
Effect of 3D Synthetic Microscaffold Nichoid on the Morphology of Cultured Hippocampal Neurons and Astrocytes

 ,
,  ,
,  ,
,  ,
,  , and
, and 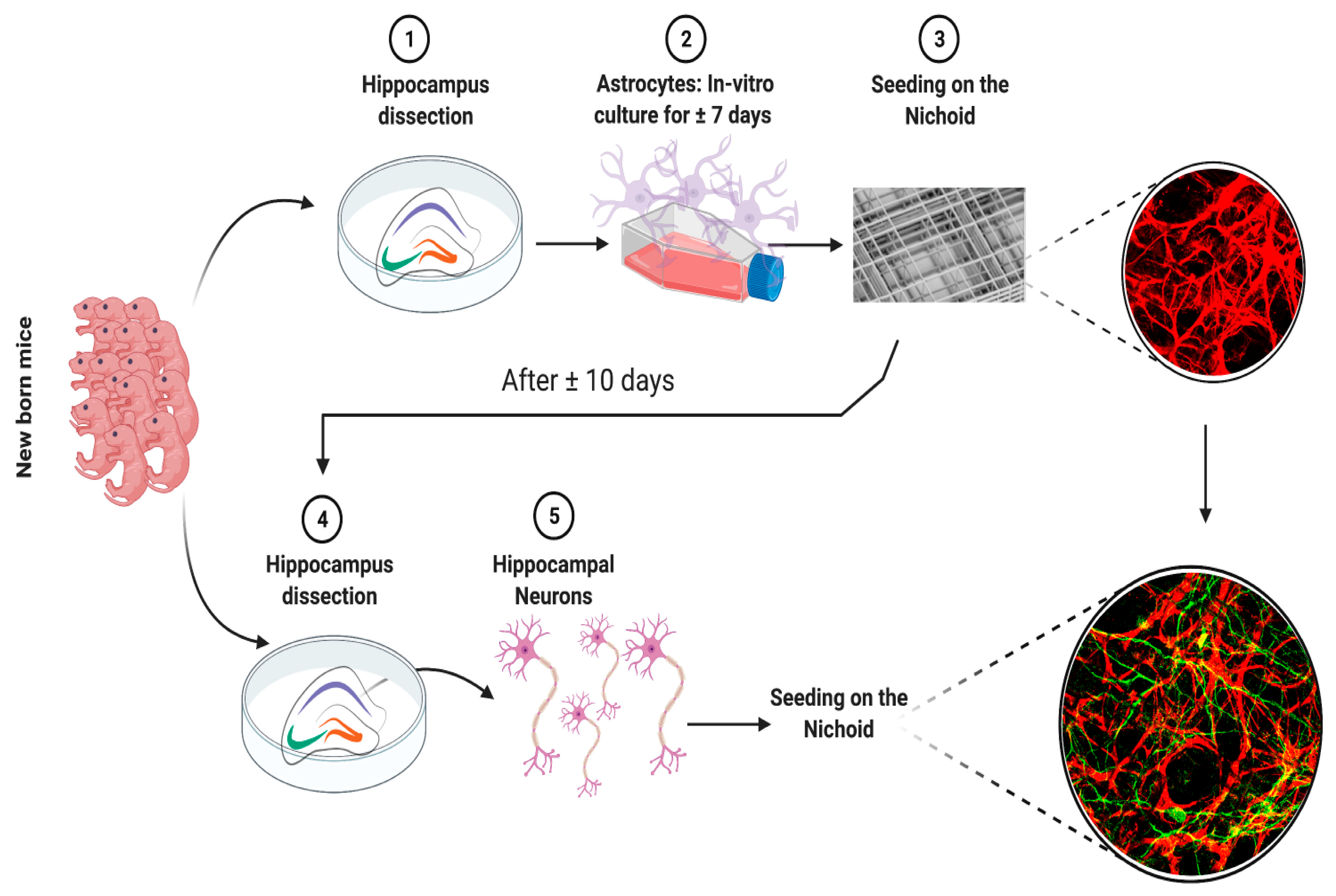{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
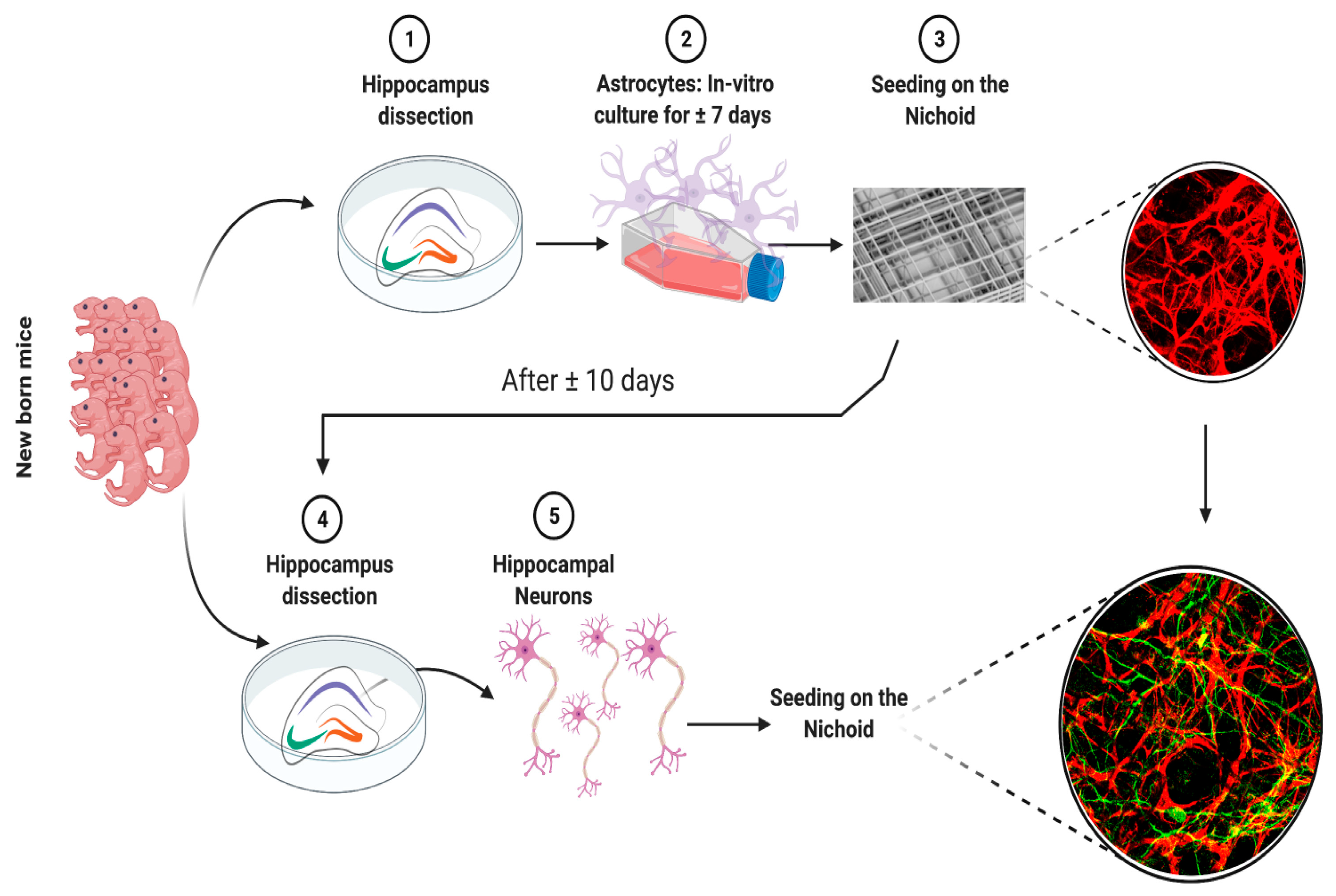
2. Materials and Methods
2.1. Laser Microfabrication
2.2. Animal Procedures
2.3. Primary Astrocyte Culture
2.4. Primary Neuronal Culture
2.5. Set-Up of the Co-Culture
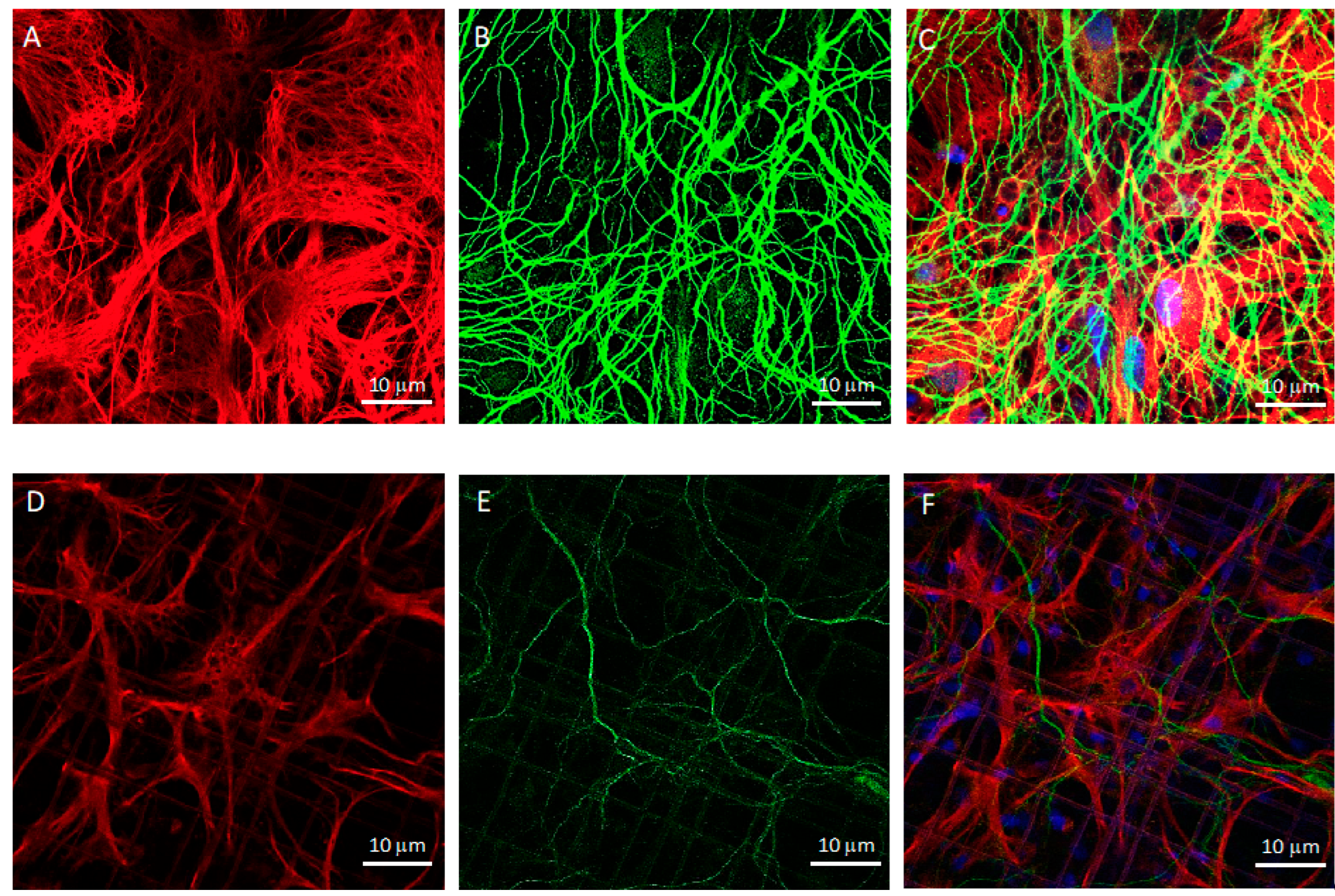
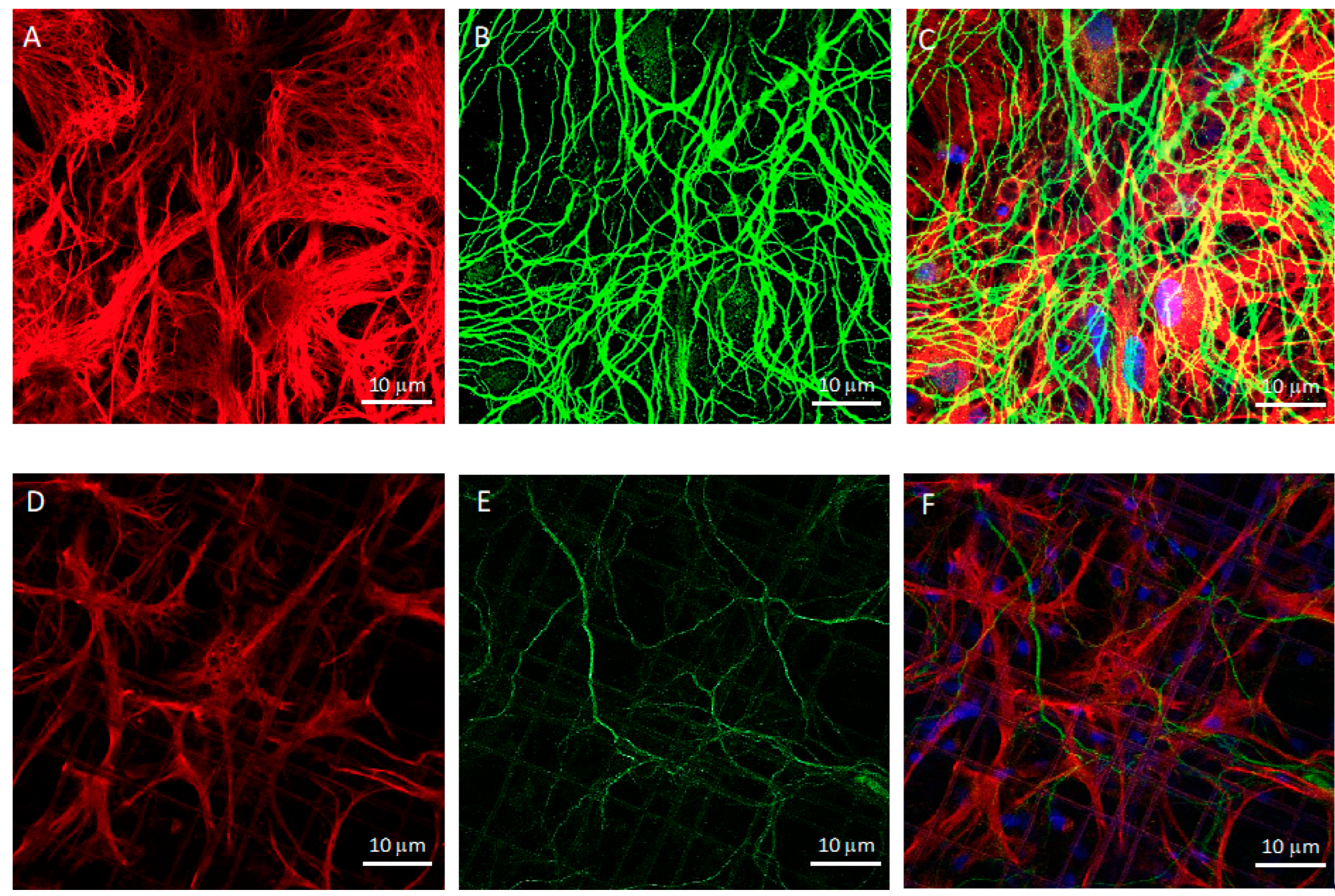
2.6. Immunofluorescence
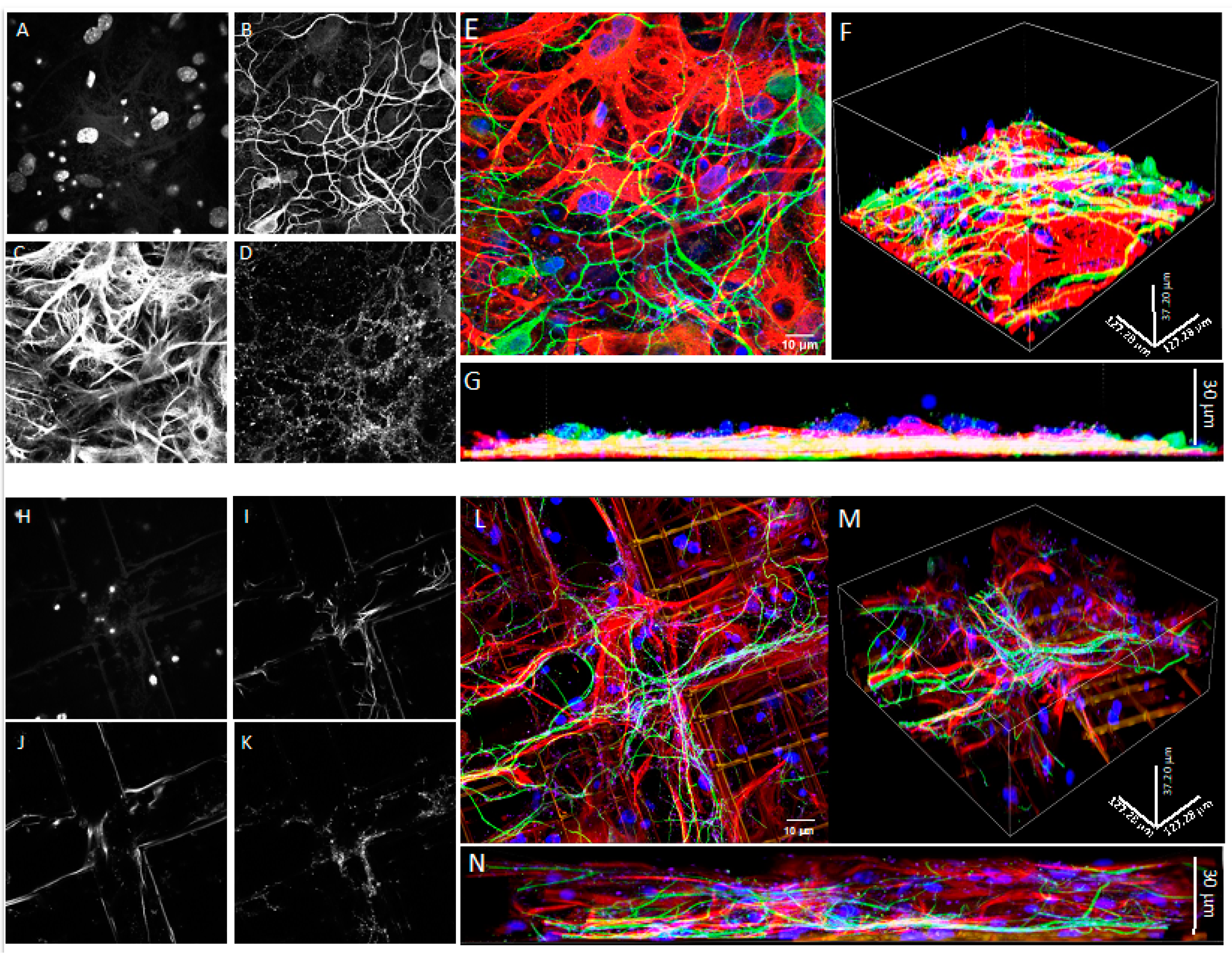
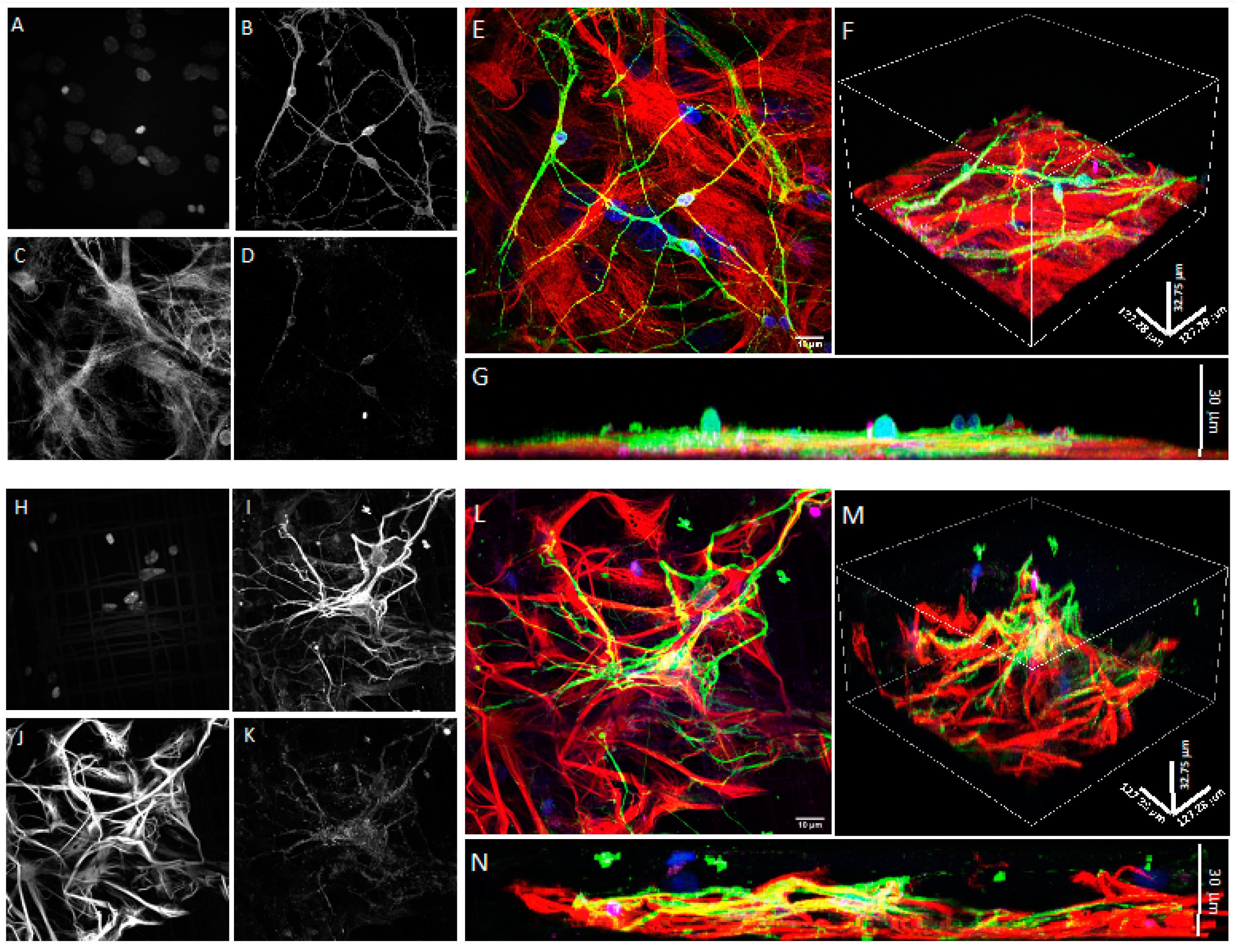
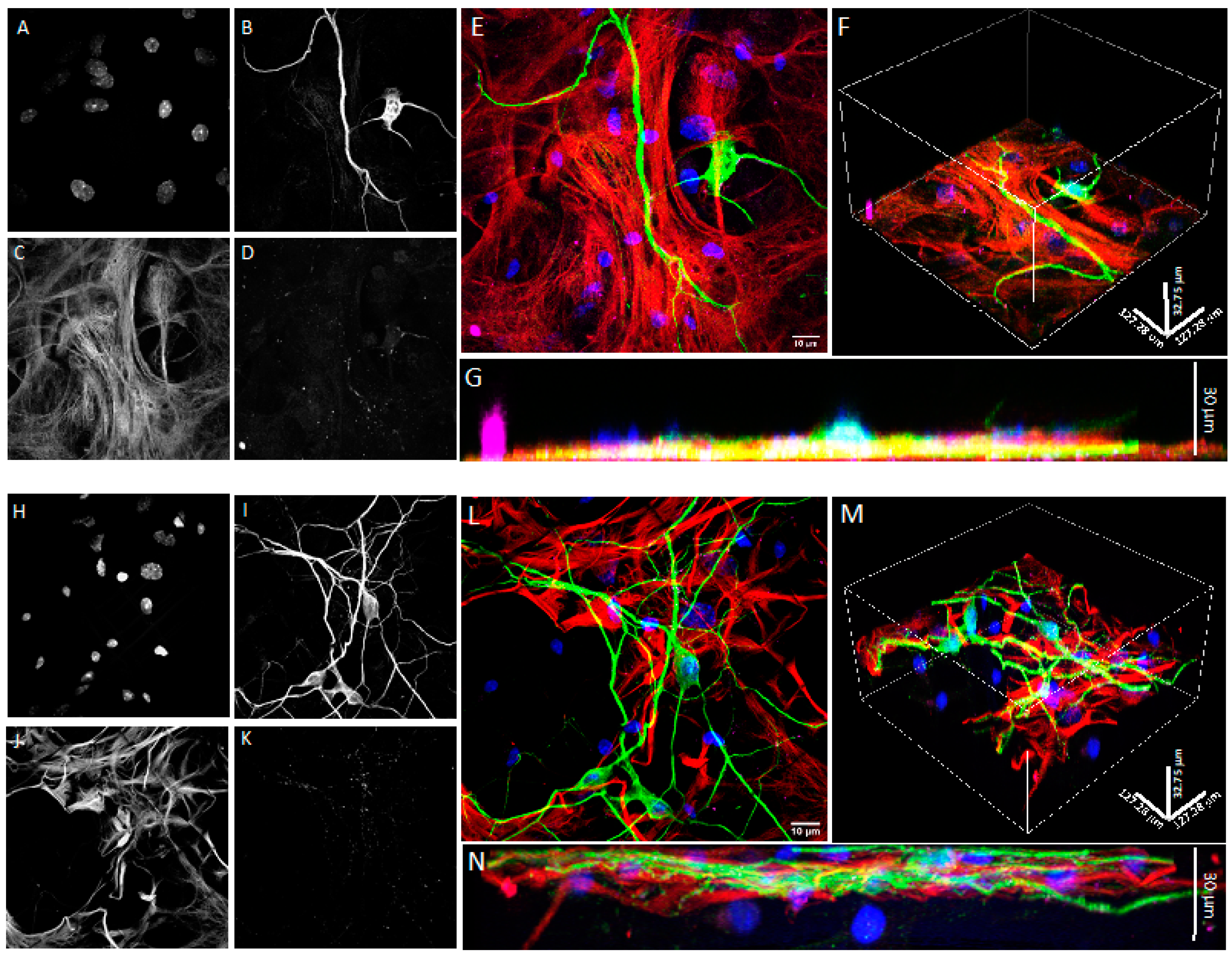
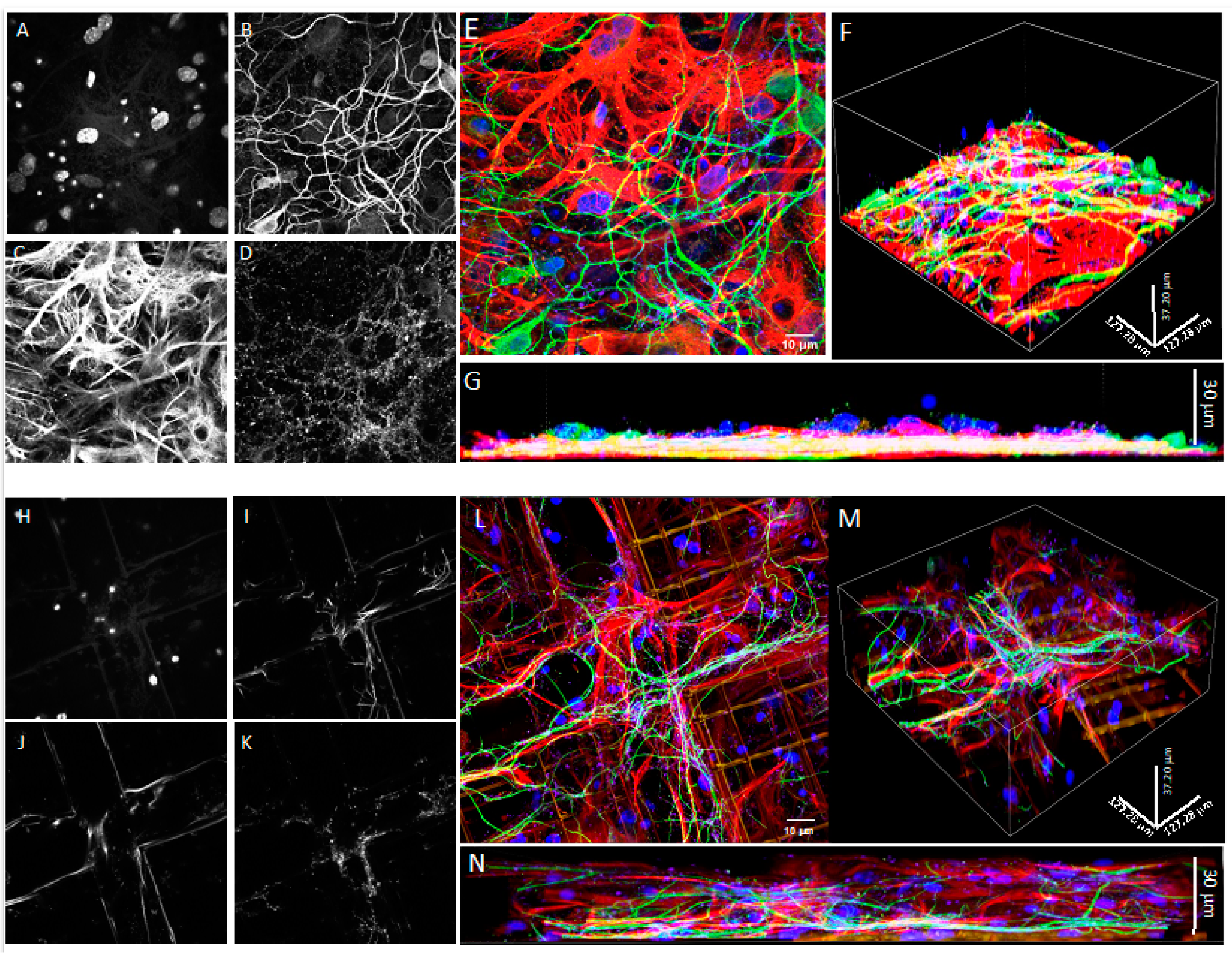
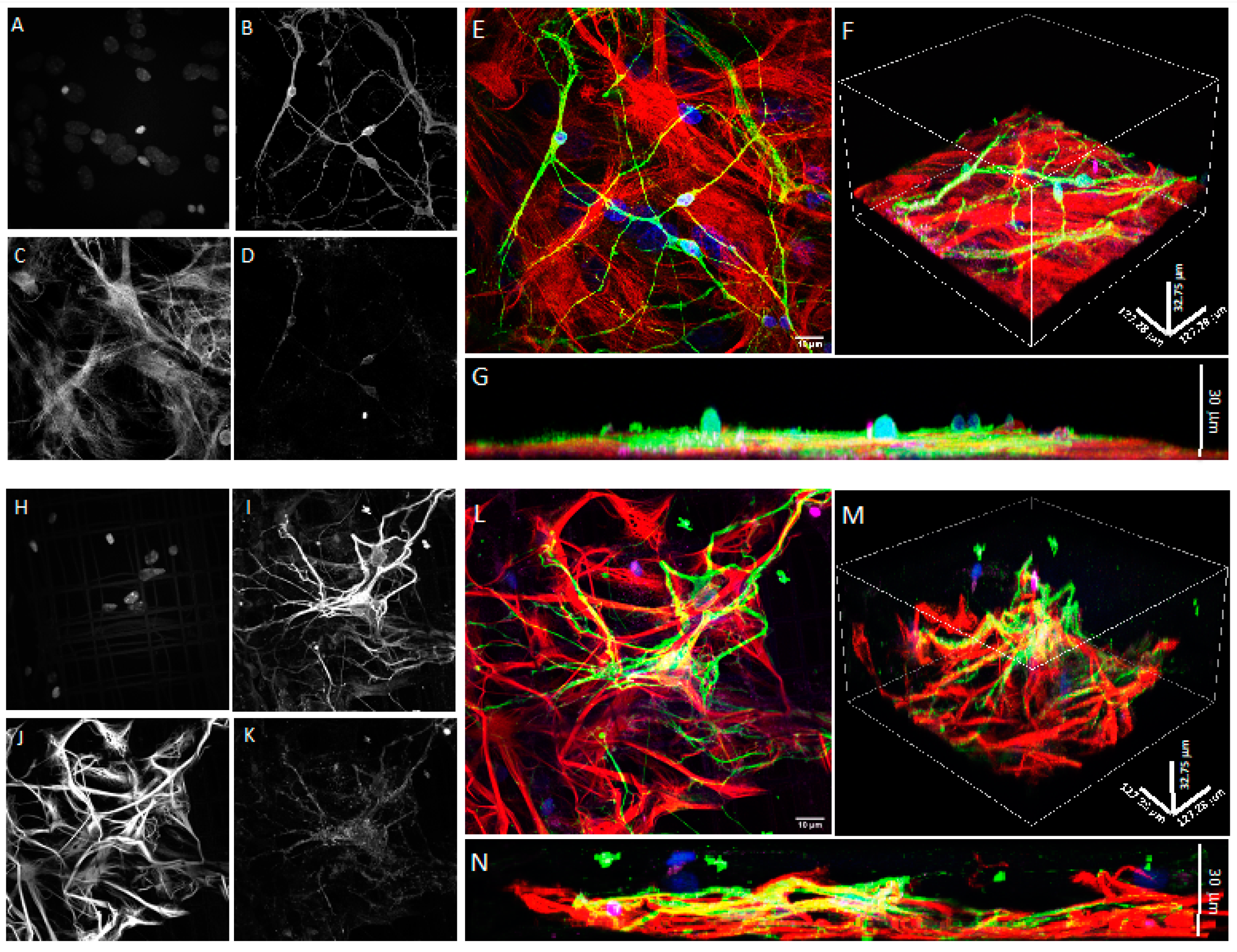
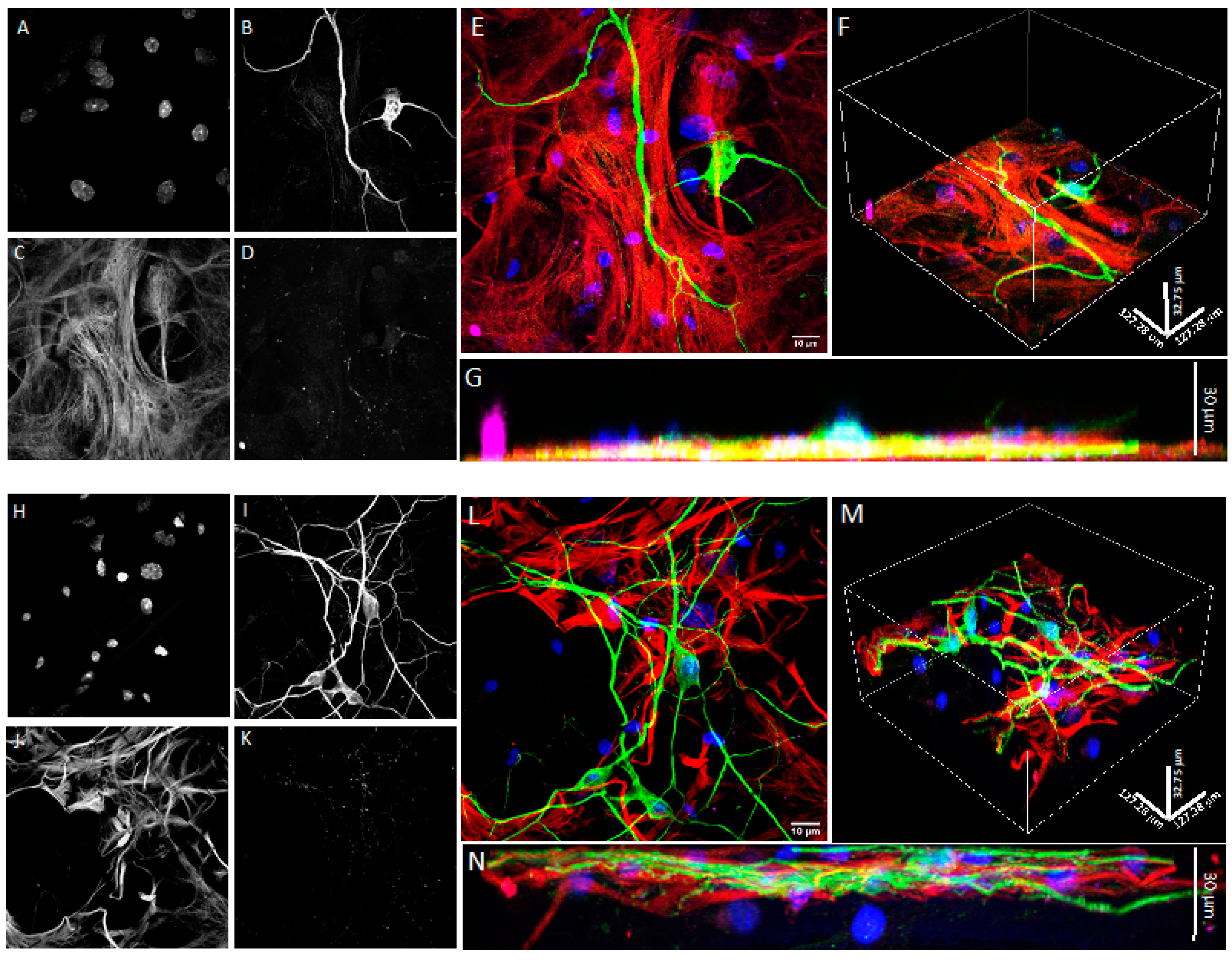
2.7. Confocal Microscopy
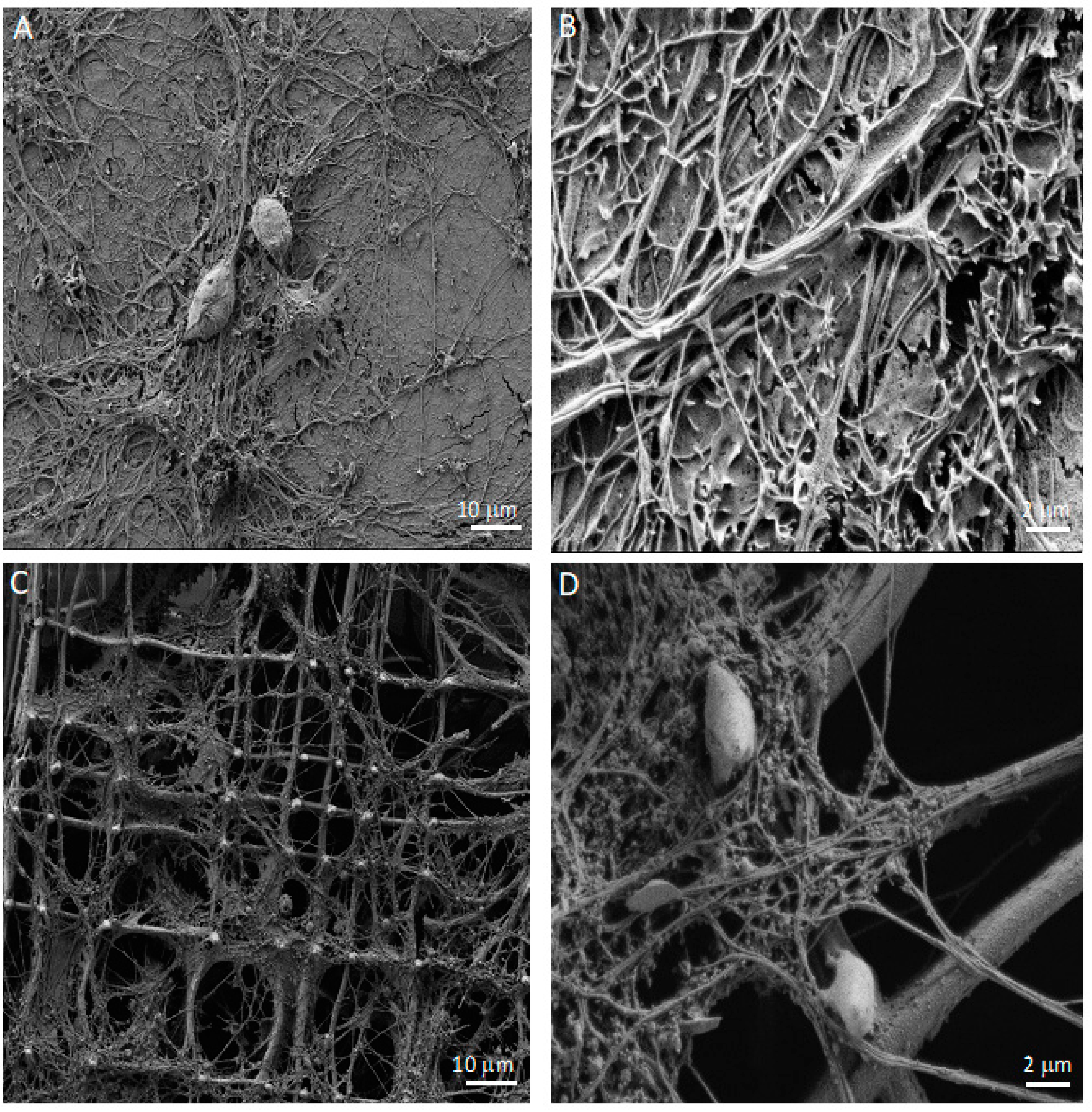
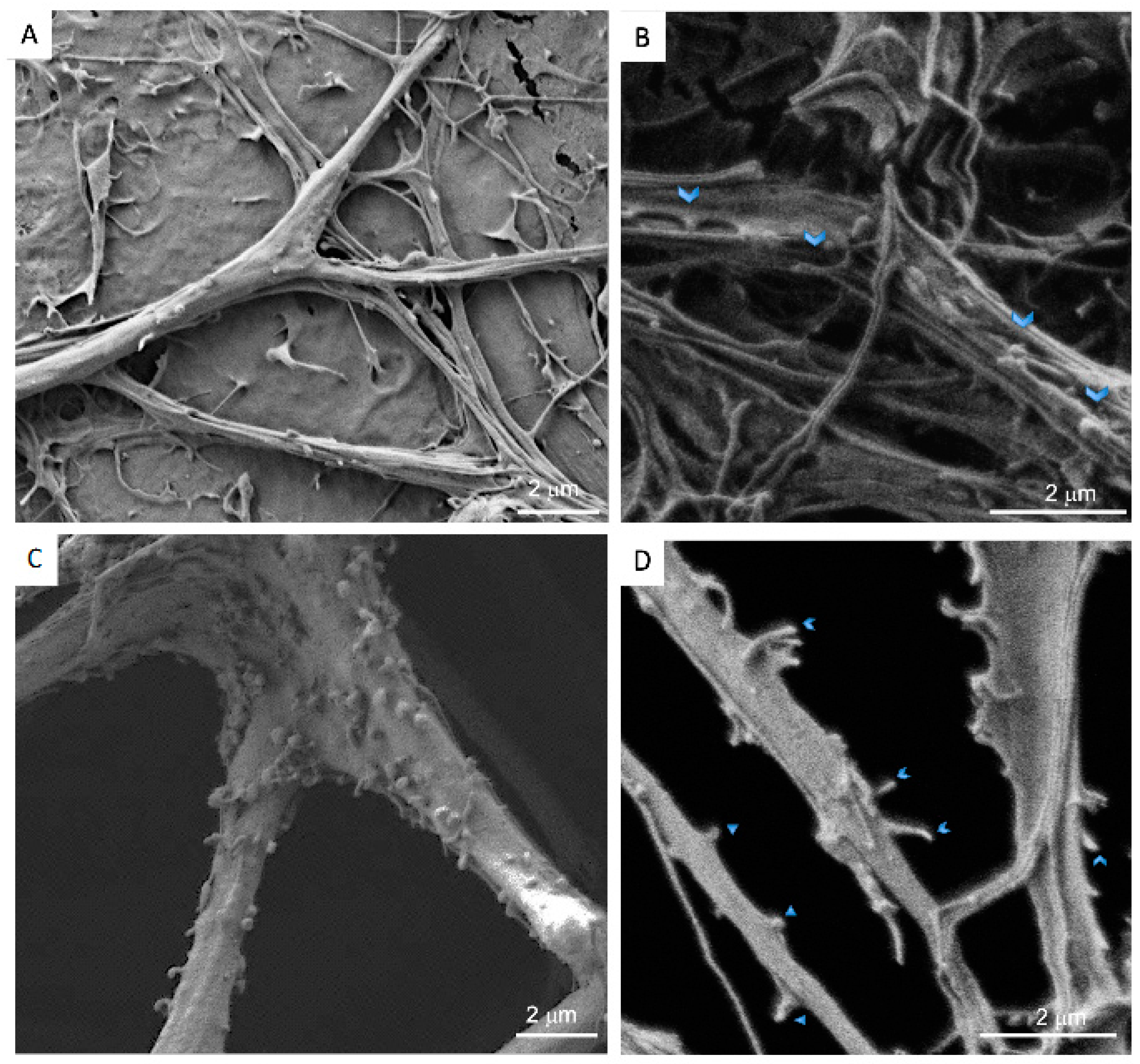
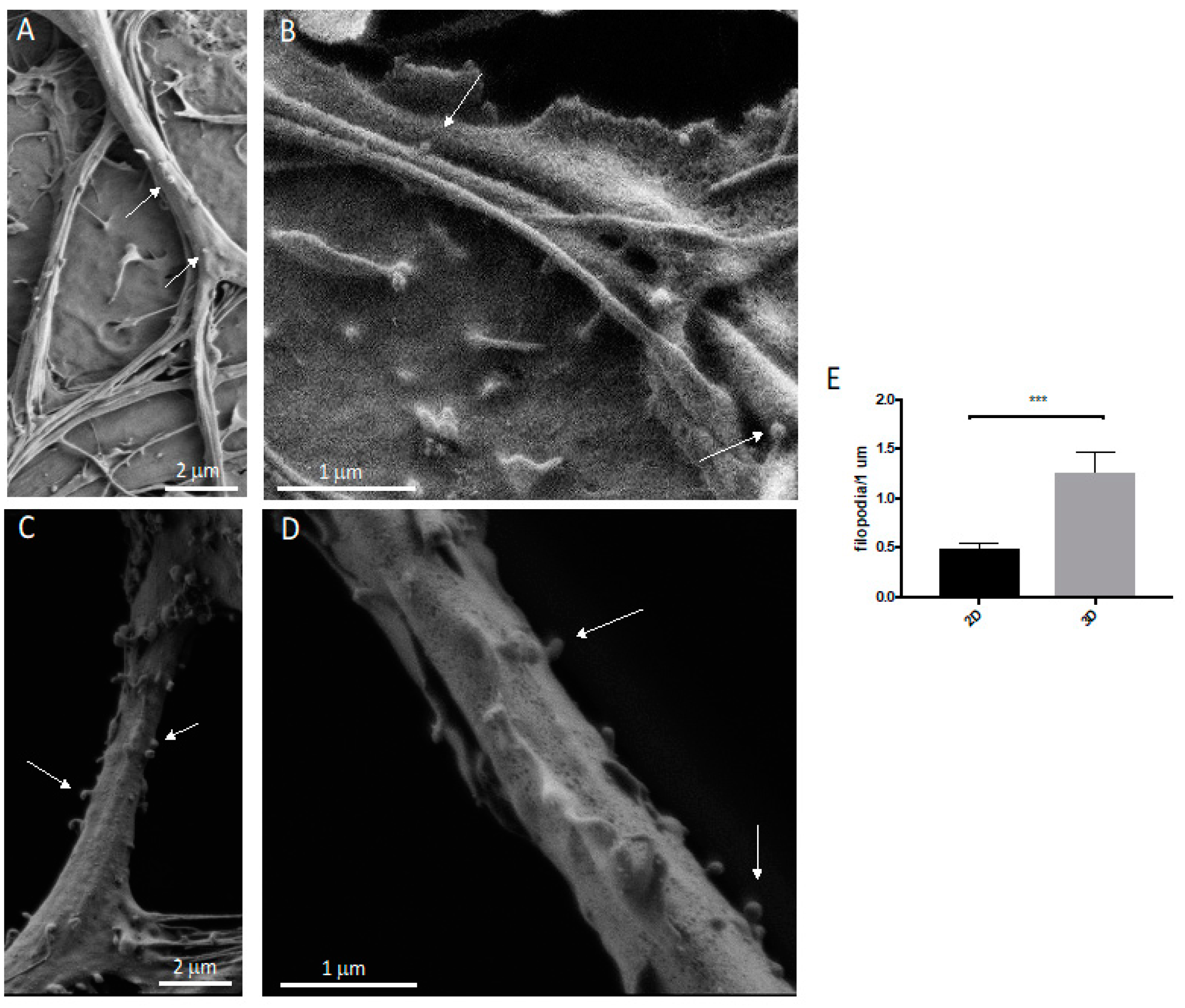
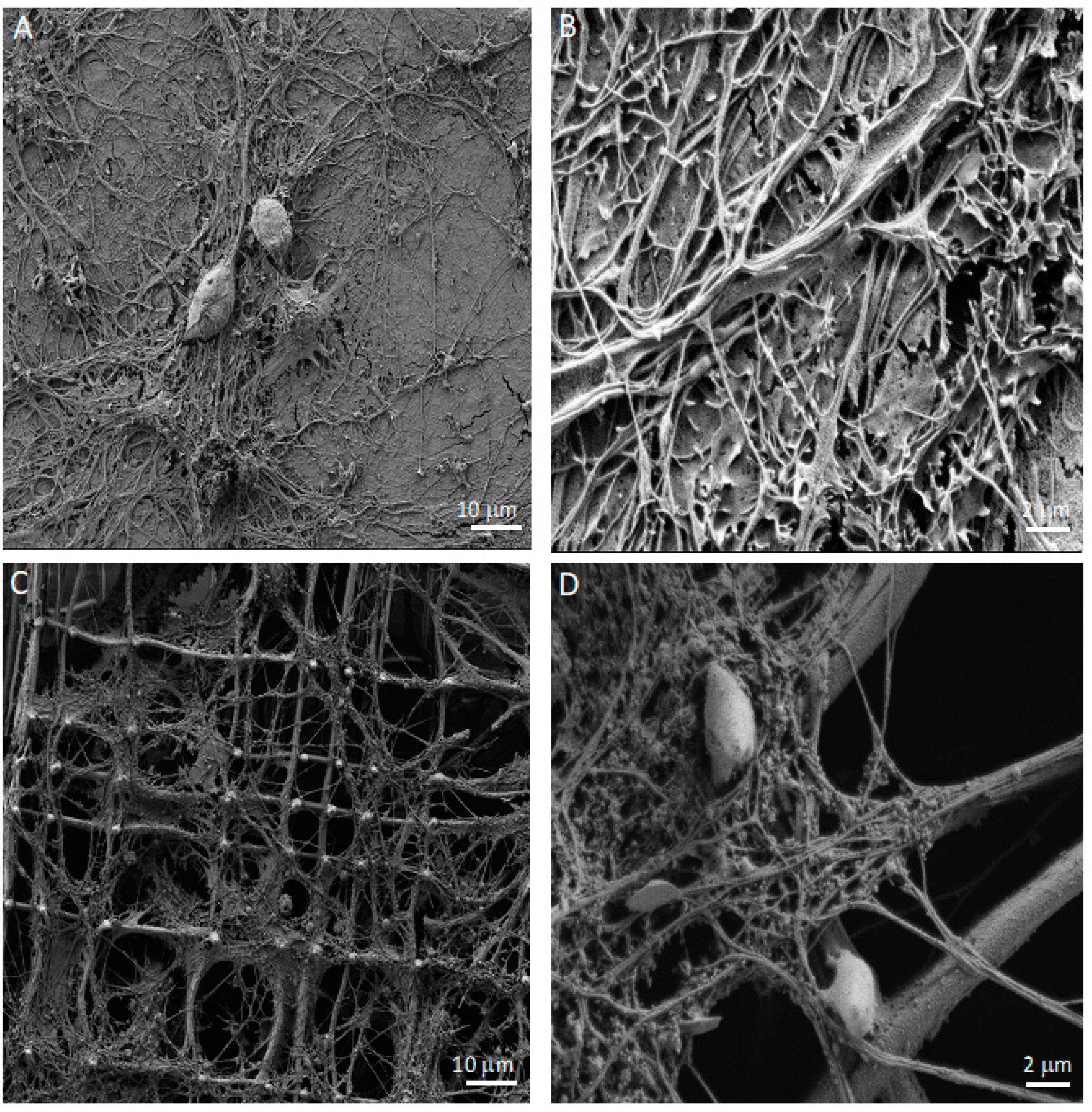
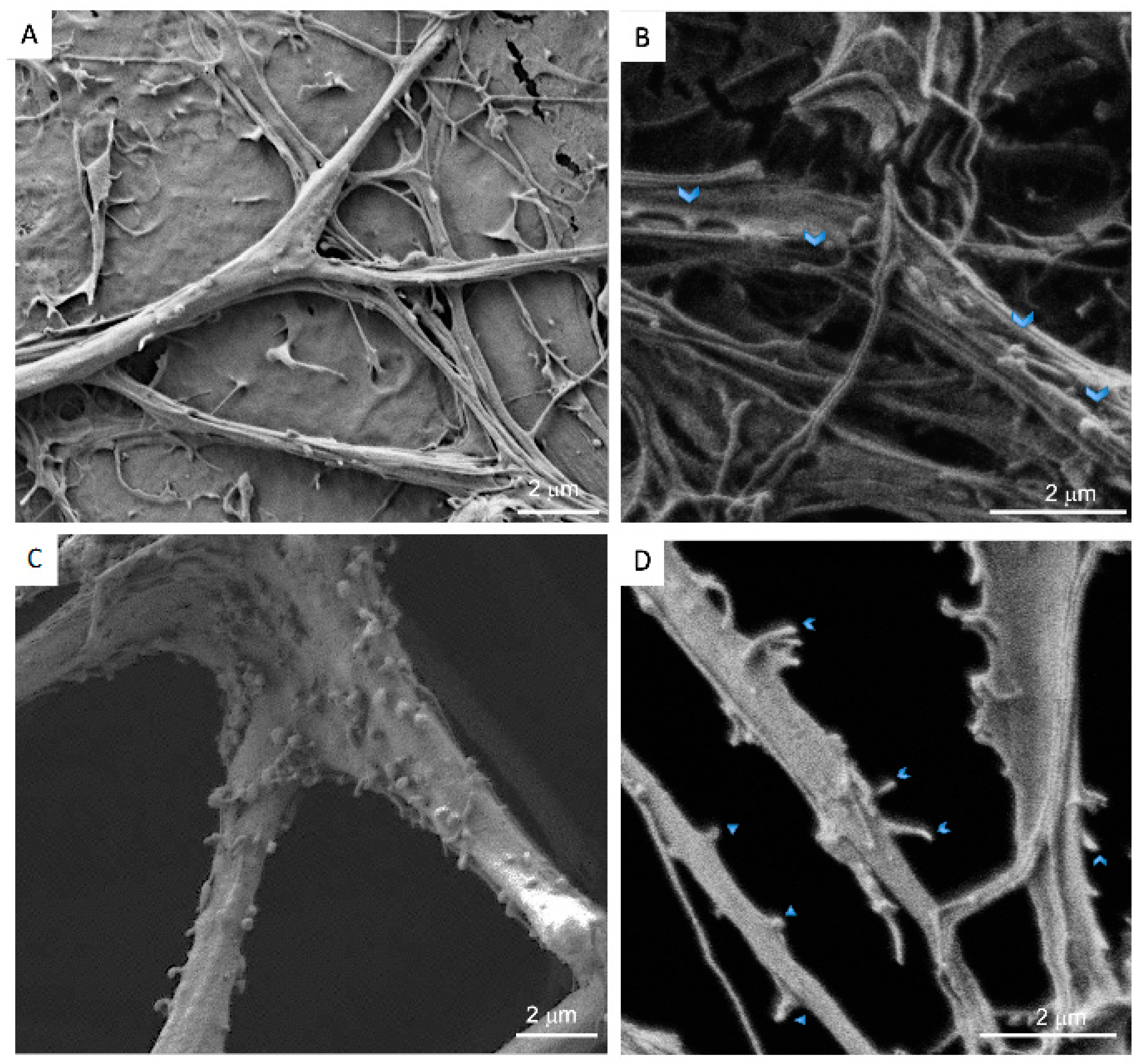
2.8. Scanning Electron Microscopy (SEM)
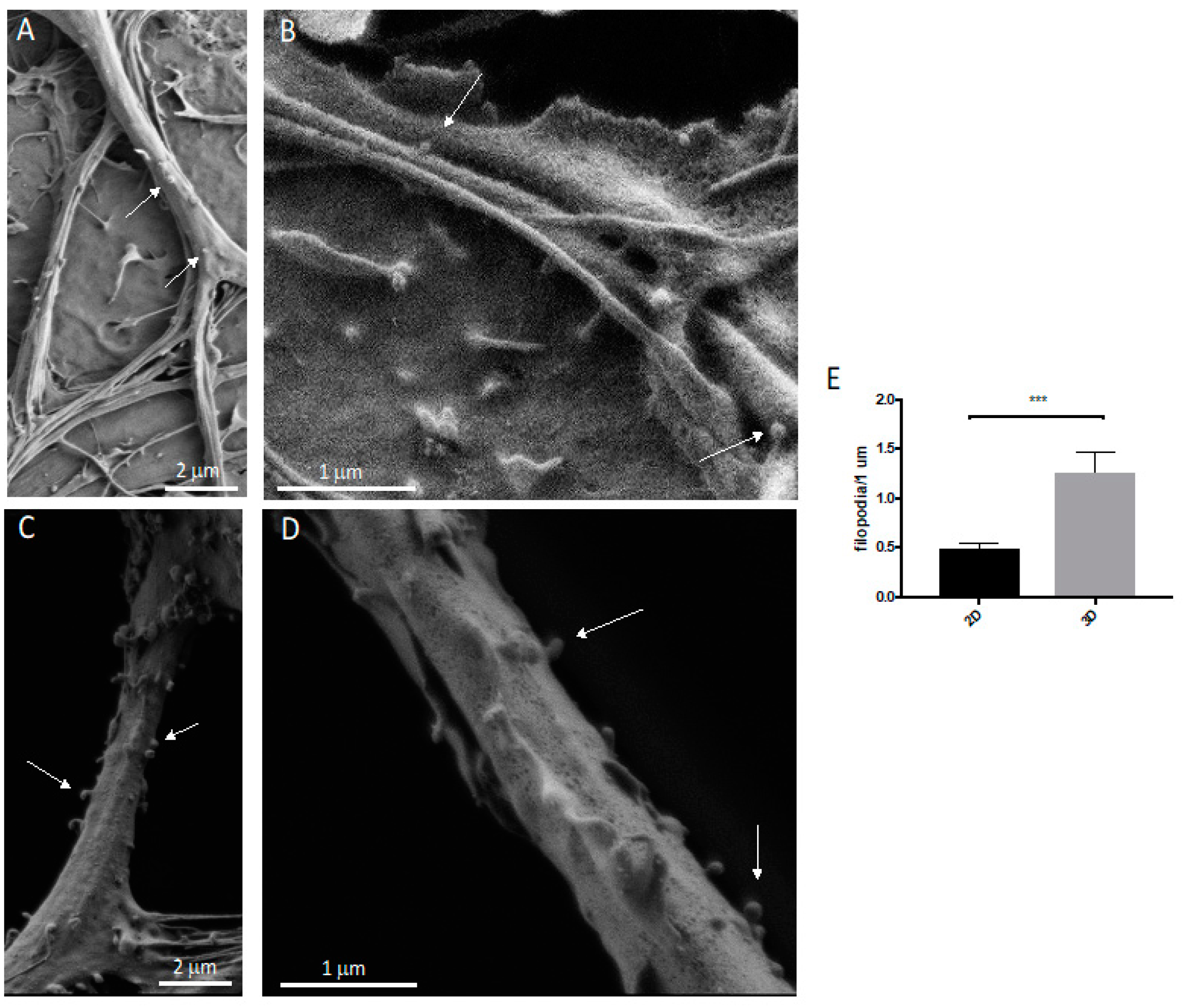
2.9. Neuronal Protrusion Analysis
2.10. Statistical Analysis
3. Results
3.1. Co-Cultures of Astrocytes and Neurons in 2D vs. 3D Microscaffold
3.2. Neurons: Spines/Protrusions
3.3. Spine/Protrusion Counts in 2D vs. 3D
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chiaradia, I.; Lancaster, M.A. Brain organoids for the study of human neurobiology at the interface of in vitro and in vivo. Nat. Neurosci. 2020, 23, 1496–1508. