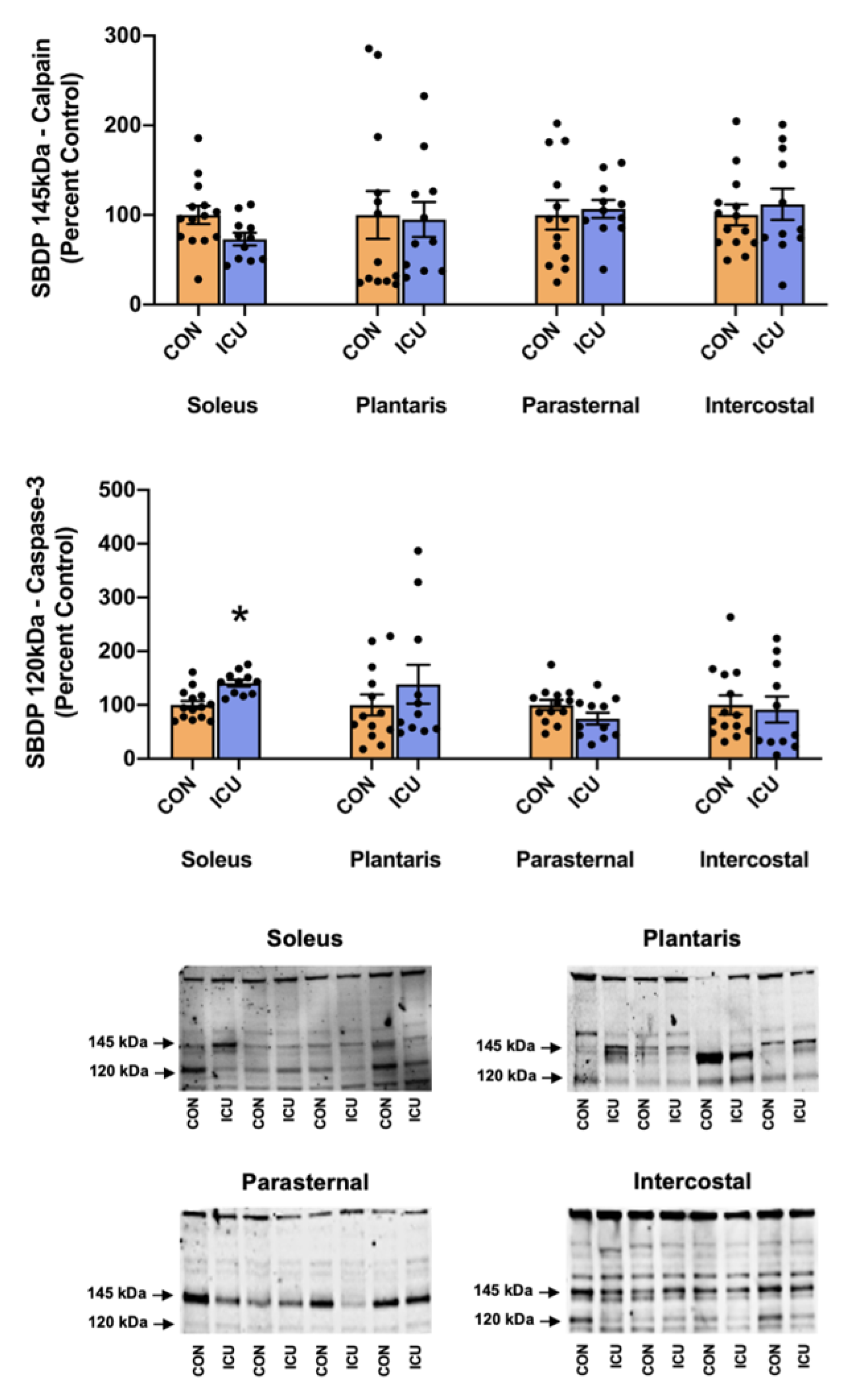
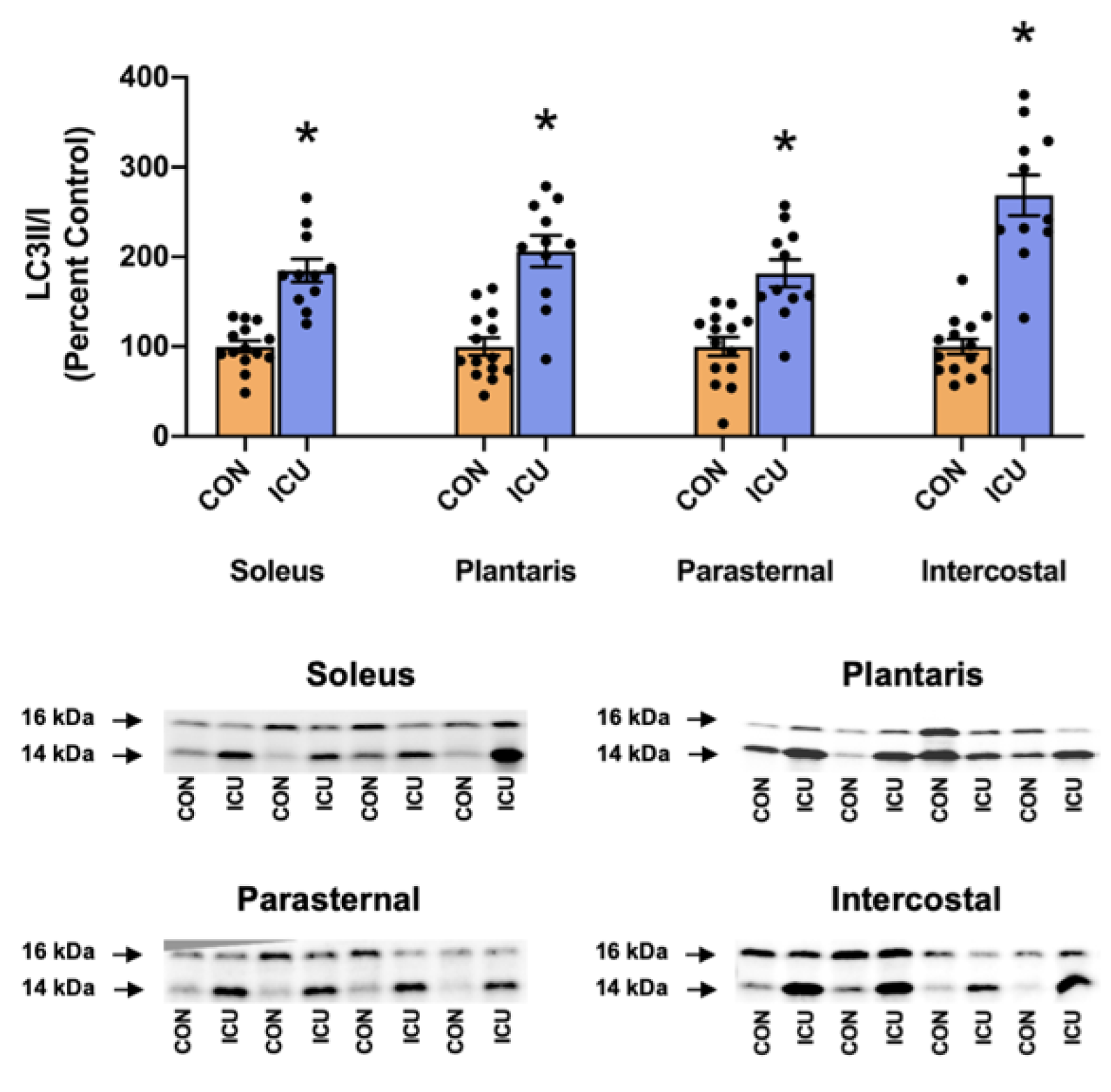
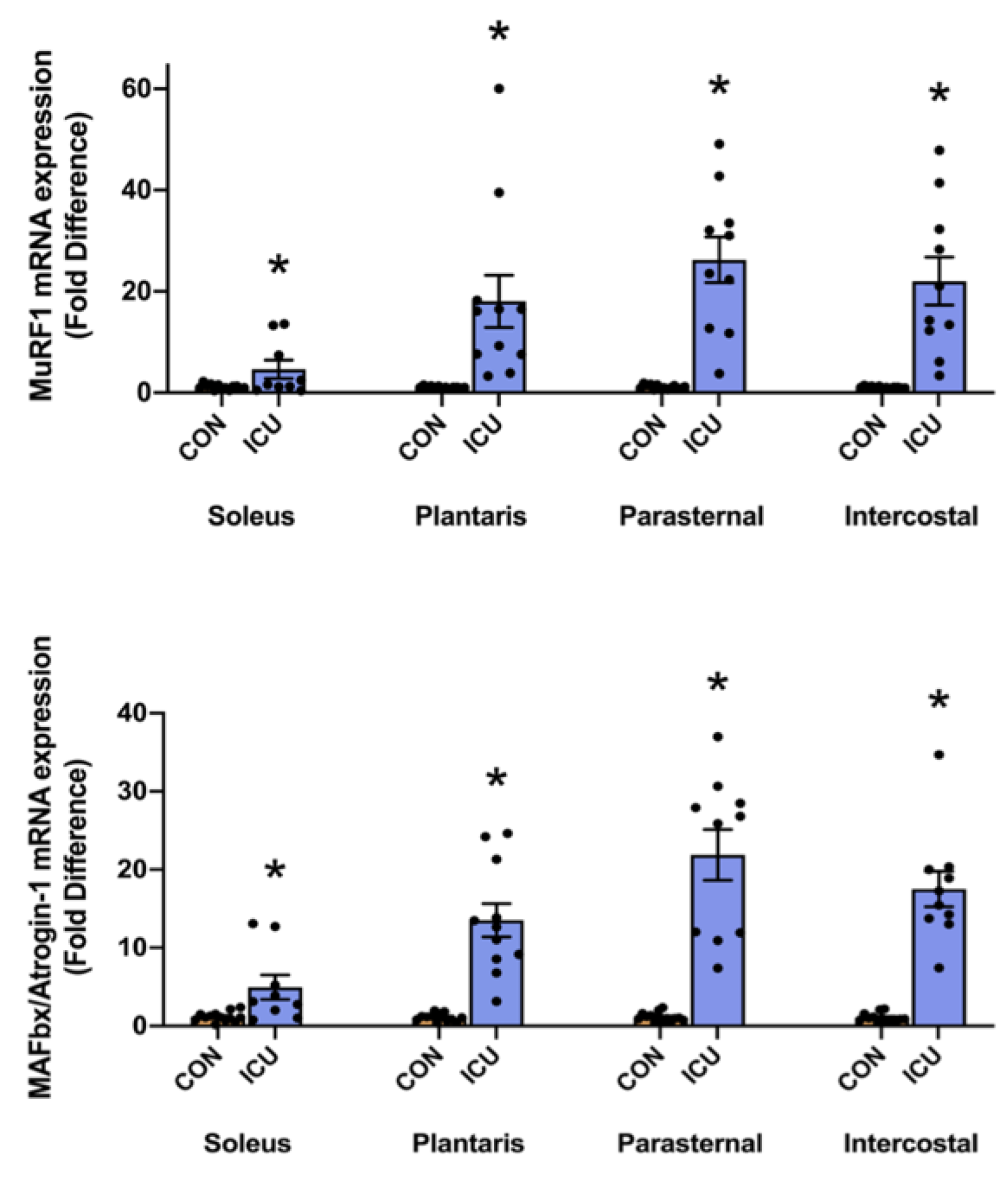
3.2. Simulated ICU Care Rapidly Increases Biomarkers of Skeletal Muscle Proteolysis
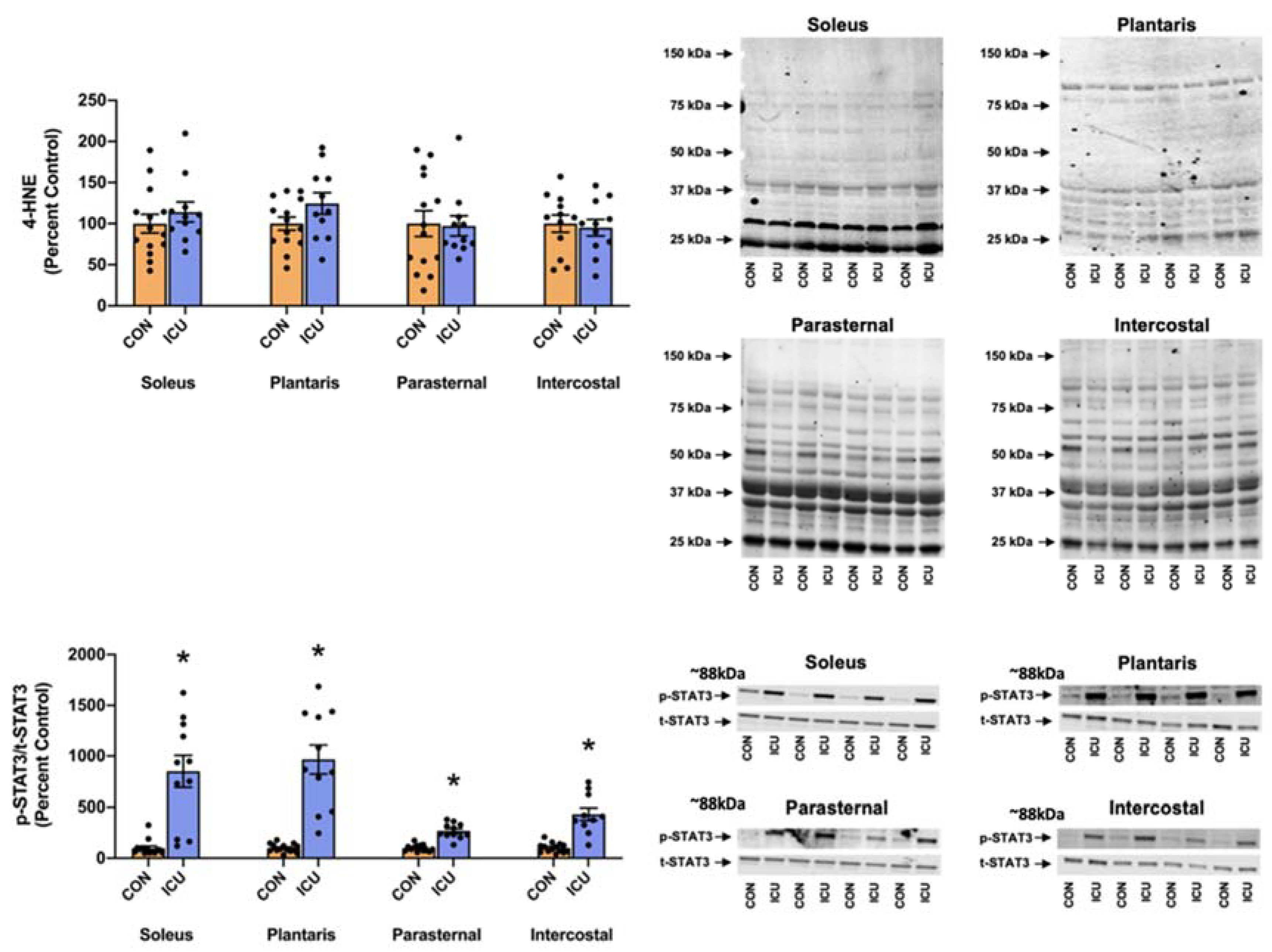
3.3. ICU-Induced Activation of STAT3
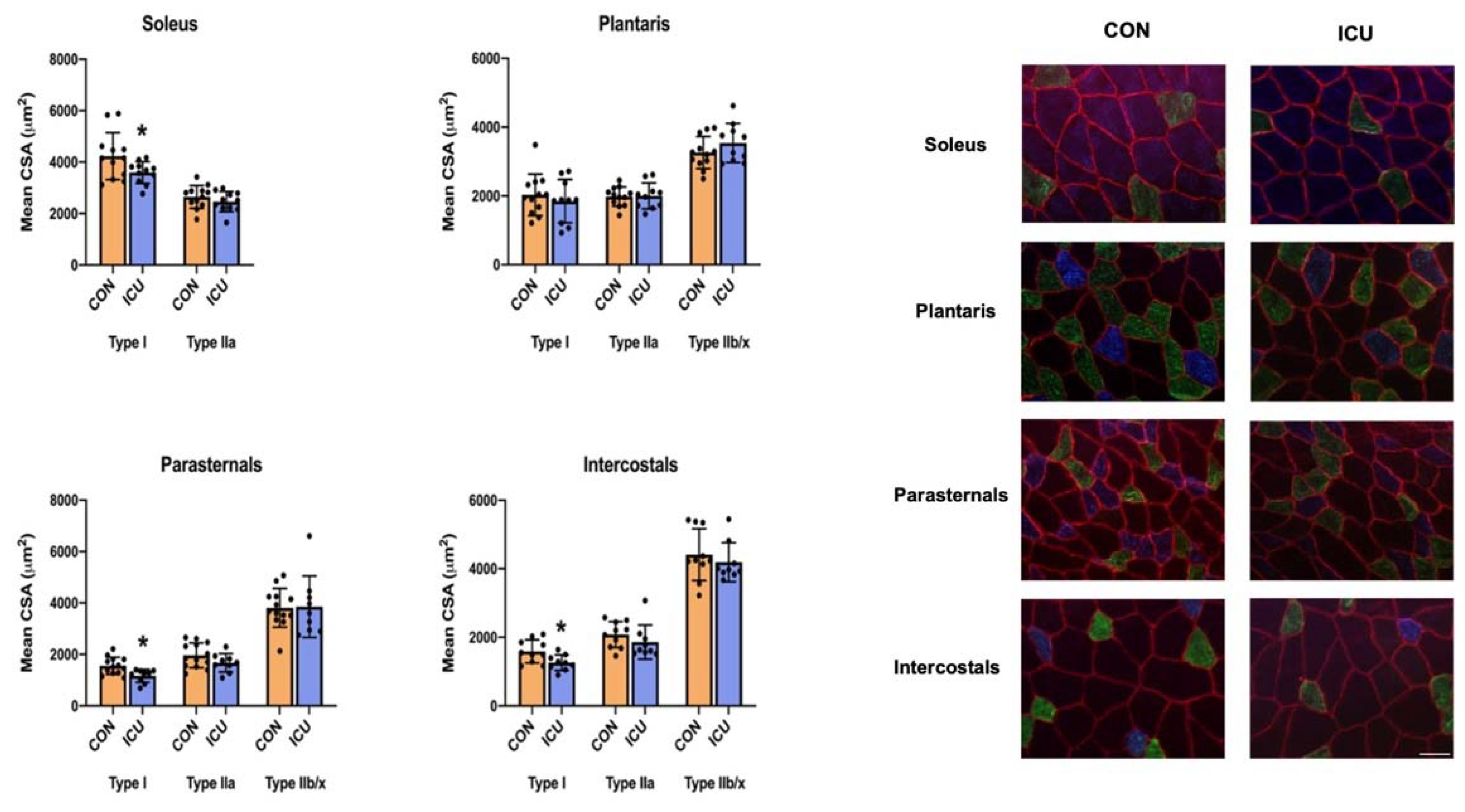
3.4. ICU-Induced Muscle Fiber Atrophy
4. Discussion
4.1. Critique of Experimental Model
4.2. ICU Care-Induced Impairment in Muscle Proteolysis Occurs Rapidly in Limb Skeletal Muscles
4.3. ICU Care-Induced Impairment in Muscle Proteolysis Occurs Rapidly in Respiratory Muscles
5. Summary and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Adhikari, N.K.; Fowler, R.A.; Bhagwanjee, S.; Rubenfeld, G.D. Critical care and the global burden of critical illness in adults. Lancet 2010, 376, 1339–1346. [Google Scholar] [CrossRef] [PubMed]
- de Jonghe, B.; Lacherade, J.C.; Sharshar, T.; Outin, H. Intensive care unit-acquired weakness: Risk factors and prevention. Crit. Care Med. 2009, 37, S309–S315. [Google Scholar] [CrossRef] [PubMed]
- Latronico, N.; Herridge, M.; Hopkins, R.O.; Angus, D.; Hart, N.; Hermans, G.; Iwashyna, T.; Arabi, Y.; Citerio, G.; Ely, E.W.; et al. The ICM research agenda on intensive care unit-acquired weakness. Intensive Care Med. 2017, 43, 1270–1281. [Google Scholar] [CrossRef] [PubMed]
