
Consecutive-Day Ventricular and Atrial Cardiomyocyte Isolations from the Same Heart: Shifting the Cost–Benefit Balance of Cardiac Primary Cell Research

 ,
, 
 , , , , ,
, , , , ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Cardiac Slice Preparation
2.3. Cell Isolation
2.4. Assessment of Cell Isolation Yields
2.5. High-Resolution Imaging
2.6. Sarcomere Shortening Dynamics
2.7. Resting Membrane Potential Recordings
2.8. RNA Isolation, Reverse Transcription, and Quantitative PCR
2.9. Data Analysis
3. Results
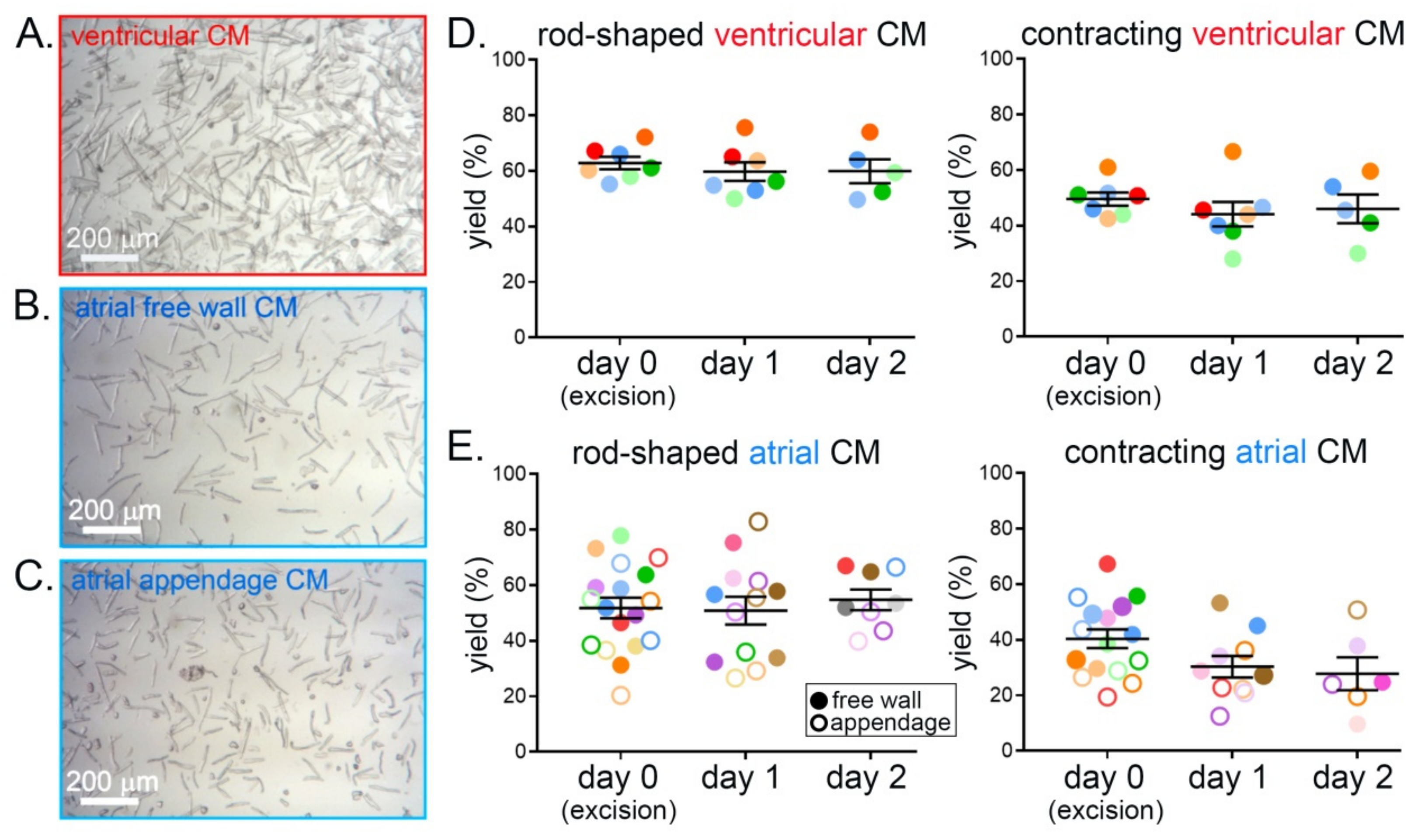
3.1. Yield of Calcium-Tolerant CM over Three Consecutive Days
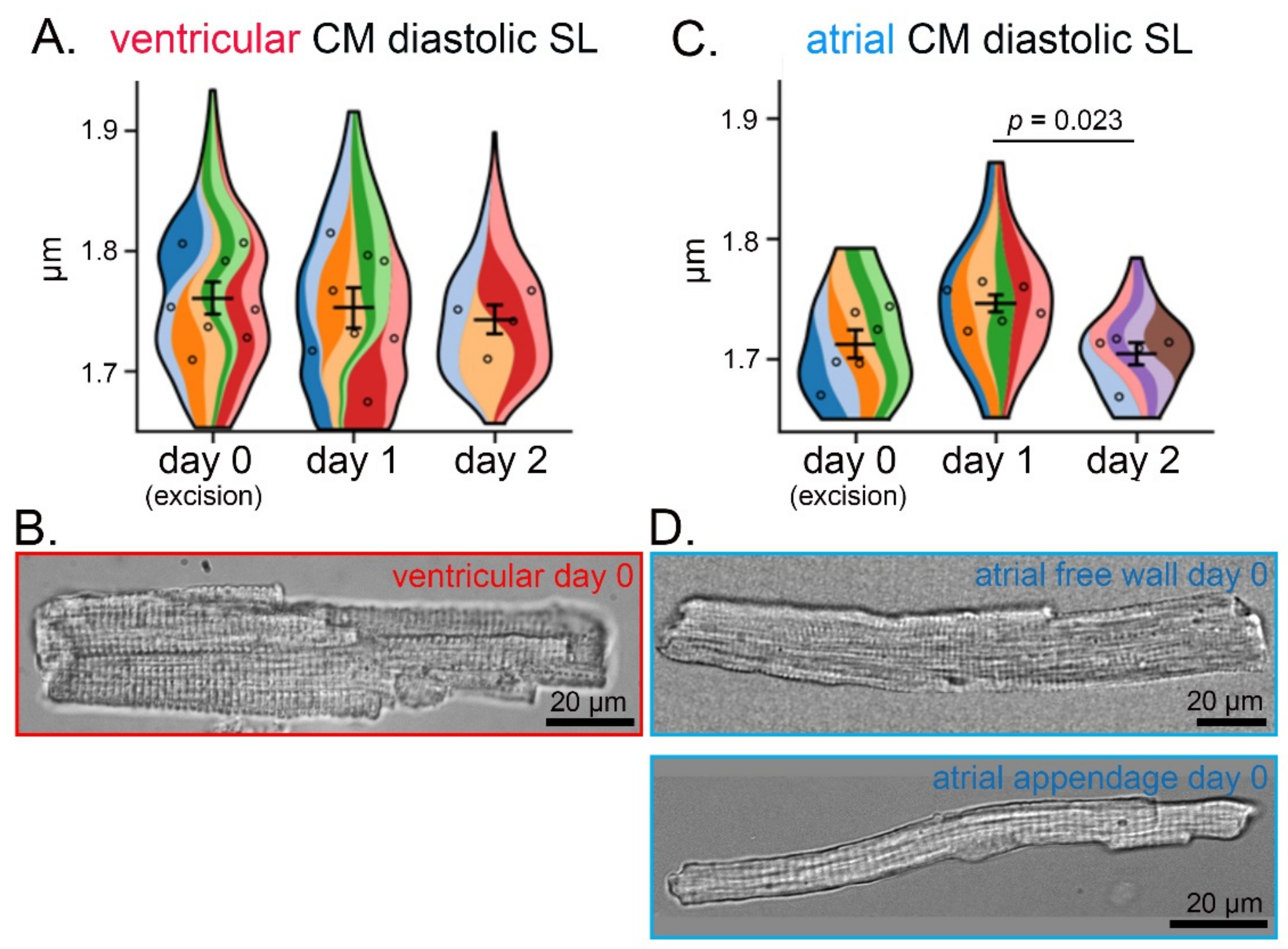
3.2. Preservation of CM Morphology over Three Consecutive Days
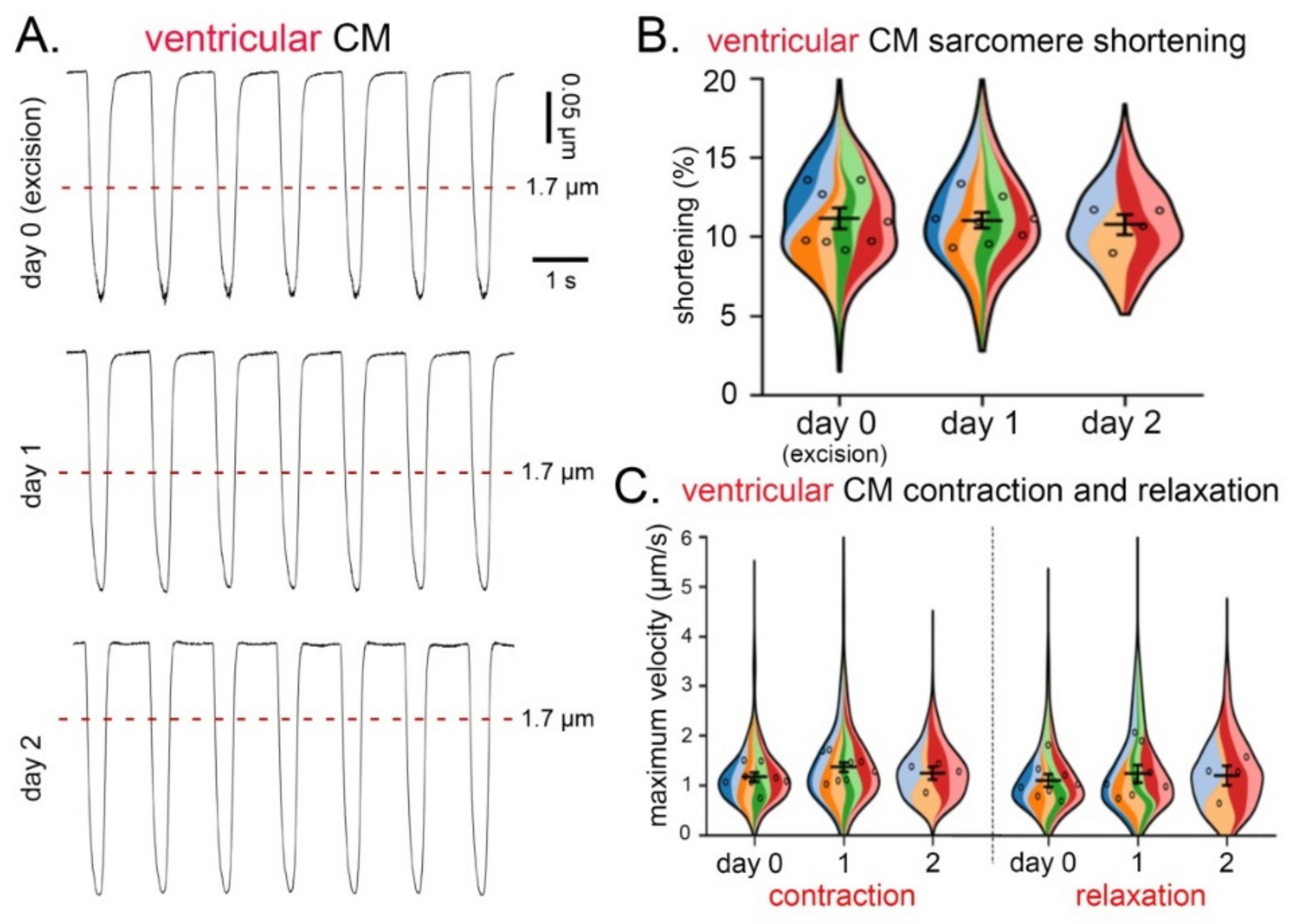
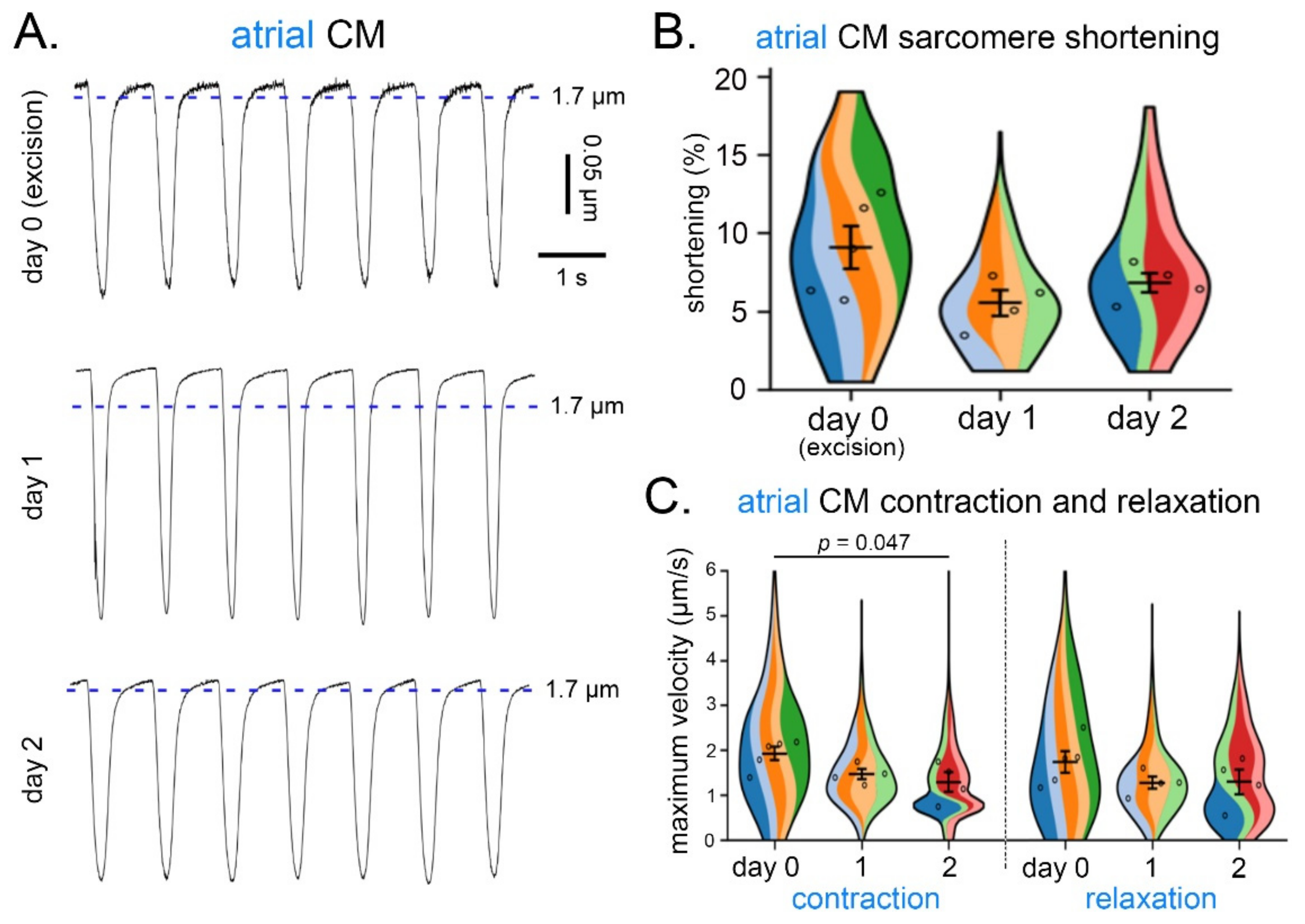
3.3. CM Contraction and Relaxation over Three Consecutive Days
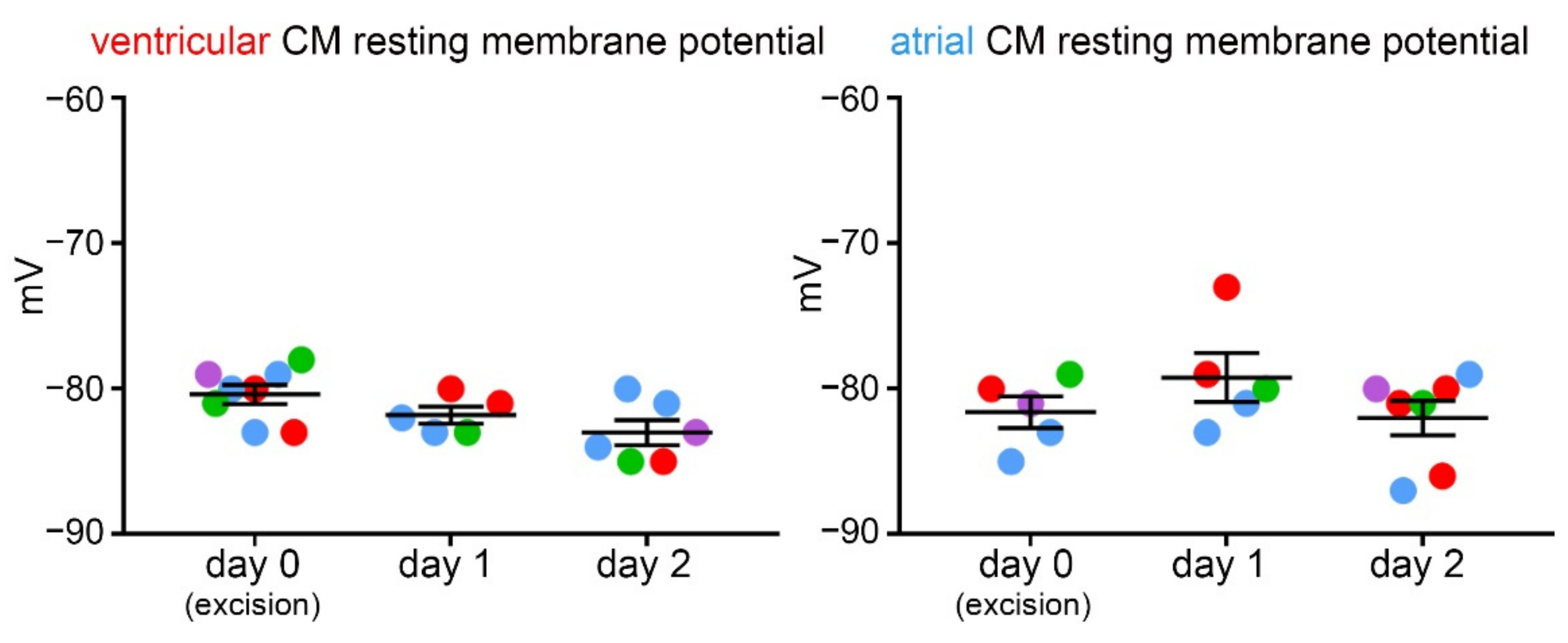
3.4. Resting Membrane Potential over Three Consecutive Days
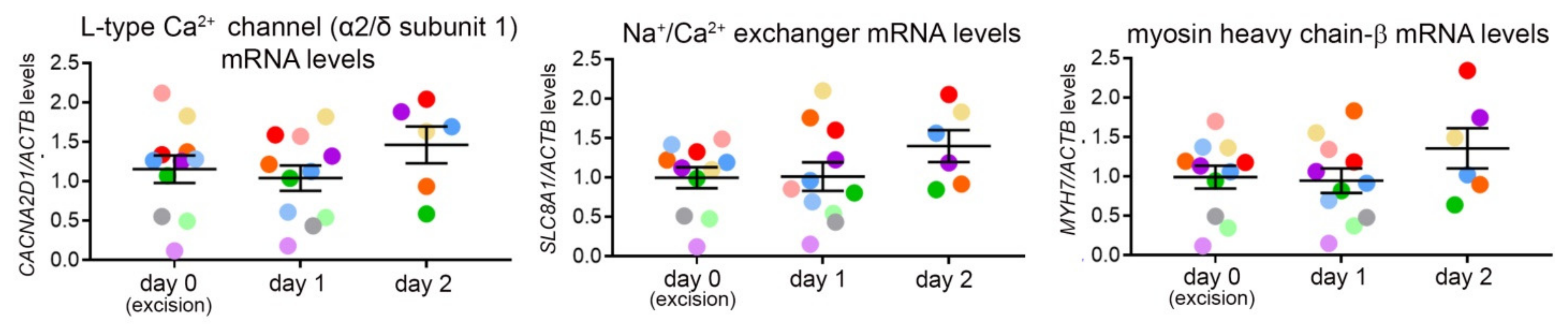
3.5. Gene Expression Analysis over Three Consecutive Days
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gould, R.P.; Powell, T. Intact isolated muscle cells from the adult rat heart. J. Physiol. 1972, 225, 16–19. [Google Scholar]