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, M.A. Brain organoids: A new frontier of human neuroscience research. Semin. Cell Dev. Biol. 2021, 111, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Pacitti, D.; Privolizzi, R.; Bax, B.E. Organs to Cells and Cells to Organoids: The Evolution of in vitro Central Nervous System Modelling. Front. Cell. Neurosci. 2019, 13, 129. [Google Scholar] [CrossRef] [PubMed]
- Nikolakopoulou, P.; Rauti, R.; Voulgaris, D.; Shlomy, I.; Maoz, B.M.; Herland, A. Recent progress in translational engineered in vitro models of the central nervous system. Brain 2020, 143, 3181–3213. [Google Scholar] [CrossRef]
- Zhuang, P.; Sun, A.X.; An, J.; Chua, C.K.; Chew, S.Y. 3D neural tissue models: From spheroids to bioprinting. Biomaterials 2018, 154, 113–133. [Google Scholar] [CrossRef]
- Gribkoff, V.K.; Kaczmarek, L.K. The need for new approaches in CNS drug discovery: Why drugs have failed, and what can be done to improve outcomes. Neuropharmacology 2017, 120, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Kesselheim, A.S.; Hwang, T.J.; Franklin, J.M. Two decades of new drug development for central nervous system disorders. Nat. Rev. Drug Discov. 2015, 14, 815–816. [Google Scholar] [CrossRef]
- Wang, H. Modeling Neurological Diseases With Human Brain Organoids. Front. Synaptic Neurosci. 2018, 10, 15. [Google Scholar] [CrossRef]
- Pfeiffer, B.E.; Huber, K.M. The State of Synapses in Fragile X Syndrome. Neuroscientist 2009, 15, 549–567. [Google Scholar] [CrossRef]