- Schefold, J.C.; Bierbrauer, J.; Weber-Carstens, S. Intensive care unit-acquired weakness (ICUAW) and muscle wasting in critically ill patients with severe sepsis and septic shock. J. Cachexia Sarcopenia Muscle 2010, 1, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Nelson, W.B.; Smuder, A.J.; Hudson, M.B.; Talbert, E.E.; Powers, S.K. Cross-talk between the calpain and caspase-3 proteolytic systems in the diaphragm during prolonged mechanical ventilation. Crit. Care Med. 2012, 40, 1857–1863. [Google Scholar] [CrossRef]
- Smuder, A.J.; Nelson, W.B.; Hudson, M.B.; Kavazis, A.N.; Powers, S.K. Inhibition of the ubiquitin-proteasome pathway does not protect against ventilator-induced accelerated proteolysis or atrophy in the diaphragm. Anesthesiology 2014, 121, 115–126. [Google Scholar] [CrossRef]
- Smuder, A.J.; Sollanek, K.J.; Nelson, W.B.; Min, K.; Talbert, E.E.; Kavazis, A.N.; Hudson, M.B.; Sandri, M.; Szeto, H.H.; Powers, S.K. Crosstalk between autophagy and oxidative stress regulates proteolysis in the diaphragm during mechanical ventilation. Free Radic. Biol. Med. 2018, 115, 179–190. [Google Scholar] [CrossRef]
- Vassilakopoulos, T.; Petrof, B.J. Ventilator-induced diaphragmatic dysfunction. Am. J. Respir. Crit. Care Med. 2004, 169, 336–341. [Google Scholar] [CrossRef]