- Jacobson, S.L.; Piper, H.M. Cell cultures of adult cardiomyocytes as models of the myocardium. J. Mol. Cell Cardiol. 1986, 18, 661–678. [Google Scholar] [CrossRef]
- Mitcheson, J.S.; Hancox, J.C.; Levi, A.J. Cultured adult cardiac myocytes: Future applications, culture methods, morphological and electrophysiological properties. Cardiovasc. Res. 1998, 39, 280–300. [Google Scholar] [CrossRef]
- Volz, A.; Piper, H.M.; Siegmund, B.; Schwartz, P. Longevity of adult ventricular rat heart muscle cells in serum-free primary culture. J. Mol. Cell Cardiol. 1991, 23, 161–173. [Google Scholar] [CrossRef]
- Burton, R.A.B.; Rog-Zielinska, E.A.; Corbett, A.D.; Peyronnet, R.; Bodi, I.; Fink, M.; Sheldon, J.; Hoenger, A.; Calaghan, S.C.; Bub, G.; et al. Caveolae in rabbit ventricular myocytes: Distribution and dynamic diminution after cell isolation. Biophys. J. 2017, 113, 1047–1059. [Google Scholar] [CrossRef]
- Hammer, K.; Ruppenthal, S.; Viero, C.; Scholz, A.; Edelmann, L.; Kaestner, L.; Lipp, P. Remodelling of Ca2+ handling organelles in adult rat ventricular myocytes during long term culture. J. Mol. Cell Cardiol. 2010, 49, 427–437. [Google Scholar] [CrossRef]
- Mitcheson, J.S.; Hancox, J.C.; Levi, A.J. Action potentials, ion channel currents and transverse tubule density in adult rabbit ventricular myocytes maintained for 6 days in cell culture. Pflug. Arch. 1996, 431, 814–827. [Google Scholar] [CrossRef]