- Stephan, K.E.; Baldeweg, T.; Friston, K.J. Synaptic plasticity and dysconnection in schizophrenia. Biol. Psychiatry 2006, 59, 929–939. [Google Scholar] [CrossRef]
- Calabresi, P.; Picconi, B.; Parnetti, L.; Di Filippo, M. A convergent model for cognitive dysfunctions in Parkinson’s disease: The critical dopamine-acetylcholine synaptic balance. Lancet Neurol. 2006, 5, 974–983. [Google Scholar] [CrossRef]
- Südhof, T.C. Neuroligins and Neurexins Link Synaptic Function to Cognitive Disease. Nature 2008, 455, 903–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selkoe, D.J. Alzheimer’s disease is a synaptic failure. Science 2002, 298, 789–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welch, J.M.; Lu, J.; Rodriguiz, R.M.; Trotta, N.C.; Peca, J.; Ding, J.-D.; Feliciano, C.; Chen, M.; Adams, J.P.; Luo, J.; et al. Cortico-striatal synaptic defects and OCD-like behaviors in SAPAP3 mutant mice. Nature 2007, 448, 894–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kauer, J.A.; Malenka, R.C. Synaptic plasticity and addiction. Nat. Rev. Neurosci. 2007, 8, 844–858. [Google Scholar] [CrossRef]
- Araque, A.; Perea, G. Glial modulation of synaptic transmission in culture. Glia 2004, 47, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Bernardinelli, Y.; Muller, D.; Nikonenko, I. Astrocyte-synapse structural plasticity. Neural Plast. 2014, 2014, 232105. [Google Scholar] [CrossRef] [Green Version]
- Zehendner, C.M.; White, R.; Hedrich, J.; Luhmann, H.J. A neurovascular blood-brain barrier in vitro model. Methods Mol. Biol. 2014, 1135, 403–413. [Google Scholar] [CrossRef]