- Hudson, M.B.; Smuder, A.J.; Nelson, W.B.; Wiggs, M.P.; Shimkus, K.L.; Fluckey, J.D.; Szeto, H.H.; Powers, S.K. Partial Support Ventilation and Mitochondrial-Targeted Antioxidants Protect against Ventilator-Induced Decreases in Diaphragm Muscle Protein Synthesis. PLoS ONE 2015, 10, e0137693. [Google Scholar] [CrossRef]
- Wieske, L.; Dettling-Ihnenfeldt, D.S.; Verhamme, C.; Nollet, F.; van Schaik, I.N.; Schultz, M.J.; Horn, J.; van der Schaaf, M. Impact of ICU-acquired weakness on post-ICU physical functioning: A follow-up study. Crit. Care 2015, 19, 196. [Google Scholar] [CrossRef]
- Vanhorebeek, I.; Latronico, N.; Van den Berghe, G. ICU-acquired weakness. Intensive Care Med. 2020, 46, 637–653. [Google Scholar] [CrossRef] [PubMed]
- Hermans, G.; Van Mechelen, H.; Clerckx, B.; Vanhullebusch, T.; Mesotten, D.; Wilmer, A.; Casaer, M.P.; Meersseman, P.; Debaveye, Y.; Van Cromphaut, S.; et al. Acute outcomes and 1-year mortality of intensive care unit-acquired weakness. A cohort study and propensity-matched analysis. Am. J. Respir. Crit. Care Med. 2014, 190, 410–420. [Google Scholar] [CrossRef] [PubMed]