- Himmel, H.M.; Pietsch, M.; Streller, U.; Graf, E.M.; Ravens, U. Changes in morphology and inward rectifier currents in human atrial myocytes depend on culture conditions. Basic Res. Cardiol. 2002, 97, 434–444. [Google Scholar] [CrossRef]
- Ellingsen, O.; Davidoff, A.J.; Prasad, S.K.; Berger, H.-J.; Springhorn, J.P.; Marsh, J.D.; Kelly, R.A.; Smith, T.W. Adult rat ventricular myocytes cultured in defined medium: Phenotype and electromechanical function. Am. J. Physiol. Heart Circ. Physiol. 1993, 265, 747–754. [Google Scholar] [CrossRef]
- Laird, D.W.; Puranam, K.L.; Revel, J.P. Turnover and phosphorylation dynamics of connexin-43 gap junction protein in cultured cardiac myocytes. Biochem. J. 1991, 273, 67–72. [Google Scholar] [CrossRef]
- Louch, W.E.; Sheehan, K.A.; Wolska, B.M. Methods in cardiomyocyte isolation, culture, and gene transfer. J. Mol. Cell Cardiol. 2011, 51, 288–298. [Google Scholar] [CrossRef]
- Fischer, C.; Milting, H.; Fein, E.; Reiser, E.; Lu, K.; Seidel, T.; Schinner, C.; Schwarzmayr, T.; Schramm, R.; Tomasi, R.; et al. Long-term functional and structural preservation of precision-cut human myocardium under continuous electromechanical stimulation in vitro. Nat. Commun. 2019, 10, 117. [Google Scholar] [CrossRef]