- Shein-Idelson, M.; Ben-Jacob, E.; Hanein, Y. Engineered Neuronal Circuits: A New Platform for Studying the Role of Modular Topology. Front. Neuroeng. 2011, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Simi, A.; Amin, H.; Maccione, A.; Nieus, T.; Berdondini, L. Integration of microstructured scaffolds, neurons, and multielectrode arrays. Prog. Brain Res. 2014, 214, 415–442. [Google Scholar] [CrossRef]
- Imamura, Y.; Mukohara, T.; Shimono, Y.; Funakoshi, Y.; Chayahara, N.; Toyoda, M.; Kiyota, N.; Takao, S.; Kono, S.; Nakatsura, T.; et al. Comparison of 2D- and 3D-culture models as drug-testing platforms in breast cancer. Oncol. Rep. 2015, 33, 1837–1843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Pekkanen-Mattila, M.; Shahsavani, M.; Falk, A.; Teixeira, A.I.; Herland, A. A 3D Alzheimer’s disease culture model and the induction of P21-activated kinase mediated sensing in iPSC derived neurons. Biomaterials 2014, 35, 1420–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulloa Severino, F.P.; Ban, J.; Song, Q.; Tang, M.; Bianconi, G.; Cheng, G.; Torre, V. The role of dimensionality in neuronal network dynamics. Sci. Rep. 2016, 6, 29640. [Google Scholar] [CrossRef] [Green Version]
- Baker, B.M.; Chen, C.S. Deconstructing the third dimension: How 3D culture microenvironments alter cellular cues. J. Cell Sci. 2012, 125, 3015–3024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irons, H.R.; Cullen, D.K.; Shapiro, N.P.; Lambert, N.A.; Lee, R.H.; Laplaca, M.C. Three-dimensional neural constructs: A novel platform for neurophysiological investigation. J. Neural Eng. 2008, 5, 333–341. [Google Scholar] [CrossRef]