- Goodman, C.A.; Hornberger, T.A. Measuring protein synthesis with SUnSET: A valid alternative to traditional techniques? Exerc. Sport Sci. Rev. 2013, 41, 107–115. [Google Scholar] [CrossRef]
- Harris, A.S.; Croall, D.E.; Morrow, J.S. The calmodulin-binding site in alpha-fodrin is near the calcium-dependent protease-I cleavage site. J. Biol. Chem. 1988, 263, 15754–15761. [Google Scholar] [CrossRef] [PubMed]
- Nath, R.; Probert, A., Jr.; McGinnis, K.M.; Wang, K.K. Evidence for activation of caspase-3-like protease in excitotoxin- and hypoxia/hypoglycemia-injured neurons. J. Neurochem. 1998, 71, 186–195. [Google Scholar] [CrossRef]
- Xia, Q.; Huang, X.; Huang, J.; Zheng, Y.; March, M.E.; Li, J.; Wei, Y. The Role of Autophagy in Skeletal Muscle Diseases. Front. Physiol. 2021, 12, 638983. [Google Scholar] [CrossRef]
- Tang, H.; Smith, I.J.; Hussain, S.N.; Goldberg, P.; Lee, M.; Sugiarto, S.; Godinez, G.L.; Singh, B.K.; Payan, D.G.; Rando, T.A.; et al. The JAK-STAT pathway is critical in ventilator-induced diaphragm dysfunction. Mol. Med. 2015, 20, 579–589. [Google Scholar] [CrossRef]
- Rodriguez, B.; Larsson, L.; Z’Graggen, W.J. Critical Illness Myopathy: Diagnostic Approach and Resulting Therapeutic Implications. Curr. Treat Options Neurol. 2022, 24, 173–182. [Google Scholar] [CrossRef]