- Fiegle, D.J.; Volk, T.; Seidel, T. Isolation of human ventricular cardiomyocytes from vibratome-cut myocardial slices. J. Vis. Exp. 2020, 159, e61167. [Google Scholar] [CrossRef]
- Peyronnet, R.; Bollensdorff, C.; Capel, R.A.; Rog-Zielinska, E.A.; Woods, C.E.; Charo, D.N.; Lookin, O.; Fajardo, G.; Ho, M.; Quertermous, T.; et al. Load-dependent effects of apelin on murine cardiomyocytes. Prog. Biophys. Mol. Biol. 2017, 130, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Bernal Sierra, Y.A.; Rost, B.R.; Pofahl, M.; Fernandes, A.M.; Kopton, R.A.; Moser, S.; Holtkamp, D.; Masala, N.; Beed, P.; Tukker, J.J.; et al. Potassium channel-based optogenetic silencing. Nat. Commun. 2018, 9, 4611. [Google Scholar] [CrossRef]
- Kenny, M.; Schoen, I. Violin SuperPlots: Visualizing replicate heterogeneity in large data sets. Mol. Biol. Cell 2021, 32, 1333–1334. [Google Scholar] [CrossRef]
- Odening, K.E.; Kohl, P. Follow the white rabbit: Experimental and computational models of the rabbit heart provide insights into cardiac (patho-) physiology. Prog. Biophys. Mol. Biol. 2016, 121, 75–76. [Google Scholar] [CrossRef]
- Hebisch, S.; Bischoff, E.; Soboll, S. Influence of 2,3-butanedione monoxime on heart energy metabolism. Basic Res. Cardiol. 1993, 88, 566–575. [Google Scholar] [CrossRef]