- Puschmann, T.B.; Zandén, C.; De Pablo, Y.; Kirchhoff, F.; Pekna, M.; Liu, J.; Pekny, M. Bioactive 3D cell culture system minimizes cellular stress and maintains the in vivo-like morphological complexity of astroglial cells. Glia 2013, 61, 432–440. [Google Scholar] [CrossRef]
- Forro, C.; Caron, D.; Angotzi, G.N.; Gallo, V.; Berdondini, L.; Santoro, F.; Palazzolo, G.; Panuccio, G. Electrophysiology Read-Out Tools for Brain-on-Chip Biotechnology. Micromachines 2021, 12, 124. [Google Scholar] [CrossRef]
- Nava, M.M.; Piuma, A.; Figliuzzi, M.; Cattaneo, I.; Bonandrini, B.; Zandrini, T.; Cerullo, G.; Osellame, R.; Remuzzi, A.; Raimondi, M.T. Two-photon polymerized “nichoid” substrates maintain function of pluripotent stem cells when expanded under feeder-free conditions. Stem Cell Res. Ther. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Ovsianikov, A.; Gruene, M.; Pflaum, M.; Koch, L.; Maiorana, F.; Wilhelmi, M.; Haverich, A.; Chichkov, B. Laser printing of cells into 3D scaffolds. Biofabrication 2010, 2, 014104. [Google Scholar] [CrossRef]
- Raimondi, M.T.; Nava, M.M.; Eaton, S.M.; Bernasconi, A.; Vishnubhatla, K.C.; Cerullo, G.; Osellame, R. Optimization of Femtosecond Laser Polymerized Structural Niches to Control Mesenchymal Stromal Cell Fate in Culture. Micromachines 2014, 5, 341–358. [Google Scholar] [CrossRef] [Green Version]