- Levine, S.; Nguyen, T.; Taylor, N.; Friscia, M.E.; Budak, M.T.; Rothenberg, P.; Zhu, J.; Sachdeva, R.; Sonnad, S.; Kaiser, L.R.; et al. Rapid disuse atrophy of diaphragm fibers in mechanically ventilated humans. N. Engl. J. Med. 2008, 358, 1327–1335. [Google Scholar] [CrossRef]
- Powers, S.K.; Wiggs, M.P.; Sollanek, K.J.; Smuder, A.J. Ventilator-induced diaphragm dysfunction: Cause and effect. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R464–R477. [Google Scholar] [CrossRef]
- Shanely, R.A.; Zergeroglu, M.A.; Lennon, S.L.; Sugiura, T.; Yimlamai, T.; Enns, D.; Belcastro, A.; Powers, S.K. Mechanical ventilation-induced diaphragmatic atrophy is associated with oxidative injury and increased proteolytic activity. Am. J. Respir. Crit. Care Med. 2002, 166, 1369–1374. [Google Scholar] [CrossRef] [PubMed]
- Agten, A.; Maes, K.; Smuder, A.; Powers, S.K.; Decramer, M.; Gayan-Ramirez, G. N-Acetylcysteine protects the rat diaphragm from the decreased contractility associated with controlled mechanical ventilation. Crit. Care Med. 2011, 39, 777–782. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Shanely, R.A.; Coombes, J.S.; Koesterer, T.J.; McKenzie, M.; Van Gammeren, D.; Cicale, M.; Dodd, S.L. Mechanical ventilation results in progressive contractile dysfunction in the diaphragm. J. Appl. Physiol. 2002, 92, 1851–1858. [Google Scholar] [CrossRef] [PubMed]