- Lee, K.C.; Chang, C.Y.; Chuang, Y.C.; Sue, S.H.; Yang, H.S.; Weng, C.F.; Lee, Y.T.; Huang, W.S.; Chen, I.C.; Wei, J. Combined St. Thomas and histidine-tryptophan-ketoglutarat solutions for myocardial preservation in heart transplantation patients. Transpl. Proc. 2012, 44, 886–889. [Google Scholar] [CrossRef]
- Jahania, M.S.; Sanchez, J.A.; Narayan, P.; Lasley, R.D.; Mentzer, R.M., Jr. Heart preservation for transplantation: Principles and strategies. Ann. Thorac. Surg. 1999, 68, 1983–1987. [Google Scholar] [CrossRef]
- Gharagozloo, F.; Melendez, F.J.; Hein, R.A.; Laurence, R.G.; Shemin, R.J.; DiSesa, V.J.; Cohn, L.H. The effect of amino acid L-glutamate on the extended preservation ex vivo of the heart for transplantation. Circulation 1987, 76, 65–70. [Google Scholar]
- Chung, C.S.; Mechas, C.; Campbell, K.S. Myocyte contractility can be maintained by storing cells with the myosin ATPase inhibitor 2,3 butanedione monoxime. Physiol. Rep. 2015, 3, e12445. [Google Scholar] [CrossRef] [PubMed]
- Watson, S.A.; Scigliano, M.; Bardi, I.; Ascione, R.; Terracciano, C.M.; Perbellini, F. Preparation of viable adult ventricular myocardial slices from large and small mammals. Nat. Protoc. 2017, 12, 2623–2639. [Google Scholar] [CrossRef] [PubMed]