- Zandrini, T.; Shan, O.; Parodi, V.; Cerullo, G.; Raimondi, M.T.; Osellame, R. Multi-foci laser microfabrication of 3D polymeric scaffolds for stem cell expansion in regenerative medicine. Sci. Rep. 2019, 9, 11761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sclip, A.; Arnaboldi, A.; Colombo, I.; Veglianese, P.; Colombo, L.; Messa, M.; Mancini, S.; Cimini, S.; Morelli, F.; Antoniou, X.; et al. Soluble Aβ oligomer-induced synaptopathy: C-Jun N-terminal kinase’s role. J. Mol. Cell Biol. 2013, 5, 277–279. [Google Scholar] [CrossRef] [PubMed]
- Colnaghi, L.; Russo, L.; Natale, C.; Restelli, E.; Cagnotto, A.; Salmona, M.; Chiesa, R.; Fioriti, L. Super Resolution Microscopy of SUMO Proteins in Neurons. Front. Cell. Neurosci. 2019, 13, 486. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drisaldi, B.; Colnaghi, L.; Fioriti, L.; Rao, N.; Myers, C.; Snyder, A.M.; Metzger, D.J.; Tarasoff, J.; Konstantinov, E.; Fraser, P.E.; et al. SUMOylation Is an Inhibitory Constraint that Regulates the Prion-like Aggregation and Activity of CPEB3. Cell Rep. 2015, 11, 1694–1702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raimondi, M.T.; Eaton, S.M.; Laganà, M.; Aprile, V.; Nava, M.M.; Cerullo, G.; Osellame, R. Three-dimensional structural niches engineered via two-photon laser polymerization promote stem cell homing. Acta Biomater. 2013, 9, 4579–4584. [Google Scholar] [CrossRef]
- Remuzzi, A.; Bonandrini, B.; Tironi, M.; Longaretti, L.; Figliuzzi, M.; Conti, S.; Zandrini, T.; Osellame, R.; Cerullo, G.; Raimondi, M.T. Effect of the 3D Artificial Nichoid on the Morphology and Mechanobiological Response of Mesenchymal Stem Cells Cultured In Vitro. Cells 2020, 9, 1873. [Google Scholar] [CrossRef]