- Delp, M.D.; Duan, C. Composition and size of type I, IIA, IID/X, and IIB fibers and citrate synthase activity of rat muscle. J. Appl. Physiol. 1996, 80, 261–270. [Google Scholar] [CrossRef] [PubMed]
- De Jonghe, B.; Sharshar, T.; Lefaucheur, J.P.; Authier, F.J.; Durand-Zaleski, I.; Boussarsar, M.; Cerf, C.; Renaud, E.; Mesrati, F.; Carlet, J.; et al. Paresis acquired in the intensive care unit: A prospective multicenter study. JAMA 2002, 288, 2859–2867. [Google Scholar] [CrossRef]
- Batt, J.; Herridge, M.S.; Dos Santos, C.C. From skeletal muscle weakness to functional outcomes following critical illness: A translational biology perspective. Thorax 2019, 74, 1091–1098. [Google Scholar] [CrossRef]
- Ali, N.A.; O’Brien, J.M., Jr.; Hoffmann, S.P.; Phillips, G.; Garland, A.; Finley, J.C.; Almoosa, K.; Hejal, R.; Wolf, K.M.; Lemeshow, S.; et al. Acquired weakness, handgrip strength, and mortality in critically ill patients. Am. J. Respir. Crit. Care Med. 2008, 178, 261–268. [Google Scholar] [CrossRef]
- Sharshar, T.; Bastuji-Garin, S.; Stevens, R.D.; Durand, M.C.; Malissin, I.; Rodriguez, P.; Cerf, C.; Outin, H.; De Jonghe, B.; Groupe de Réflexion et d’Etude des Neuromyopathies En Réanimation. Presence and severity of intensive care unit-acquired paresis at time of awakening are associated with increased intensive care unit and hospital mortality. Crit. Care Med. 2009, 37, 3047–3053. [Google Scholar] [CrossRef]