- Kivisto, T.; Makiranta, M.; Oikarinen, E.L.; Karhu, S.; Weckstrom, M.; Sellin, L.C. 2,3-butanedione monoxime (BDM) increases initial yields and improves long-term survival of isolated cardiac myocytes. Jpn. J. Physiol. 1995, 45, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Mulieri, L.A.; Hasenfuss, G.; Ittleman, F.; Blanchard, E.M.; Alpert, N.R. Protection of human left ventricular myocardium from cutting injury with 2,3-butanedione monoxime. Circ. Res. 1989, 65, 1441–1449. [Google Scholar] [CrossRef]
- Hall, A.R.; Hausenloy, D.J. Mitochondrial respiratory inhibition by 2,3-butanedione monoxime (BDM): Implications for culturing isolated mouse ventricular cardiomyocytes. Physiol. Rep. 2016, 4, e12606. [Google Scholar] [CrossRef]
- Abi-Gerges, N.; Pointon, A.; Pullen, G.F.; Morton, M.J.; Oldman, K.L.; Armstrong, D.; Valentin, J.P.; Pollard, C.E. Preservation of cardiomyocytes from the adult heart. J. Mol. Cell Cardiol. 2013, 64, 108–119. [Google Scholar] [CrossRef]
- Brandenburg, S.; Arakel, E.C.; Schwappach, B.; Lehnart, S.E. The molecular and functional identities of atrial cardiomyocytes in health and disease. Biochim. Biophys. Acta Mol. Cell Res. 2016, 1863, 1882–1893. [Google Scholar] [CrossRef]
- Guo, G.-R.; Chen, L.; Rao, M.; Chen, K.; Song, J.P.; Hu, S.S. A modified method for isolation of human cardiomyocytes to model cardiac diseases. J. Transl. Med. 2018, 16, 288. [Google Scholar] [CrossRef]