- Führmann, T.; Hillen, L.M.; Montzka, K.; Wöltje, M.; Brook, G.A. Cell-cell interactions of human neural progenitor-derived astrocytes within a microstructured 3D-scaffold. Biomaterials 2010, 31, 7705–7715. [Google Scholar] [CrossRef]
- Piccolo, S.; Dupont, S.; Cordenonsi, M. The biology of YAP/TAZ: Hippo signaling and beyond. Physiol. Rev. 2014, 94, 1287–1312. [Google Scholar] [CrossRef]
- Reilly, G.; Engler, A. Intrinsic Extracellular Matrix Properties Regulate Stem Cell Differentiation. J. Biomech. 2009, 43, 55–62. [Google Scholar] [CrossRef]
- Aoki, C.; Sherpa, A.D. Making of a Synapse: Recurrent Roles of Drebrin A at Excitatory Synapses Throughout Life. Adv. Exp. Med. Biol. 2017, 1006, 119–139. [Google Scholar] [CrossRef] [PubMed]
- Jacchetti, E.; Nasehi, R.; Boeri, L.; Parodi, V.; Negro, A.; Albani, D.; Osellame, R.; Cerullo, G.; Matas, J.F.R.; Raimondi, M.T. The nuclear import of the transcription factor MyoD is reduced in mesenchymal stem cells grown in a 3D micro-engineered niche. Sci. Rep. 2021, 11, 3021. [Google Scholar] [CrossRef] [PubMed]
- Carelli, S.; Giallongo, T.; Rey, F.; Barzaghini, B.; Zandrini, T.; Pulcinelli, A.; Nardomarino, R.; Cerullo, G.; Osellame, R.; Cereda, C.; et al. Neural precursors cells expanded in a 3D micro-engineered niche present enhanced therapeutic efficacy in vivo. Nanotheranostics 2021, 5, 8–26. [Google Scholar] [CrossRef] [PubMed]
- Haber, M.; Zhou, L.; Murai, K.K. Cooperative astrocyte and dendritic spine dynamics at hippocampal excitatory synapses. J. Neurosci. 2006, 26, 8881–8891. [Google Scholar] [CrossRef]