- Puthucheary, Z.; Harridge, S.; Hart, N. Skeletal muscle dysfunction in critical care: Wasting, weakness, and rehabilitation strategies. Crit. Care Med. 2010, 38, S676–S682. [Google Scholar] [CrossRef]
- Puthucheary, Z.A.; Astin, R.; McPhail, M.J.W.; Saeed, S.; Pasha, Y.; Bear, D.E.; Constantin, D.; Velloso, C.; Manning, S.; Calvert, L.; et al. Metabolic phenotype of skeletal muscle in early critical illness. Thorax 2018, 73, 926–935. [Google Scholar] [CrossRef]
- Puthucheary, Z.A.; Rawal, J.; McPhail, M.; Connolly, B.; Ratnayake, G.; Chan, P.; Hopkinson, N.S.; Phadke, R.; Dew, T.; Sidhu, P.S.; et al. Acute skeletal muscle wasting in critical illness. JAMA 2013, 310, 1591–1600. [Google Scholar] [CrossRef]
- Staron, R.S.; Hagerman, F.C.; Hikida, R.S.; Murray, T.F.; Hostler, D.P.; Crill, M.T.; Ragg, K.E.; Toma, K. Fiber type composition of the vastus lateralis muscle of young men and women. J. Histochem. Cytochem. 2000, 48, 623–629. [Google Scholar] [CrossRef] [PubMed]