- Stringham, J.C.; Paulsen, K.L.; Southard, J.H.; Mentzer, R.M., Jr.; Belzer, F.O. Forty-hour preservation of the rabbit heart: Optimal osmolarity, [Mg2+], and pH of a modified UW solution. Ann. Thorac. Surg. 1994, 58, 7–13. [Google Scholar] [CrossRef]
- Mitcheson, J.S.; Hancox, J.C.; Levi, A.J. Cultured adult rabbit myocytes: Effect of adding supplements to the medium, and response to isoprenaline. J. Cardiovasc. Electrophysiol. 1997, 8, 1020–1030. [Google Scholar] [CrossRef]
- Meki, M.H.; Miller, J.M.; Mohamed, T.M.A. Heart slices to model cardiac physiology. Front. Pharmacol. 2021, 12, 617922. [Google Scholar] [CrossRef]
- Berger, H.J.; Prasad, S.K.; Davidoff, A.J.; Pimental, D.; Ellingsen, O.; Marsh, J.D.; Smith, T.W.; Kelly, R.A. Continual electric field stimulation preserves contractile function of adult ventricular myocytes in primary culture. Am. J. Physiol. 1994, 266, 341–349. [Google Scholar] [CrossRef]
- Edelmann, J.C.; Jones, L.; Peyronnet, R.; Lu, L.; Kohl, P.; Ravens, U. A Bioreactor to apply multimodal physical stimuli to cultured cells. Methods Mol. Biol. 2016, 1502, 21–33. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Greiner, J.; Schiatti, T.; Kaltenbacher, W.; Dente, M.; Semenjakin, A.; Kok, T.; Fiegle, D.J.; Seidel, T.; Ravens, U.; Kohl, P.; et al. Consecutive-Day Ventricular and Atrial Cardiomyocyte Isolations from the Same Heart: Shifting the Cost–Benefit Balance of Cardiac Primary Cell Research. Cells 2022, 11, 233. https://doi.org/10.3390/cells11020233