- Murphy, A.R.; Laslett, A.; O’Brien, C.M.; Cameron, N.R. Scaffolds for 3D in vitro culture of neural lineage cells. Acta Biomater. 2017, 54, 1–20. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Musi, C.A.; Colnaghi, L.; Giani, A.; Priori, E.C.; Marchini, G.; Tironi, M.; Conci, C.; Cerullo, G.; Osellame, R.; Raimondi, M.T.; et al. Effect of 3D Synthetic Microscaffold Nichoid on the Morphology of Cultured Hippocampal Neurons and Astrocytes. Cells 2022, 11, 2008. https://doi.org/10.3390/cells11132008
Musi CA, Colnaghi L, Giani A, Priori EC, Marchini G, Tironi M, Conci C, Cerullo G, Osellame R, Raimondi MT, et al. Effect of 3D Synthetic Microscaffold Nichoid on the Morphology of Cultured Hippocampal Neurons and Astrocytes. Cells. 2022; 11(13):2008. https://doi.org/10.3390/cells11132008
Chicago/Turabian StyleMusi, Clara Alice, Luca Colnaghi, Arianna Giani, Erica Cecilia Priori, Giacomo Marchini, Matteo Tironi, Claudio Conci, Giulio Cerullo, Roberto Osellame, Manuela Teresa Raimondi, and et al. 2022. "Effect of 3D Synthetic Microscaffold Nichoid on the Morphology of Cultured Hippocampal Neurons and Astrocytes" Cells 11, no. 13: 2008. https://doi.org/10.3390/cells11132008
APA StyleMusi, C. A., Colnaghi, L., Giani, A., Priori, E. C., Marchini, G., Tironi, M., Conci, C., Cerullo, G., Osellame, R., Raimondi, M. T., Remuzzi, A., & Borsello, T. (2022). Effect of 3D Synthetic Microscaffold Nichoid on the Morphology of Cultured Hippocampal Neurons and Astrocytes. Cells, 11(13), 2008. https://doi.org/10.3390/cells11132008