- De Troyer, A.; Estenne, M. Functional anatomy of the respiratory muscles. Clin. Chest. Med. 1988, 9, 175–193. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Demirel, H.A.; Coombes, J.S.; Fletcher, L.; Calliaud, C.; Vrabas, I.; Prezant, D. Myosin phenotype and bioenergetic characteristics of rat respiratory muscles. Med. Sci. Sport. Exerc. 1997, 29, 1573–1579. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ozdemir, M.; Bomkamp, M.P.; Hyatt, H.W.; Smuder, A.J.; Powers, S.K. Intensive Care Unit Acquired Weakness Is Associated with Rapid Changes to Skeletal Muscle Proteostasis. Cells 2022, 11, 4005. https://doi.org/10.3390/cells11244005
Ozdemir M, Bomkamp MP, Hyatt HW, Smuder AJ, Powers SK. Intensive Care Unit Acquired Weakness Is Associated with Rapid Changes to Skeletal Muscle Proteostasis. Cells. 2022; 11(24):4005. https://doi.org/10.3390/cells11244005
Chicago/Turabian StyleOzdemir, Mustafa, Matthew P. Bomkamp, Hayden W. Hyatt, Ashley J. Smuder, and Scott K. Powers. 2022. "Intensive Care Unit Acquired Weakness Is Associated with Rapid Changes to Skeletal Muscle Proteostasis" Cells 11, no. 24: 4005. https://doi.org/10.3390/cells11244005
APA StyleOzdemir, M., Bomkamp, M. P., Hyatt, H. W., Smuder, A. J., & Powers, S. K. (2022). Intensive Care Unit Acquired Weakness Is Associated with Rapid Changes to Skeletal Muscle Proteostasis. Cells, 11(24), 4005. https://doi.org/10.3390/cells11244005