Greiner J, Schiatti T, Kaltenbacher W, Dente M, Semenjakin A, Kok T, Fiegle DJ, Seidel T, Ravens U, Kohl P, et al. Consecutive-Day Ventricular and Atrial Cardiomyocyte Isolations from the Same Heart: Shifting the Cost–Benefit Balance of Cardiac Primary Cell Research. Cells. 2022; 11(2):233. https://doi.org/10.3390/cells11020233
Chicago/Turabian StyleGreiner, Joachim, Teresa Schiatti, Wenzel Kaltenbacher, Marica Dente, Alina Semenjakin, Thomas Kok, Dominik J. Fiegle, Thomas Seidel, Ursula Ravens, Peter Kohl, and et al. 2022. "Consecutive-Day Ventricular and Atrial Cardiomyocyte Isolations from the Same Heart: Shifting the Cost–Benefit Balance of Cardiac Primary Cell Research" Cells 11, no. 2: 233. https://doi.org/10.3390/cells11020233
APA StyleGreiner, J., Schiatti, T., Kaltenbacher, W., Dente, M., Semenjakin, A., Kok, T., Fiegle, D. J., Seidel, T., Ravens, U., Kohl, P., Peyronnet, R., & Rog-Zielinska, E. A. (2022). Consecutive-Day Ventricular and Atrial Cardiomyocyte Isolations from the Same Heart: Shifting the Cost–Benefit Balance of Cardiac Primary Cell Research. Cells, 11(2), 233. https://doi.org/10.3390/cells11020233